Молекулярные участники рецепторного пути регуляции апоптоза в опухолевых и нормальных лимфоцитах в условиях ингибирования белка теплового шока 90 in vitro

Автор: Рязанцева Н.В., Кайгородова Е.В., Новицкий В.В., Белкина М.В., Марошкина А.Н., Клименченко М.В., Таширева Л.А.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 2 (44), 2011 года.
Бесплатный доступ
Проведено исследование программированной гибели лимфоцитов, полученных у здоровых доноров, и опухолевых кле- ток линии Jurkat в условиях культивирования с селективным ингибитором Hsp90 (Heat shock protein - Hsp) 17-AAG. Оценку реализации апоптоза проводили методом флуоресцентной микроскопии с использованием FITC-меченного аннексина V и пропидий иодида. Активность каспазы-8 определяли спектрофотометрическим методом. Установлено, что ингибирование Hsp90 приводит к активации каспазы-8, увеличению числа клеток, презентрирующих TNFR1, что облегчает запуск програм- мированной гибели опухолевых клеток линии Jurkat.
Короткий адрес: https://sciup.org/14055897
IDR: 14055897 | УДК: (612.112.94+616.155.32)-091.818-092.4
Molecular participants of receptor pathway of apoptosis regulation in tumor and normal lympfocytes in inhibition of heat shock protein 90 in vitro
The study of programmed death of lymphocytes from healthy donors and tumor-cell line Jurkat in culture conditions with a selective inhibitor of Hsp90 (Heat shock protein - Hsp) 17-AAG was investigated. Assessment of apoptosis was performed by fluorescent microscopy using FITC-labeled annexin V and propidium iodide. Caspase-8 was determined by spectrophotometer assay. It has been established that inhibition of Hsp9 leads to activation of caspase-8, increasing the number of TNFR1-positive cells, which facilitates the launch of the programmed death of tumor cell line Jurkat.
Текст научной статьи Молекулярные участники рецепторного пути регуляции апоптоза в опухолевых и нормальных лимфоцитах в условиях ингибирования белка теплового шока 90 in vitro
Идентификация молекулярных механизмов эндогенной и экзогенной регуляции апоптоза, уравновешивающего эффекты клеточной пролиферации, элиминации поврежденных, функционально неполноценных и опухолевых клеток, является актуальным направлением фундаментальных исследований. Механизмы, участвующие в запуске и реализации апоптоза, весьма разнообразны [1, 4]. Известно, что после индукции апоптоза дальнейшая судьба клетки – гибель или выживание – зависит от наличия или активации многочисленных модуляторов программированной клеточной гибели. К числу последних относятся белки теплового шока (Heat shock proteins – Hsps). Данные протеины принимают участие в формировании трехмерной конформации вновь синтезированных полипептидов; поддержании функциональной активности внутриклеточных белков и элиминации поврежденных белковых молекул, а также в осуществлении транспорта через клеточные мембраны, процессах ассоциации-диссоциации внутриклеточных надмолекулярных комплексов, защите белков от агрегации [8]. Роль Hsps в апоптозе неоднозначна: в одних случаях они обеспечивают выживание клетки, а в других – способствуют ее гибели [4, 5].
Одним из представителей семейства белков теплового шока является Hsp90. Известно, что белок теплового шока 90 обладает преимущественно антиапоптотическим эффектом и имеет большое значение в регуляции рецепторного и митохондриального путей апоптоза [9, 11]. Рецептор-опосредованный путь апоптоза может запускаться при связывании специфических лигандов c рецепторами семейства фактора некроза опухоли (TNF), расположенными на плазматической мембране [13]. Презентация рецепторов на плазматической мембране осуществляется при помощи молекулярных шаперонов, к группе которых относится белок теплового шока Hsp90 [10]. Также показано, что Hsp90 и Hsp70 регулируют функции комплекса IKK (IκB (inhibitors of NF-κB) kinase), являющегося основным активатором NF-kappaB. Белки теплового шока приводят к диссоциации комплекса IKK, что подавляет NF-kappaB-опосредованную трансдукцию сигнала [12]. Повышенная экспрессия Hsp90 может рассматриваться среди причин устойчивости трансформированных клеток к апоптогенным сигналам [6, 7]. Наряду с этим остаётся нерешённым вопрос о молекулярных механизмах участия вышеуказанного шаперона в регуляции апоптоза опухолевых и нормальных клеток.
