Морфологическая характеристика постоянных моляров из палеолитических слоев Денисовой пещеры

Автор: Зубова А.В., Чикишева Т.А., Шуньков М.В.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология и палеогенетика
Статья в выпуске: 1 т.45, 2017 года.
Бесплатный доступ
В статье анализируется морфология двух постоянных зубов, обнаруженных в плейстоценовых слоях Денисовой пещеры в Горном Алтае. Образец Денисова 4 представляет собой верхний левый третий (второй?) моляр, Денисова 8 -верхний левый третий моляр. Обе находки были обследованы по расширенной одонтологической программе. Результаты проведенного исследования продемонстрировали высокий уровень значимости одонтологических данных для анализа межгрупповой дифференциации в пределах рода Homo. Они подтверждают, что денисовцы или H. altaiensis составляют особую группу гомининов, которая отличается от H. sapiens и H. neanderthalensis не только генетически, но и морфологически. Одонтологическая специфика денисовцев заключается в мегадонтизме и наличии признаков, сближающих эти находки с эректусами Сангирана и среднеплейстоценовыми гомининами Китая, которым принадлежала стоянка Суйцзиайяо. Анализ морфологии верхних моляров денисовца также позволил сделать предположение о таксономической принадлежности неидентифицированной части денисовского генома представителям вида H. erectus. Выявленные особенности зубной системы H. altaiensis свидетельствуют о том, что он представлял собой крайне консервативную форму.
Денисовцы, средний/верхний палеолит, одонтология
Короткий адрес: https://sciup.org/145145790
IDR: 145145790 | УДК: 572.77 | DOI: 10.17746/1563-0102.2017.45.1.121-134
Текст научной статьи Морфологическая характеристика постоянных моляров из палеолитических слоев Денисовой пещеры
Археологический памятник Денисова пещера находится на юге Западно-Сибирского региона, в горах Алтая, недалеко от границы Российской Федерации с Китаем, Монголией и Казахстаном.
История изучения останков плейстоценовых го-мининов из Денисовой пещеры насчитывает более 30 лет. Первые одонтологические материалы были получены в 1984 г.: в стратиграфическом слое 21.1 обнаружен молочный моляр Денисова 2 [Шпакова, Деревянко, 2000]. В том же году в слое 12 найден зуб, который сначала был атрибутирован как верхний постоянный резец [Turner, 1990; Шпакова, Деревянко, 2000], а позже по результатам сравнительных исследований определен как принадлежащий животному из семейства бовидов [Viola et al., 2011, p. 209]. В 2000 г. из слоя 11.1 был извлечен верхний постоянный моляр очень хорошей сохранности (Денисова 4), в 2010 г. в подошве стратиграфического слоя 11.4 на контакте со слоем 12 были обнаружены фрагменты коронки верхнего постоянного моляра Денисова 8.
По стоянные моляры Денисова 4 и Денисова 8 по геохронологическим данным датируются периодом 50–40 тыс. л.н., однако слой, содержавший Денисова 4, моложе, чем слой, из которого извлечен моляр Денисова 8 [Sawyer et al., 2015].
Результаты секвенирования митохондриального и ядерного геномов из дистальной фаланги мизинца кисти девочки 6–7 лет, обнаруженной в слое 11.2 (Денисова 3), и моляра Денисова 4 показали, что находки принадлежат ранее неизвестному науке гоминину, обозначенному авторами исследования как денисо-вец [Reich et al., 2010] или Homo altaiensis [Деревянко, 2011]. Впервые в антропологической науке новая группа гомининов была выделена не по морфологическим критериям, а по данным генетического анализа.
Анализ митохондриального генома показал, что расхождение ДНК предков денисовского человека, неандертальцев и Homo sapiens имело место ок. 1 млн л.н. [Krause et al., 2010]. Ядерная ДНК дает более позднюю дату: отделение предков H. sapiens от общего ствола, объединявшего их с эволюционными предшественниками H. neanderthalensis и H. altaiensis , произошло ок. 800 тыс. л.н. [Meyer et al., 2012]. Разделение предков денисовцев и неандертальцев по первоначальным данным состоялось ок. 640 тыс. л.н. [Reich et al., 2010], а по результатам последних исследований – 430 тыс. л.н. [Meyer et al., 2016].
Период существования денисовцев в Горном Алтае был продолжительным: если исходить из скорости мутирования мтДНК современного человека, то можно предположить, что индивиды Денисова 2 и Денисова 8 на 65 тыс. лет древнее, чем Денисова 3 и Денисова 4 [Slon et al., 2015]. Как было установлено, в составе денисовского генома присутствовали аллели, ассоциируемые у современных людей с темным цветом кожи, каштановыми волосами и карими глазами [Meyer et al., 2012]. Согласно данным митохондриального генома всех индивидов, останки которых обнаружены в Денисовой пещере, уровень генетического разнообразия в этой популяции был заметно ниже, чем среди неандертальцев и людей современного вида [Meyer et al., 2012; Slon et al., 2015].
