Морфогенез побега возобновления в предзимний период у луковичных и клубнелуковичных эфемероидов

Автор: Седельникова Л.Л.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 4, 2019 года.
Бесплатный доступ
Цель работы - сравнительный анализ морфогенеза луковичных и клубнелуковичных эфемероидов для выявления состояния побега возобновления в предзимье в условиях лесостепной зоны Новосибирской области. Проанализированы особенности органогенеза по бега возобновления у раннецветущих луковичных и клубнелуковичных геофитов. Представлено описание апекса генеративного побега у семи видов: Eryhtronium sibiricum, Chionodoxa gigantea, Muscari armeniacum, M. aucheri, Puschkinia scilloides, Scilla sibirica, Crocus hybridus. Установлено, что для этих видов свойственно короткое ранневесеннее надземное развитие побега в течение 1,5 месяцев, чему предшествует продолжительное подземное развитие в течение 22-23 месяцев. Сделано заключение, что в генеративный период онтогенеза у изученных растений в предзимье проявляется приуроченность дифференциации конуса нарастания побега возобновления к определенным этапам органогенеза. У видов, цветущих ранней весной, - Muscari aucheri, Crocus hybridus, Chionodoxa gigantean, Scilla sibirica, Eryhtronium sibiricum перед вынужденным зимним покоем луковиц и клубнелуко-виц в побеге возобновления сформированы органы цветка и соцветия, что определяет их состояние VI-VIII этапами органогенеза...
Короткий адрес: https://sciup.org/140243425
IDR: 140243425 | УДК: 581.4:582.572.7/.8+581.543.4
Morphogenesis of shoot renewal in pre-winter period in bulbous and corms ephemeroids
The aim of the study was comparative analysis of the morphogenesis of bulbous and tuberous ephemeroids to identify the state of shoot renewal in pre-winter in the forest-steppe zone of Novosibirsk Region. The features of organogenesis of shoot renewal from early-flowering bulbous and bulbotuberiferous geoview were analyzed. The description of generative apex of root from seven species: Erythronium sibiricum, Chionodoxa gigantea, Muscari armeniacum, M. aucheri, Puschkinia scilloides, Scilla sibirica, Crocus hybridus was presented. It was established that these species were characterized by a short early spring aboveground development of shoot renewal within 1.5 months preceded by a long underground development for 22-23 months. The conclusion was made that in generative period of ontogenesis in studied plants in pre-winter period there was the correlation of differentiation of the cone of growth of the shoot of renewal to certain stages of organo- genesis. The species flowering in early spring Muscari aucheri, hybridus, Crocus, Chionodoxa gigantea, Scilla sibirica, Eryhtronium sibiricum before winter forced the rest of the bulbs and corms to have the shoot of a resume-formed organs of the flower and inflorescence, which determined their state of VI-VIII stages of organogenesis...
Текст научной статьи Морфогенез побега возобновления в предзимний период у луковичных и клубнелуковичных эфемероидов
Введение. Луковичные и клубнелуковичные эфемероиды представляют одну из групп растений, скелетная основа жизненной формы которых сведена до минимума. Их морфогенез достаточно хорошо выражен в адаптации формообразования органов к условиям обитания. Это позволило довольно широко исследовать данные геофиты в 60–80-х годах прошлого столетия [1–5] в местах обитания, не свойственных их естественным эколого-географическим условиям существования. Многофункциональность структурных органов побега формировалась в процессе эволюции и выполняла приспособительную роль у растений с эфемероидным типом развития. В связи с этим наличие двух типов вынужденного покоя, летнего и зимнего, связано с адаптацией органов к самосохранению и выживанию. Жизнедеятельность апекса побега при этом претерпевает короткое надземное и длительное подземное развитие. Последнее обеспечивает побегу поэтапное развитие вегетативных и генеративных органов, исследование которых актуально для осуществления биологического контроля за процессами морфогенеза у луковичных и клубнелуковичных растений в течение сезонного развития. Использова- ние новейшего оборудования дает точную информацию внутрипочечного формирования вегетативных и генеративных зачаточных органов побега, что послужило основанием для выполнения работы.