В связи с этим целью настоящей работы явилась оценка экспрессии молекулярных участников рецепторного пути апоптоза (TNFR1,TNFα, sTNFR1, каспаза-8) в опухолевых и нормальных лимфоцитах при ингбировании белка теплового шока 90 in vitro.
Материалы и методы
В качестве материала исследования были использованы опухолевая клеточная линия Jurkat (Т-лимфобластного лейкоза человека), полученная из Российской коллекции клеточных культур Института цитологии РАН (г. Санкт-Петербург), и мононуклеарные лейкоциты, выделенные из крови относительно здоровых доноров (11 мужчин и 16 женщин в возрасте от 18 до 45 лет).
Культивирование опухолевых клеток линии Jurkat проводили суспензионным методом в полной питательной среде, содержащей 90 % RPMI-1640 («Вектор-Бест», Россия), 10 % эмбриональной телячьей сыворотки («Invitrogen», США), инактивированной при 56°С в течение 30 мин, 0,3 мг/мл L-глутамина («Вектор-Бест», Россия) и 100 мкг/мл гентамицина («INS», США) при температуре 37°С и в 5 % атмосфере СО2. Клетки поддерживали в логарифмической фазе роста и пересаживали через 3 сут. Ингибирование белка теплового шока Hsp90 в клетках осуществляли с помощью добавления в культуральную среду 17-аллиламино-17-деметоксигелданамицин (17-AAG) («Sigma Aldrich», США) в концентрации 5,0 мкМ.
Выделение мононуклеарных лейкоцитов проводили методом градиентного центрифугирования в стерильных условиях из венозной гепаринизированной крови на градиенте плотности (ρ=1,077 г/см3) Ficoll-Paque (‹‹Pharmacia››, Швеция) в соотношении 1:2. Оценку апоптоза проводили методом флуоресцентной микроскопии на микроскопе «Axiostar plus» («Carl Zeiss», Германия) с использованием FITC-меченного аннексина V и пропидий иодида («Abcam», Великобритания) согласно инструкции фирмы-производителя.
Уровень фосфорилированной и нефосфо-рилированной формы Hsp90 в опухолевых и нормальных лимфоцитах определяли методом вестерн-иммуноблоттинга, используя первичные моноклональные антитела к белкам Hsp90-фосфо-S226 и Hsp90 («Abcam», Великобритания) и вторичные антитела, конъюгированные с HRP. Детекцию результатов осуществляли с помощью хемилюминесцентного субстрата Novex («Invitrogen», США) на рентгеновской пленке («Kodak X-ray film», США). Вывод о содержании исследуемого белка в клетке делали по изменению отношения величины сигнала метки искомого белка к величине сигнала с фермента глицеральдегид-3-фосфатдегидрогеназы, используя программное обеспечение TotalLab. Результаты выражали в условных единицах.
Количество TNFR1-презентирующих клеток
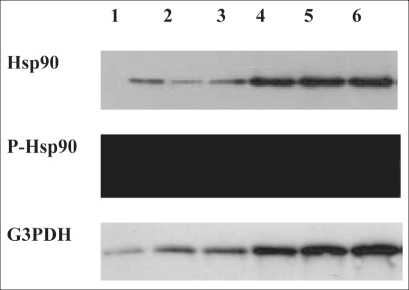
Рис. 1. Вестерн-блоттинг белка теплового шока Hsp90 в опухолевых и нормальных лимфоцитах. Примечание: 1–3 – лизаты мононуклеарных лейкоцитов здоровых доноров; 4–6 – лизаты опухолевых клеток линии Jurkat;
G3PDF-глицеральдегид – 3 – фосфатдегидрогеназа определяли методом проточной цитофлуори-метрии с помощью набора моноклональных антител к человеческому антигену CD120 («Invitrogen», США) на цитофлуориметре FACS-Canto II («Becton Dickinson», США). Результаты выражали в % от общего числа клеток.