Вопросы о локализации очага формирования денисовского таксона и его статусе в рамках существующей палеоантропологической классификации являются предметом дискуссий. Опираясь на археологические данные, А.П. Деревянко выдвинул гипотезу о том, что денисовец представляет один из древних подвидов полиморфного вида H. sapiens , существовавших наряду с подвидами H. sapiens neanderthalensis, H. sapiens africaniensis, H. sapiens orientalensis [Деревянко, 2011], его формирование происходило на основе азиатских представителей вида H. erectus [Там же]. Однако последние палеогенетические данные, полученные для плейстоценовых гомининов из местонахождения Сима де Лос Уэсос в Испании, свидетельствуют о близости состава митохондриального генома этих популяций и денисовцев [Meyer et al., 2016]. Это ставит под вопрос азиатское происхождение наиболее древних компонентов в генофонде H. altaiensis .
В настоящей статье представлены результаты расширенного морфологического анализа постоянных моляров Денисова 4 и Денисова 8, который позволил определить основные направления биологических связей денисовской популяции и в первом приближении сравнить особенности зубной системы H. altaien-sis c характеристиками таксонов H. erectus s.l., H. hei-delbergensis s.l., H. neanderthalensis, H. sapiens s.l.
Программа исследования
Описание одонтологических находок из Денисовой пещеры на основе стандартного протокола ASUDAS и программы учета неандертальских апоморфий ранее было выполнено в контексте палеогенетического изучения денисовских гомининов [Krause et al., 2010; Sawyer et al., 2015].
Нами проведено расширенное обследование верхних моляров Денисова 4 и Денисова 8 по значительно бóльшему набору признаков. В качестве базовых схем одонтологического обследования взяты ASU-DAS [Turner, Nichol, Scott, 1991; Scott, Turner, 1997] и система А.А. Зубова, принятая в российской одонтологии и включающая в программу описания полного узора борозд коронки [1968, 1974, 2006; Зубов, Халде-ева, 1989, 1993]. Привлекались также программа учета маркеров неандертальского комплекса [Bailey, 2002;
Bailey, Skinner, Hublin, 2011] и программа, разработанная для описания одонтологической специфики архаичных антропологических форм [Зубова, 2013].
Каждая из указанных систем имеет свои преимущества, позволяющие максимально использовать информационные возможности одонтологического материала. Система ASUDAS включает наиболее диверсифицированные шкалы учета одонтологических признаков, которые предельно точно соотносятся с различными морфологическими вариантами в популяциях человека современного вида. Главным преимуществом признаков, описанных Ш. Бэйли, является их высокая чувствительность к наличию в составе популяции неандертальских генов. Программа А.А. Зубова дает возможность провести анализ не только деталей макрорельефа, но и вариантов узора коронок зубов, представляющих собой независимую, иерархически организованную систему признаков – одонтоглифику.
Всего учитывались более 60 признаков, на обследованных молярах обнаружены следующие: редукция гипоконуса, редукция метаконуса, ромбовидная форма верхних моляров, метаконулюс, мезио стиль, затек эмали, бугорок Карабелли, дистальный и мезиальный дополнительные бугорки верхних моляров, эпикриста, плагиокриста (косой гребень), энтокриста, задняя ямка, одонтоглифические элементы.
Редукция гипоконуса. Признак характеризует динамику размеров гипоконуса относительно протоконуса и метаконуса. Шкалы систем ASUDAS и А.А. Зубова имеют различия: первая включает 6 баллов (0 – отсутствие бугорка, 5 – максимальное развитие) [Turner, Nichol, Scott, 1991, р. 18], вторая – 4 балла (4, 4–, 3+, 3), где балл 4 описывает максимальное развитие бугорка, а балл 3 – его отсутствие [Зубов, 1968, с. 152].
Редукция метаконуса. Признак характеризует динамику размеров метаконуса относительно параконуса. В системе ASUDAS оценивается аналогично редукции гипоконуса [Turner, Nichol, Scott, 1991, р. 18], в системе А.А. Зубова принята 5-балльная шкала (от 1 до 5), где отсутствию редукции соответствует балл 1, максимальному ее уровню – балл 5 [Зубов, 1968, с. 160].
Ромбовидная форма верхних моляров. Специфическая форма верхних постоянных моляров, с нередуцированным, сильно выступающим в дисто-лингваль-ном направлении гипоконусом и редуцированным метаконусом, формирующим скошенный дисто-ве-стибулярный угол коронки [Bailey, 2004].
Метаконулюс. Небольшой бугорок в эмалевой толще осевого гребня метаконуса, в центральной его части [Зубов, Халдеева, 1993, с. 68].
Бугорок Карабелли. Стилоидный бугорок в мезио-лингвальном отделе коронки. В системе ASUDAS оценивается баллами от 0 до 7, где 0 – отсутствие признака, 7 – максимальное развитие бугорка [Turner, Nichol, Scott, 1991, p. 19]. В системе А.А. Зубова аналогич- ным образом оценивается баллами от 0 до 5 [1968, с. 157]. Соотношение баллов следующее:
ASUDAS Система А.А. Зубова
-
1 ,21
-
3 ,42
Дистальный дополнительный бугорок верхних моляров. Стилоидный бугорок на дистальном краевом гребне, в зоне контакта метаконуса и гипоконуса. В обеих системах используется шкала, где отсутствию бугорка соответствует балл 1, максимальному его развитию – балл 6 [Зубов, 2006, с. 56; Turner, Nichol, Scott, 1991, p. 19].