Цель работы . Сравнительный анализ морфогенеза у луковичных и клубнелуковичных эфемероидов для выявления состояния побега возобновления в предзимье в условиях лесостепной зоны Новосибирской области.
Объекты и методы исследований. Работа выполнена в Центральном сибирском ботаническом саду (ЦСБС СО РАН) г. Новосибирска. Объектами служили луковичные растения семейства Liliaceae Juss. – Eryhtronium sibiricum (Fischer et Meyer) Krylov – кандык сибирский; Hyacinthaceae Batsch Chionodoxa gigantea Witt – хионодокса гигантская, Muscari armeniacum Leichtl. ex Baker 'Blue Spike' – мускари (мышиный гиацинт) армянский, сорт Блу Спайк , отборная бело-розовая форма, M. aucheri Boiss. – мускари ауши, Puschkinia scilloides Adams – пушкиния пролесковидная, Scilla sibirica Haw. – сцилла (пролеска) сибирская; клубнелуковичные из Iridaceae Juss. Crocus hybridus hort. 'Flower Record' – крокус гибридный, сорт Флауэр Рекорд . Растения выращивали на участке лаборатории декоративных растений, расположенном в юго-восточном районе лесостепной Приобской агроклиматической провинции. Изучали растения генеративного возрастного состояния. Пробы для морфоанализа взяты 15.10.2018 г. Описание состояния конуса нарастания побега и определение этапов его органогенеза проводили по методике [4, 6]. Апикальная зона побега возобновления проанализирована с помощью стереомикроскопа Carl Zeiss Sterio Discovery V 12 с использованием микрофотографий, полученных в центре коллективного пользования ЦСБС СО РАН. Морфологическое описание побеговой системы сделано согласно работам [7, 8].
Результаты исследований и их обсуждение. В условиях лесостепной зоны Западной Сибири эфемероидные геофиты цветут сразу после схода снежного покрова – с последней декады апреля по третью декаду мая. Вегетационный период короткий и продолжается 30– 46 дней, во второй – третьей декаде июня надземная часть отмирает, а подземная вступает в вынужденный летний покой. Продолжитель- ность жизни побега возобновления от его формирования в почке до вегетации и цветения составляет около двух лет [9]. По феноритмотипу это коротковегетирующие весеннецветущие луковичные и клубнелуковичные многолетники, зимующие в грунте. Апекс побега возобновления ранневесенних луковичных и клубнелуковичных растений переходит из вегетативного в генеративное состояние в период летнего покоя и к осени уходит в зимний покой.
Взрослые луковицы и клубнелуковицы имели индивидуальные морфометрические показания у видов в осенний период 2018 г. (табл.). У них хорошо сформирована корневая система мочковатого типа. Длина корней у видов составляла от 1,0 до 6,5 см. К осеннему укоренению приурочен VI этап органогенеза. На период первой декады октября у Chionodoxa gigantea, Crocus hybridus, Eryhtronium sibiricum Muscari armeniacum сорт'Blue Spike' сформирован один генеративный побег, у Scilla sibirica 1–2, у M. aucheri 2–3 побега. У луковиц 1–2 покровные чешуи и 1–15 запасающих чешуй. Одну запасающую чешую имели ежегодно заменяющиеся луковицы туникатного типа кандыка сибирского и клубнелуковицы крокуса гибридного.