Уровнь sTNF-R и TNFα в супернатантах опухолевых клеток Jurkat оценивали методом иммуноферментного анализа по протоколу фирмы-производителя («Bender MedSystem», Австрия). Измерения проводили при 450 нм на микропланшетном фотометре Multiscan EX («Thermo Labsystem», Финляндия). Количество sTNF-R и TNFα в супернатантах опухолевых клеток линии Jurkat определяли по калибровочной кривой и выражали в нг/мл.
Активность каспазы-8 определяли с помощью спектрофотометрического метода согласно инструкции фирмы-производителя («Abcam», Великобритания). Анализ основан на спектрофотометрической детекции хромофора р-nitroanilide (p-NA), который отщепляется под действием каспазы-8 из субстрата IETD-pNA. Эмиссию света pNA измеряли при длине волны 400 и 405 нм. Результаты выражали в коэффициентах активности каспазы-8, соответственно, показывающих соотношение прироста оптической плотности pNA опытных и контрольных образцов с учетом содержания белка в пробе. Концентрацию белка в пробе определяли методом Бретфорда.
Полученные данные обрабатывали методами вариационной статистики. Проверку нормальности распределения количественных показателей проводили с использованием критерия Шапиро-Вилка. Достоверность различий оценивали с помощью непараметрического критерия Манна–Уитни (для выборок, подчиняющихся ненормальному закону распределения) и двухвыборочного критерия Стьюдента (для выборок, подчиняющихся нормальному закону распределения). Данные представлены в виде медианы (Ме), верхнего и нижнего квартилей (Q1-Q3). Статистически значимыми различия считались при р<0,05.
Результаты и обсуждение
На первом этапе работы с помощью вестерн-блоттинга было оценено внутриклеточное содержание фосфорилированной и нефосфори-лированной формы Hsp90 в опухолевых и нормальных лимфоцитах. Результаты исследования показали повышенное количество фосфорилированной (активной) формы Hsp90 в опухолевых клетках линии Jurkat (7,01 (6,18-7,91) усл.ед.) по сравнению с мононуклеарными лейкоцитами, полученными из крови относительно здоровых доноров, где данная форма белка теплового шока 90 практически не обнаруживалась. Уровень нефосфорилированной формы Hsp90 в клетках Jurkat и мононуклеарных лейкоцитах не различался (5,83(5,62-6,16) усл. ед. и 4,67(4,426,86) усл.ед. соответственно) (рис. 1).
Известно, что опухолевые клетки в процессе канцерогенеза вырабатывают собственные защитные белки, вследствие чего они значительно отличаются от нормальных клеток организма, в том числе и по способности вступать в апоптоз. Одним из важнейших антагонистов проапоптоз-ных молекул в процессе программированной клеточной смерти является белок теплового шока Hsp90 [1, 4, 5].
В этой связи на втором этапе исследования нами была проведена оценка количества апоп-тотически измененных клеток линии Jurkat и мононуклеарных лейкоцитов в условиях их культивирования со специфическим ингибитором Hsp90 17-AAG в концентрации 5мкМ. При исследовании апоптоза в интактных опухолевых клетках линии Jurkat было обнаружено, что число апоптотически измененных клеток составляет 4,99 (1,78–6,38) %. Добавление в
Таблица
Изменения молекулярных показателей рецепторного пути апоптоза опухолевых клеток линии Jurkat при действии ингибитора Hsp90 in vitro Mе (Q1-Q3)
|
Показатели |
Интактная культура Jurkat |
Культивирование опухолевых клеток линии Jurkat с 5мкМ 17-AAG |
|
TNF-R1, % |
3,80 (1,80–7,20) |
21,90 (20,30–27,250) р<0,05 |
|
sTNF-R, нг/мл |
1,715 (1,655–1,880) |
1,685 (1,670–1,745) р>0,05 |
|
TNFα, нг/мл |
0,058 (0,024–0,069) |
0,040 (0,031–0,053) р>0,05 |
|
Активность каспазы-8, усл. ед. |
0,53 (0,46–0,58) |
1,00 (0,98–1,30) р<0,05 |
Примечание: р– статистическая значимость различий по сравнению с аналогичным показателем в интактной культуре.