Мезиальный дополнительный бугорок верхних моляров. Бугорок на мезиальном крае коронки. Выделяется дополнительной бороздой, которая впадает в фиссуру, разделяющую параконус и протоконус. Филогенетически отличается от бугорков, формируемых дистальными сегментами параконуса и протоконуса [Зубов, Халдеева, 1993, с. 67, Scott, Turner, 1997, p. 45]. Шкала для определения балла выраженности этого признака не разработана.
Эпикриста. Гребень, соединяющий протоконус и параконус, фиксируется при полном перекрывании фиссуры, разделяющей бугорки.
Плагиокриста. Гребень, соединяющий метаконус и протоконус. В данном исследовании для его фиксации используется 4-балльная шкала [Зубова, 2013, с. 114].
Энтокриста. Краевой гребень, соединяющий протоконус и гипоконус [Зубов, Халдеева, 1989, с. 62]. Практически никогда не встречается у современного человека.
Задняя ямка. Представляет собой вытянутое углубление в дистальном отделе верхних и нижних моляров, которое может иметь различную протяженность. На верхних молярах современного человека она, как правило, локализована на окклюзивной поверхности метаконуса, где с мезиальной стороны ограничивается его дистальным сегментом, а с дистальной – дистальным краевым гребнем [Зубов, 2006, с. 61]. На древних формах может заходить также на поверхность гипоконуса, где ограничивается аналогичными элементами этого бугорка. Специальные шкалы учета степени выраженности данного признака не разработаны.
Одонтоглифические элементы. Как и обычные одонтологические признаки, они различаются по уровню таксономической значимости; их комбинации отражают эволюционный статус популяции [Зубов, 1974]. Номенклатура одонтоглифических элементов в про- цессе разработки неоднократно менялась, что обусловило несогласованность в системе обозначений признаков, используемых разными исследователями (см.: [Зубов, 1974, 2006, Зубов, Халдеева, 1989; Hillson, 1996]). Мы опираемся на последние работы А.А. Зубова [Зубов, Халдеева, 1989, Зубов, 2006]. Ниже приводится классификация одонтоглифических признаков для верхних моляров и раскрывается морфологическое содержание каждого обозначения.
Борозды окклюзальной поверхности коронок подразделяются на три типа. Наиболее древними по происхождению являются интертуберкулярные борозды первого порядка. Они дифференцируют главные бугорки коронки и обозначаются римскими цифрами I–IV (I–VI – на нижних). Фиссура I отделяет метаконус от параконуса, фиссура II – параконус от протоконуса, фиссура III – метаконус от протоконуса, фиссура IV – гипоконус от метаконуса и протоконуса.
Туберкулярные борозды второго порядка ограничивают осевые гребни каждого бугорка, расчленяя его на три сегмента. В работах некоторых исследователей эти сегменты обозначаются как мезиальный, центральный и дистальный [Carlsen, 1987; Bailey, Skinner, Hublin, 2011]. Межсегментарные борозды маркируются арабскими цифрами (1 и 2) и первыми двумя буквами названия бугорка (на нижних – тремя) (например, ра – параконус, prd – протоконид) [Зубов, 1974].
Они отделяют на метаконусе и параконусе осевой сегмент от мезиального (борозда 1) и дистального (борозда 2) и обозначаются 1me, 2 me, 1pa, 2pa соответственно. На протоконусе и гипоконусе борозда 1 отделяет осевой сегмент от дистального (1pr, 1hy), борозда 2 – от мезиального (2pr, 2hy).
Остальные борозды относятся к третьему порядку и подразделяются на две категории. Первая включает дополнительные бороздки, которые разделяют мезиальные и дистальные сегменты каждого бугорка на две части в продольном направлении, пролегающие параллельно бороздам 1 и 2. Они обозначаются 1’ и 2’ соответственно. Во вторую категорию входят дополнительные бороздки осевых гребней главных бугорков. Они обозначаются цифрами 3 и 4 [Зубов, Халде-ева, 1989; Зубов, 2006].
На метаконусе это 1’me (впадает в фиссуру I, разделяет мезиальный сегмент в вестибулярном направлении) и 2’me (впадает в фиссуру III параллельно задней ямке, разделяет дистальный сегмент в вестибулярном направлении). На параконусе – 1’pa (впадает в фиссуру I, разделяет дистальный сегмент в вестибулярном направлении), 2’pa (впадает в фиссуру II, разделяет мезиальный сегмент в вестибулярном направлении), 3pa (впадает в фиссуру II, разделяет осевой сегмент в вестибулярном направлении), 4pa (рассекает осевой сегмент в мезио-дистальном направлении). На протоконусе – 1’pr (впадает в фиссуру IV, разде- ляет дистальный сегмент в лингвальном направлении), 2’pr (впадает в фиссуру II, разделяет мезиальный сегмент в лингвальном направлении), 2’’pr (дублирует 2’pr в мезиальном отделе, впадает в фиссуру II или в бороздку 2’pr), 3pr (впадает в фиссуру II и центральную ямку, делит осевой сегмент в лингвальном направлении), 4pr (рассекает осевой сегмент в поперечном направлении). На гипоконусе – 1’hy (впадает в фиссуру IV или заднюю ямку, разделяет дистальный сегмент в лингвальном направлении), 2’hy (впадает в фиссуру IV или бассейн талона, разделяет мезиальный сегмент в лингвальном направлении).