Морфометрические показатели (min-max) состояния луковичных и клубнелуковичных геофитов в предзимний период (10.10.2018 г.)
|
Вид |
Луковица, см |
Число, шт. |
Длина корней, см |
||
|
длина |
ширина |
листьев |
чешуй* |
||
|
Chionodoxa gigantea |
2,0±0,2 |
1,0±0,1 |
2 |
15 |
1,5–3,0 |
|
Crocus hybridus ** |
0,5±0,01 |
1,2±0,1 |
3-4 |
1 |
3,0–6,3 |
|
Eryhtronium sibiricum |
4,5±0,2 |
1,0±0,1 |
2 |
1 |
1,0–1,5 |
|
Muscari armeniacum |
1,5±0,1 |
0,6±0,1 |
3 |
9-10 |
1,5–2,0 |
|
M. aucheri |
3,5±0,2 |
2,0±0,2 |
4 |
15 |
2,5–6,5 |
|
Puschkinia scilloides |
2,0±0,3 |
1,7±0,2 |
2 |
8-10 |
1,0–1,5 |
|
Scilla sibirica |
2,5±0,3 |
2,0±0,3 |
2 |
10-11 |
1,5–2,0 |
* – зачаточные чешуи; ** – клубнелуковичный геофит.
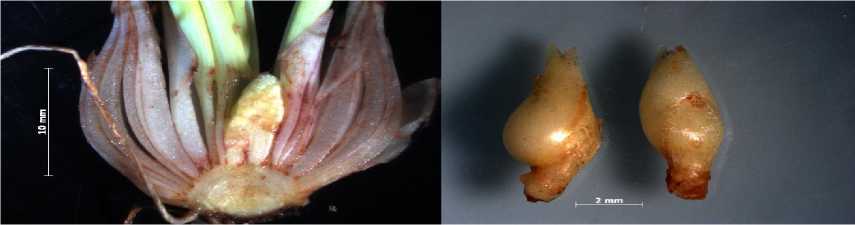
Зимнезеленые листья наблюдали только у M. aucheri и бело-розовой формы от M. armeniacum. Вегетативный апекс побега экзогенно формирует на поверхности так называемые валики или зачатки листьев – листовые примордии, появляющиеся в акропетальном направлении. Число зачаточных листьев от 2 до 4 (см. табл.). Органы цветка также изначально появляются в виде экзогенных выростов на апексе акропетально. Это хорошо выражено у представителей рода Muscari Mill., у которых в предзимье сформировано зачаточное соцветие из 20–25 цветков, нижние из которых имели зачатки пыльников и завязи, что характерно для VII–VIII этапов органогенеза (рис. 1 а, в). Под первой чешуей заложены зачаточные луковички, так называемые детки (рис.1, г). У M. aucheri отчетливо выражено на многолетнем побеге-луковице два сохранившихся остатка соцветий прошедшего года вегетации (2018 г.) и два зачаточных соцветия (2019 г.) (рис. 1, а). У сорта мускари Блу Спайк (рис. 1, б) позднего срока цветения сформировано одно соцветие, только с элементами лепестков цветка (VI этап органогенеза).
Морфоанализ луковиц Eryhtronium sibiricum показал, что развитие побега возобновления на генеративные органы цветка ускорено и соответствует VIII этапу органогенеза. У побега сформировано два зачаточных листа, лепестки околоцветника и андроцея, в котором наблюдали формирование пыльцевых зерен – микроспорогенез (рис. 2 а, б ). Причем явление протандрии четко прослеживается уже на ранних этапах органогенеза внутрипочечного состояния побега, где наблюдали отставание развития гинецея. Формируется одиночный цветок. В базальной части луковицы заложена новая почка возобновления 2020 г. вегетации, которая в течение двух лет развивается подземно и ее конус нарастания на период от 10.10. 18 г. первого года соответствует II этапу органогенеза (рис. 2, в, г ). Следует отметить, что такая почка заложена у всех исследованных видов.