среду культивирования 5мкМ ингибитора 17-AAG приводило к увеличению числа клеток в апоптозе до 25,96 (18,62–30,78) % (р1<0,05) по сравнению с интактной культурой. Повышение экспрессии белка теплового шока Hsp90 подавляет программированную клеточную смерть, так как данный шаперон регулирует активность многих транскрипционных факторов, способствующих выживанию клеток [9]. Действие ингибитора Hsp90 сопровождается проведением «смертельных» сигналов внутрь клетки и повышением активности эффекторных каспаз, что неизбежно приводит к увеличению уровня клеток, вступивших в апоптоз [1]. Именно эта способность Hsp90 положена в основу разработки противоопухолевых лекарств на основе ингибиторов Hsps [3]. Анализ содержания апоптотически измененных клеток в культуре мононуклеарных лейкоцитов у здоровых доноров не выявил достоверных различий между интактной культурой и при добавлении ингибитора Hsp90 in vitro – 19,13 (13,84–22,10) % и 22,44 (20,93–23,59) % соответственно.
На третьем этапе исследования для оценки молекулярных механизмов рецепторного пути регуляции апоптоза клеток в условиях действия ингибитора Hsp90 in vitro были оценены TNFR1,TNFα, sTNFR1 и каспаза-8 (таблица). Было показано, что добавление в культуру опухолевых клеток линии Jurkat селективного ингибитора 17-AAG в концентрации 5мкМ увеличивало в 7 раз количество клеток, презентирующих на своей поверхности TNF-R1, по сравнению с интактной культурой. Однако содержание TNFα и sTNF-R в культуральной среде не изменялось.
Одним из главных молекулярных участников рецептор-опосредованного пути апоптоза является каспаза-8 [2]. Было установлено, что при действии ингибитора Hsp90 17-AAG активность каспазы-8 в опухолевых клетках линии Jurkat повышается (р<0,05). Исходя из полученных данных, можно заключить, что белок Hsp90 влияет на активацию инициаторной каспазы-8. Нами высказано предположение, что Hsp90 регулирует рецепторный путь реализации апоптоза не только через RIP, Akt, NF-κB [4], но и напрямую, ингибируя превращение прокаспазы-8 в активную форму.
Таким образом, в опухолевых клетках линии Jurkat, в отличие от мононуклеарных лейкоцитов, полученных у здоровых доноров, повышено содержание фосфорилированной формы белка Hsр90, играющей в опухолевых клетках антиапоптотическую роль. Действие специфического ингибитора Hsр90 17-AAG на культуру клеток линии Jurkat приводило к возрастанию активности каспазы-8, увеличению количества TNF-R1- презентирующих клеток. В связи с этим с практической точки зрения возможность влиять на противоапоптотическую роль Hsp90 в опухолевых клетках с помощью его ингибиторов, например 17-AAG, представляет большой интерес для разработки новых подходов в тар-гетной терапии онкологических заболеваний.
Работа выполнена в рамках Федеральной целевой программы «Научные и научнопедагогические кадры инновационной России»
на 2009–2013 г. (ГК П1203; ГК 02.740.11.0311), поддержана грантами Carl Zeiss, РФФИ (№ 0904-99025) и Совета по грантам при Президенте РФ (ГК№ 02.120.11.3842-МД, МК-480.2011.7).