Общее количество туберкулярных борозд и направление хода некоторых из них различаются у представителей разных видов в пределах рода Homo . Наибольшая вариабельность характерна для положения борозд третьего порядка, многие из которых у современных представителей H. sapiens редуцированы и встречаются крайне редко. У ранних представителей рода Homo наблюдается повышенная складчатость эмали зубов, могут быть дополнительные борозды третьего порядка, отсутствующие в поздних группах. Может также меняться положение точек впадения туберкулярных борозд в межбугорковые фиссуры. Кроме того, у представителей эволюционно ранних форм более выражена, чем у H. sapiens, тенденция к гребнеобразованию.
Все учитываемые признаки подразделяются на три группы. К первой относятся эволюционно стабильные, таксономически нейтральные характеристики, связанные с базовой морфологией зубной системы в пределах гомининной линии эволюции и маркирующие эволюционную преемственность (например, четырехбугорковые верхние моляры и пятибугорковые нижние, количество интертуберкулярных борозд коронки). Они стабильно присутствуют на всех стадиях эволюции рода Homo.
Во вторую группу входят т.н. маркеры генерализованной архаики – плезиоморфные признаки, имеющие отрицательную эволюционную динамику, т.е. их частоты наиболее высоки на ранних ступенях эволюции рода
Homo
и уменьшаются по мере приближения к современности. К числу таких признаков относятся дериваты цингулюма, гребнеобразующие структуры моляров, соотношения M1
Третью группу составляют эволюционно прогрессивные в пределах рода Homo признаки, подразделяющиеся на два блока. Один из них объединяет показатели редукции зубной системы, стабильно увеличивающейся на гомининной линии эволюции (редукция гипоконуса, метаконуса, уменьшение размеров дистальных зубов в ряду, малые размеры или отсутствие стило- идных бугорков дистального отдела нижних моляров, осевое положение гипоконулида, упрощенный одон-тоглифический узор и т.п.). Другой блок включает апо-морфные характеристики отдельных таксонов.
При описании верхних моляров из Денисовой пещеры основное внимание уделялось признакам второй и третьей категории – плезиоморфным и маркерам неандертальского и сапиентного комплексов. Зафиксированный набор признаков был сопоставлен с характеристиками таксонов H. erectus s.l., H. heidelber-gensis s.l., H. neanderthalensis, H. sapiens s.l.
Для сравнительного анализа использовались оригинальные данные авторов, а также опубликованные высокоточные фотографии и морфологические описания находок. Список сравнительных материалов представлен в таблице.
Материалы
Обсуждаемые зубы из Денисовой пещеры – два постоянных моляра. Моляр Денисова 4 из литологического слоя 11.1 определен Б. Виолой как верхний левый третий зуб [Viola et al., 2011], хотя распределение фасеток стертости дает некоторые основания считать его вторым верхним моляром при отсутствующем третьем. Исследователь описал макрорельеф и одонто-метрические характеристики зуба, а также отметил его одонтометрическое сходство с зубами ранних представителей рода Homo и с австралопитековыми формами при отсутствии какого-либо морфологического сходства с неандертальскими образцами.
Моляр Денисова 8 относится также к левой стороне верхней челюсти. А.П. Бужилова определила его как верхний второй или третий постоянный коренной зуб [Buzhilova, 2014]. Б. Виола считает более вероятным, что это левый верхний третий моляр [Sawyer et al., 2015]. Данный зуб сохранился хуже, чем Денисова 4: его корневая система полностью утрачена, коронка восстановлена из фрагментов, а ее мезиальный отдел в районе контакта параконуса и протоконуса разрушен. Рельеф бугорков тригона практически полностью стерт, как и бóльшая часть межбугорковых фиссур. При этом эмаль гипоконуса и дистального отдела метаконуса практически не стерта, небольшая контактная фасетка наблюдается только на самой вершине главного гребня гипоконуса, в ее мезиальном отделе. На интерпроксимальной поверхности в дистальном отделе контактная фасетка отсутствует.
Морфологическая характеристика
Левый верхний постоянный второй или третий моляр (М2/3) из слоя 11.1 – Денисова 4 (рис. 1). Принад- лежал молодому взрослому индивиду мужского пола [Slon et al., 2015].
Зуб очень массивный, с длинной шейкой и сильно расходящимися в мезиальной и дистальной норме корнями. Коронка с округленными углами, благодаря относительно небольшому, по сравнению с параконусом, букко-лингвальному диаметру метаконуса и умеренной скошенности мезио-буккального угла, она сужена в дистальном отделе, но не имеет ромбовидной формы, характерной для неандертальских верхних моляров, поскольку отсутствует лингвальное смещение гипоконуса (рис. 2). Вершины главных бугорков наклонены к центру коронки. Межбугорковые фиссуры очень глубокие. Фиссуры I и III пролегают только по окклюзивной поверхности, а II и IV выходят и на вертикальные плоскости зуба: фиссура IV, разделяющая гипоконус и протоконус, распространяется на лингвальную поверхность, а борозда II, дифференцирующая протоконус и параконус, – на мезиальную (рис. 3).