в г
Рис. 1. Состояние побега возобновления у Muscari aucheri (а), M. armeniacum сорт Blue Spike (б), отборная форма бело-розовая (в), зачатки луковичек-деток (г)
Состояние побега возобновления у Scilla sibirica соответствует VII–VIII этапам органогенеза. У побега два супротивных листа, верхняя часть которых ассимилирована и выступает из луковицы (рис. 3, а). В них как бы вложено соцветие, которое, несмотря на зачаточное состояние, отчетливо просматривается и имеет четыре цветка (рис. 3, в, г). У цветка отчетливо видны примордии чашелистиков, частично окрашенные лепестки околоцветника (рис. 3, д). Сформированы зачатки гинецея и андроцея. В базальной части побег плотно окружен двумя молодыми запасающими чешуями (рис. 3, б). Луковица нарастающего типа, состоит из запасающих чешуй, которые с возрастом наружно отмирают.
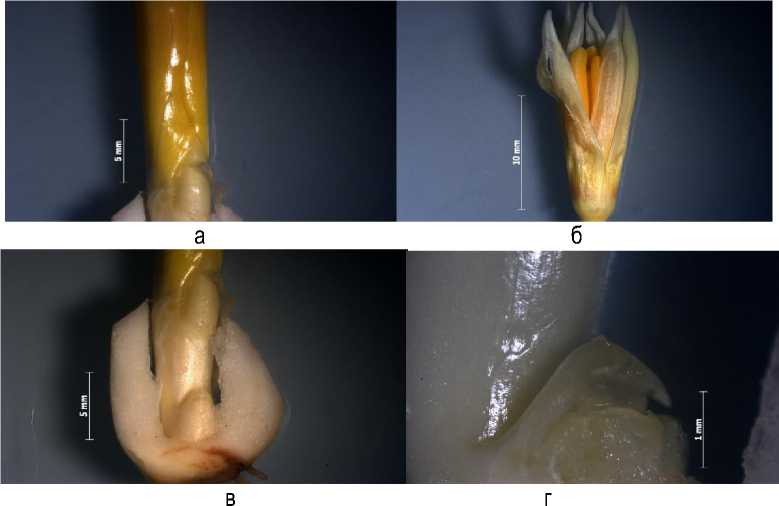
Рис. 2. Состояние побега возобновления Eryhtronium sibiricum: а – два зачаточных листа; б – зачаточный цветок; в, г – почка возобновления

а б

в г д
Рис. 3. Побег возобновления Scilla sibirica в предзимний период
У побега возобновления Chionodoxa gigantea наблюдали наличие двух зачаточных листьев и цветка. Отмечено окрашивание долей около- цветника и отношение состояния побега к VIII этапу органогенеза (рис. 4). В зачаточном бутоне сформированы пыльники и завязь.
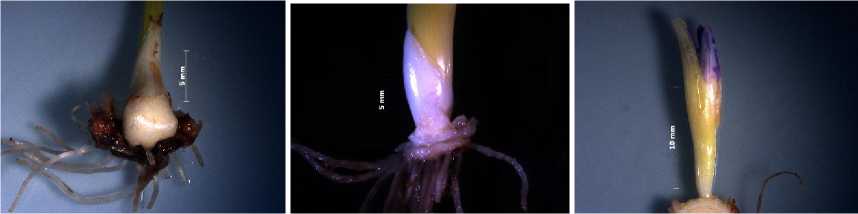
Рис. 4. Побег возобновления Chionodoxa gigantea в предзимье
Для луковиц Puschkinia scilloides характерно отставание в развитии побега возобновления, и его конус нарастания находится в период дифференциации двух листовых примордиев и на- чала дифференциации соцветия на IV–V этапах органогенеза (рис. 5). Это соответствует более позднему цветению этого вида весной.