Параконус разделен на три сегмента глубокими бороздами, рассекающими край коронки и выходящими на буккальную сторону в верхней ее части. Его мезиальный гребень шире, чем дистальный, который на половине длины осевого гребня исчезает, а выделяющая его бороздка сливается с межбугорковой фиссурой, разделяющей метаконус и параконус. На параконусе имеются глубокие борозды 1ра и 2ра, первая из которых впадает в фиссуру I, а вторая – в центральную ямку. Их терминальные отрезки, пролегая по границе окклюзивной плоскости, меняют направление хода к вершине осевого гребня (рис. 3). Таким образом, указанные борозды выделяют элементы краевого гребня вестибулярной поверхности (эокристы), соответствующие его центральной ветви, которые у современного человека не встречаются. В районе границы параконуса и метаконуса этот гребень прерван фиссурой I и короткой дублирующей бороздкой 1’pa.
Метаконус не редуцирован. По размерам мезио-дистального диаметра он не уступает параконусу. На его поверхности дифференцируются осевой, мезиальный и дистальный сегменты. Осевой сегмент очень массивен. В дистальном отделе он разделен на отдельные фрагменты дополнительными поперечными бороздками. Дистальный и мезиальный сегменты метаконуса четко выражены только на вестибулярном крае коронки.
Плагиокриста прервана центральной бороздой почти полностью, только в дистальном отделе осталась тонкая эмалевая перемычка, пролегающая параллельно фиссуре IV.
Терминальные отрезки первой и второй борозды метаконуса выходят на вестибулярную поверхность. Они формируют самостоятельные вершины мезиального и дистального сегментов бугорка. Борозда 1me
Материалы, привлеченные для сравнения
|
Регион |
Таксономический статус находки * |
Название памятника |
Номер образца |
Источник данных |
|
1 |
2 |
3 |
4 |
5 |
|
Африка |
Anatomically modern Homo sapiens |
Фиш Хук |
– |
Schwartz, Tattersall, 2003, vol. II |
|
Homo helmei |
Флорисбад |
– |
Ibid. |
|
|
Homo habilis |
Хадар |
A.L. 666 |
» |
|
|
Homo heidelbergensis |
Кабве |
– |
» |
|
|
Homo erectus |
Кооби-Фора |
KNM-ER 1813, KNM-ER 3733 |
» |
|
|
Homo ergaster |
Нариокотоме |
KNM-WT 15000 |
Халдеева, Зубов, Харламова, 2012 |
|
|
Homo habilis |
Ущелье Олдувай |
OH 6, OH 13, OH 16, OH 24 |
Schwartz, Tattersall, 2003 |
|
|
Homo erectus ? Ath-lanthropus mauritani-cus (?) |
Томас Квори (Рабат) |
– |
Ibid. |
|
|
Homo erectus |
Консо |
KGA 4-14, KGA 11-350 |
Suwa et al., 2007 |
|
|
Early Homo sapiens |
Дар-эс-Солтан |
DS II – H9, DS II – NN, DS II – H5, DS II – H10 |
Hublin et al., 2012 |
|
|
» |
Пещера контрабандистов |
Ctb H7, Ctb Ib 19, Ctb T4, Ctb T3b |
Ibid. |
|
|
Западная Азия |
Early Homo sapiens |
Джебель Кафзех |
Qafzeh 4, Qafzeh 5, Qafzeh 6, Qafzeh 9, Qafzeh 11 |
Schwartz, Tattersall, 2003, vol. II |
|
Homo neanderthalensis |
Схул |
Skhul IV, Skhul V |
Ibid. |
|
|
» |
Табун |
Tabun I, Tabun T I, Tabun T II |
» |
|
|
» |
Кебара |
KNM 24, KNM 21 |
Tillier et al., 2003 |
|
|
Early Homo sapiens |
Кесем |
– |
Hershkovitz et al., 2011 |
|
|
Восточная и Централь- |
Archaic Homo sapiens |
Цзиннюшань |
– |
Schwartz, Tattersall, 2003, vol. II |
|
ная Азия |
Homo sapiens |
Люцзян |
– |
Ibid. |
|
Homo erectus |
Сангиран |
Sangiran 4, 7, 17, 27, NG 91-G10 No1, NG 0802.1, NG 0802.3, NG 92.3, Njg 2005.05, Bpg 2001.04, PDS0712, NG0802 |
Ibid.; Zanolli, 2013; Kaifu et al., 2007; Zaim et al., 2011 |
|
|
Homo sapiens |
Ваджак |
Wajak 1, Wajak 2 |
Schwartz, Tattersall, 2003, vol. II |
|
|
Homo erectus |
Чжоукоудянь, нижний грот |
ZKD 169.25, ZKD PA 327, ZKD skull XI |
Ibid. |
|
|
Homo sapiens |
Чжоукоудянь, верхний грот |
PA 101, PA 102, PA 103 |
Ibid.; Turner, Manabe, Hawkey, 2000 |
|
|
Early Homo |
Суйцзиайяо |
PA1480, PA1481, PA 1500 |
Xing et al., 2015 |
|
|
Homo sapiens |
Даосянь |
DX 1, 4, 5, 6, 8, 12, 14, 16, 17, 20, 21, 24, 28, 31,33, 35, 36, 39, 41, 47 |
Liu et al., 2015 |
|
|
Homo erectus |
Лянг Буа |
LB1 |
Kaifu et al., 2015 |
|
|
Северная Азия |
Homo sapiens |
Мальта |
1, 2 |
Зубов, Гохман, 2003; Зубова, Чикишева, 2015 |
|
Homo neanderthalensis |
Чагырская |
10, 51.1, 57 |
Неопубликованные данные А.В. Зубовой |
|
|
Средняя Азия |
Homo neanderthalensis |
Оби-Рахмат |
– |
Glantz et al., 2008; неопубликованные данные А.В. Зубовой |
|
» |
Тешик-Таш |
– |
Неопубликованные данные А.В. Зубовой |
Окончание таблицы
|
1 |
2 |
3 |
4 |
5 |
|
Европа |
Homo sapiens |
Абри Пату |
Pataud 1 |
Schwartz, Tattersall, 2003, vol. I |
|
Homo heidelbergensis |
Араго |
Arago 21, 14, 31 |
Ibid. |
|
|
Homo antecessor |
Атапуэрка: Гран Долина |
ATD 6-69 |
» |
|
|
Homo heidelbergensis / antecessor |
Атапуэрка: Сима де Лос Уэсос |
AT-16, AT, 3177, AT-138, AT-406, AT-139, AT-26, AT-959, AT-20, AT-2076, AT-812, AT-944, AT-196, AT-2071, AT-4317, AT-3424, AT-587, AT-46, AT-4326, AT-960, AT-824, AT-2179, AT-407, AT-4319, AT-4336, AT-12, AT-2175, AT-815, AT-821, AT-15, AT-170, AT-602, AT-816, AT-274, AT-3181, AT-171, AT-826, AT-601, AT-945, AT-1471, AT-2393, AT-3183, AT-194, AT-5082, AT-2150, AT-140 |
Martinón-Torres et al., 2012 |
|
|
Homo erectus |
Дманиси |
D 2882, D 2700 |
Schwartz, Tattersall, 2003; Martinón-Torres et al., 2008 |
|
|
Homo sapiens |
Энгис |
Engis 2 |
Schwartz, Tattersall, 2003, vol. I |
|
|
» |
Гримальди |
Barma grande 2 |
Ibid. |
|
|
» |
Истюриц |
Ist 71 |
» |
|
|
Homo neanderthalensis |
Крапина |
45, 46, 47, 48, D 119, D 120, D 170, D 180, D 136, D 164, D 178, D 188 |
Radovčić et al., 1988 |
|
|
» |
Ля Кина |
H 5, H 18 |
Schwartz, Tattersall, 2003, vol. I |
|
|
» |
Ле Мустье |
– |
Ibid. |
|
|
Homo sapiens |
Младеч |
1, 2 |
» |
|
|
Homo neanderthalensis |
Пеш де л’Азе |
– |
» |
|
|
» |
Саккопасторе |
2 |
» |
|
|
» |
Сакаджия |
– |
» |
|
|
» |
Складина |
– |
» |
|
|
» |
Спи |
1, 2 |
» |
|
|
» |
Шубалюк |
2 |
» |
|
|
Homo sapiens |
Ахштырская |
– |
Неопубликованные данные А.В. Зубовой |
|
|
» |
Рожок-1 |
– |
То же |
|
|
» |
Кальдерао |
1 |
Trinkaus, Bailey, Zilhao, 2001 |
|
|
» |
Сунгирь |
2, 3 |
Зубов, 2000 |
|
|
» |
Костёнки |
14, 15, 17, 18 |
Халдеева, 2006; неопубликованные данные А.В. Зубовой |
|
|
» |
Визольяно |
6 |
Abazzi et al., 2000 |
|
|
» |
Галерея да Цистерна |
– |
Trinkaus et al., 2011 |
|
|
Early Homo |
Пештера-ку-Оасе |
2 |
Trinkaus, 2010 |
*Поскольку таксономический статус некоторых находок является дискуссионным, в этом столбце информация приводится в соответствии с точкой зрения авторов, публикации которых использованы в данной статье.
впадает в фиссуру I чуть ниже 1pa, 2me сливается с бассейном талона. От осевого гребня метаконуса в дистальном направлении отходят два дополнительных эмалевых гребня. Они идут на контакт с дополнительными бугорками дистального края коронки, но прерываются бассейном талона. Эти гребни разделяет дополнительная бороздка третьего порядка, выделяющая элементы метаконулюса на осевом гребне и не имеющая строгого соответствия со стандартными элементами одонтоглифической схемы моляров современного человека [Зубов, 2006; Зубов, Халдеева, 1989]. По аналогии с другими бугорками она обозначена как 4me. В отделе, прилежащем к центральной ямке, осевой сегмент метаконуса разделяется в продольном направлении еще одной дополнительной бороздкой третьего порядка (3me), которая берет начало примерно в средней трети гребня и впадает в фиссуру III.