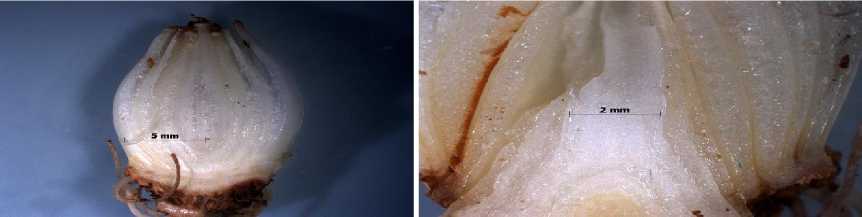
Рис. 5. Продольный срез луковицы Puschkinia scilloides в предзимье
Сформированные органы цветка установлены у клубнелуковичного геофита Crocus hybridus hort. сорт Flower Record. Причем сначала происходит формирование листовых пласти- нок в числе 4–5 шт., затем органов цветка. Развитие тычинок (протерандрия) опережает рост рыльца и лепестков околоцветника (рис. 6). Формируется одиночный цветок.

Рис. 6. Crocus hybridus hort. сорт Flower Record в предзимье
Таким образом, апекс растительного побега представленных луковичных и клубнелуковичных геофитов в предзимний период переходит от вегетативного к генеративному состоянию. Наступает фаза флоральной инициалии. Это довольно критический период в онтогенезе побега и особи, который предвещает переход растения от скрытого к открытому генеративному состоянию. Однако дальнейшие процессы роста и развития резко затормаживает наступление холодов. Наступает зимний период покоя. Среда, таким образом, играет разрешающую, а не детерминирующую роль в развитии эфемеро-идных геофитов. Понятия летнего и зимнего покоя у луковичных растений весьма условны, так как в побеге возобновления непрерывно происходят органообразовательные и замещающие процессы [9]. Адаптация формообразования побега возобновления у исследованных видов к условиям среды происходит на структурном уровне. При введении в лесостепную зону Западной Сибири луковичных и клубнелуковичных эфемероидов целесообразно осуществлять биологический контроль за их развитием.
Выводы
-
1. В генеративный период онтогенеза проявляется приуроченность дифференциации конуса нарастания побега возобновления IV–VIII
-
2. В предзимний период у видов, цветущих ранней весной (III декада апреля – II декада мая), – M. aucheri, Crocus hybridus, Chionodoxa gigantea, Scilla sibirica, Eryhtronium sibiricum – у побега возобновления сформированы органы цветка и соцветия, что определяет их состояние VII–VIII этапами органогенеза.
-
3. У позднецветущего вида (I–III декады мая) Puschkinia scilloides и M. armeniacum сорт Blue Spike конус нарастания побега возобновления в предзимье соответствует IV–VI этапам органогенеза.
этапам органогенеза в предзимний период подземного развития луковиц и клубнелуковиц.
Список литературы Морфогенез побега возобновления в предзимний период у луковичных и клубнелуковичных эфемероидов
- Капинос Г.Е. Биологические закономерности развития луковичных и клубнелуковичных растений на Апшероне. -Баку: АН АзССР, 1965. -240 с.
- Седова Е.А. Закономерности органогенеза луковичных и клубнелуковичных геофитов. -М.: МГУ, 1976, -30 с.
- Скрипчинский В.В., Скрипчинский Вл.В. Морфобиологические основы онтогенеза эфемероидных геофитов и проблема его эволюционного становления//Проблемы экологической морфологии растений: тр. МОИП. -М.: Наука, 1976. -Т.42. -С.167-185.
- Куперман Ф.М. Морфофизиология растений. -М.: Высш. шк., 1977. -288 с.
- Ростовцева З.П. Верхушечная меристема. -М.: МГУ, 1969. -80 с.
- Дударь Ю.А. Сезонные ритмы биопроцессоров у растений: адаптация или абаптация?//Общая биология. -1993. -Т.54, № 3. -С. 229-332.
- Жмылев П.Ю., Алексеев Ю.Е., Карпухина Е.А. Основные термины и понятия современной биоморфологии. -М.: МГУ, 1993. -147 с.
- Серебрякова Т.И., Воронин Н.С., Еленевский А.Г. . Ботаника с основами фитоценологии. Анатомия и морфология растений. -М.: Академкнига, 2006. -543 с.
- Седельникова Л.Л. Биоморфология геофитов в Западной Сибири. -Новосибирск: Наука, 2002. -307 с.