Гипоконус крупный, подовальной формы, вытянутый в вестибуло-лингвальном направлении. На его поверхности резко выделяется массивный центральный гребень. Мезиальный сегмент бугорка очень тонкий, выделяющая его борозда практически сливается с фиссурой IV. Дистальный сегмент морфологически выражен несколько сильнее. На окклюзивной поверхности гипоконус отделяется от метаконуса и протоконуса широким и протяженным бассейном талона, который сливается с элементами задней ямки на метаконусе. В дистальном отделе гипоконус отграничен от метаконуса округлым дополнительным бугорком на краевом гребне (С5 балл 1 ASUDAS). Обе главные борозды гипоконуса (1hy и 2hy) впадают в бассейн талона. Они небольшие по протяженности, но значительные по глубине и дублируются в мезиальном и дистальном отделах параллельными бороздками третьего порядка, которые крайне редко встречаются у современного человека. По аналогии с такими же бороздками других бугорков их можно обозначить 1’hy и 2’hy. В области вершины бугорка выделяется элемент краевого гребня, как на метаконусе и параконусе.
Протоконус массивен. Он является самым крупным бугорком коронки. Во многом это обусловлено увеличением его базальной части за счет бугорка Карабелли, который, соприкасаясь с мезио-лингвальной бороздой, разделяющей гипоконус и протоконус, занимает все основание протоконуса в цервикальной части (см. рис. 1). В средней трети высоты протоконуса этот бугорок имеет четыре вершины в виде цин-гулярных гребней (см. рис. 2) и таким образом демонстрирует более полную, чем у современного человека, реализацию, потенциально заложенную в геноме модели верхнего моляра. Множественные вершины в этом случае являются проявлением рудиментарных дериватов цингулюма, относимых по терминологии П. Хершковица к общей категории энтостилей [Hersh-kovitz P., 1971].
Соотнести наблюдаемый морфологический вариант с баллами стандартных шкал до статочно трудно, поскольку ни одна из них не предполагает многовер-шинности бугорка. В системе ASUDAS стадия, на которой бугорок Карабелли вступает в контакт с межбугорковой бороздой, соответствует баллу 5, а по шкале А.А. Зубова – баллу 4.
Протоконус, как и другие основные бугорки коронки, сегментирован на три части. Самым крупным является центральный сегмент, следующим по размеру – мезиальный и самым маленьким – дистальный. Вторая борозда протоконуса (2pr) соединяется с 2’pr, формируя изолированный трирадиус. На терминальном отрезке 2’pr в нее впадает еще одна борозда третьего порядка – 2’’pr. Борозда 1pr редуцирована, дистальный сегмент выделяется бороздой 1’pr, которая имеет вид ямки, изолированной от фиссур III и IV тонким эмалевым мостиком. Он соединяет дистальный сегмент протоконуса с дистальной частью осевого гребня метаконуса и таким образом формирует непрерывный элемент плагиокристы (см. рис. 3).
Протоконус и параконус разделены дополнительным мезиальным бугорком. Он образован дополнительными бороздками 2’ра и 2’pr, впадающими в фиссуру II. У современного человека этот бугорок, как правило, круглой формы и формируется концевым трирадиусом фиссуры II строго на мезиальном краевом гребне. В данном случае бугорок имеет форму сегмента, располагающегося параллельно фиссуре II почти до центральной ямки. Трирадиус смещен в центральную треть межбугорковой фиссуры, ввиду этого сильно укороченной. «Веточки» трирадиуса приобретают сходство с основными туберкулярны-ми бороздами. Эпикриста прервана. Передняя ямка отсутствует, затека эмали на вестибулярной стороне зуба нет.
Изначально зуб имел три корня – лингвальный, мезио-буккальный и дисто-буккальный (рис. 4). Лингвальный корень, самый длинный и массивный, был разрушен в процессе палеогенетического исследования. В сечении он овальной формы с уплощением в букко-лингвальном направлении, от буккальных корней отходит под очень большим углом. С дистальным корнем лингвальный расходится на уровне 3,6 мм от эмалево-цементной границы, с мезиальным – на уровне 4,1 мм.
Корни буккальной стороны разделяются только в нижней трети на расстоянии 8,2 мм от эмалево-цементной границы. До этого уровня они соединены цементной пластинкой. Мезиальный корень в средней трети изогнут. Он уплощен в мезио-дистальном направлении; при этом его вестибулярный отдел массивнее, чем лингвальный. Дистальный корень по сравнению с другими наименее массивен, почти плоский в сечении.
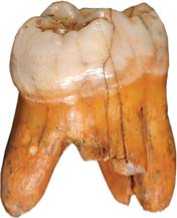
Рис. 1. Дисто-вестибуляр-ная норма Денисова 4.

Рис. 2. Окклюзальная норма Денисова 4.
Левый верхний постоянный третий моляр (М3) из подошвы слоя 11.4 на контакте со слоем 12 – Денисова 8 (рис. 5). Верхний левый моляр индивида мужского пола [Slon et al., 2015], который был чуть старше, чем Денисова 4 . После реставрации коронки появилась возможность оценить ее контуры. Она округлой формы, без скошенности мезио-вестибуляр-ного отдела, но с небольшим расширением в вестибулярной части, по сравнению с лингвальной (рис. 6).
Протоконус, несмотря на посмертные разрушения, выглядит довольно крупным. Осевой гребень бугорка очень широкий, вместе с осевым гребнем мезиальной части метаконуса он формирует широкий гребень плагиокристу, непрерывность или дискретность которого полностью установить невозможно. Борозда 1pr стерта. Борозда 2pr, видимо, была более глубокой, поскольку, несмотря на значительную стертость мезиального отдела коронки, можно установить направление ее хода. Она рассекала краевой гребень протоконуса и выходила на вестибулярную поверхность коронки (рис. 7).
Метаконус очень массивный, намного крупнее параконуса и гипоконуса. Он разделяется на две части. В состав одной части входит осевая часть бугорка с редуцированным мезиальным сегментом, в состав другой – дистальный сегмент и дистальный краевой
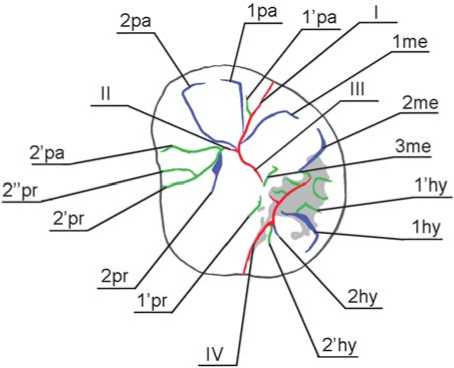
Рис. 3. Одонтоглифическая схема Денисова 4.
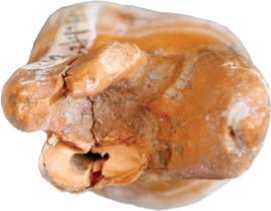
Рис. 4. Корневая система Денисова 4.
гребень. Борозда 1me стерта. Разделяющая две части бугорка борозда 2me значительна по протяженности и глубине, по морфологии приближается к межбугорковым фиссурам. В дисто-вестибулярном отделе коронки она рассекает краевой гребень и выходит далеко на внешнюю поверхность стенки зуба, продолжаясь до нижней ее трети. В центральной части борозда 2me впадает в фиссуру III. Осевой гребень метаконуса Денисова 8, как и Денисова 4, очень массивен. С дистальной стороны он разделен дополнительными
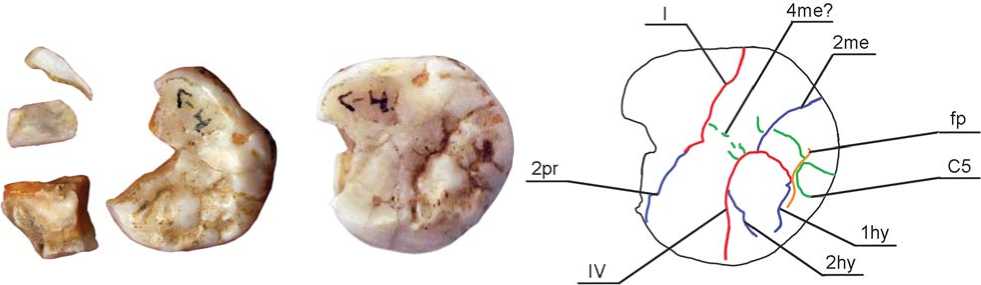
Рис. 7. Одонтоглифическая схема Денисова 8.
Рис. 5. Моляр Денисова 8 до реставрации.
Рис. 6. Моляр Денисова 8 после реставрации.
бороздками 4me и 4’me, выделяющими в его структуре элементы метаконулюса (рис. 7).
На бугорке, образованном дистальной частью метаконуса, выделяется массивный гребень, который, вступая в контакт с осевым гребнем гипоконуса, визуально формирует структуру, параллельную плагиокристе, но прерванную продольной бороздой, разделяющей метаконус и гипоконус. Гребень разделен на две части дополнительной бороздкой, впадающей с одной стороны в 2me, а с другой – в заднюю ямку, ограничивающую гребень дистально и отделяющую его от дистального краевого гребня. На вестибулярной поверхности метаконуса в районе границы между ним и параконусом имеется сформированный мезиостиль. Затек эмали на вестибулярной стороне зуба отсутствует.
Параконус заметно меньше метаконуса. По мезио-дистальному размеру он визуально соответствует мезиальной его части. Ввиду сильной стерто сти и посмертных разрушений описать строение параконуса сложно. Можно только предположить наличие у него дополнительной бороздки 4ра, которая рассекала осевой гребень параллельно фиссуре II.
Гипоконус заметно редуцирован, не формирует дисто-лигвальный угол коронки, меньше метаконуса и протоконуса и не так сильно морфологически выражен, как у индивида Денисова 4. По шкале А. Даль-берга, используемой в одонтологической системе А.А. Зубова, гипоконус соответствует баллу 4, по шкале ASUDAS – баллу 5 [Turner, Nichol, Scott, 1991]. Бугорок имеет форму сегмента с вершиной в точке пересечения фиссур III и IV, морфологически сильно дифференцирован. На его поверхности выделяются три гребня – мезиальный, центральный и дистальный маргинальный. Дистальный сегмент основной части бугорка редуцирован. Мезиальный гребень очень тонкий и короткий, имеет вид тонкого эмалевого гребешка, отходящего от края лингвальной поверхности в вестибулярном направлении. Борозда 2hy, отделяющая его от осевого гребня, сливается с межбугорковой фиссурой IV вблизи лингвального края коронки (рис. 7). Осевой гребень гипоконуса исключительно массивен. Он пролегает в вестибуло-лингвальном направлении параллельно косому гребню (плагиокрис-те). Оформляющая его в дистальном отделе борозда 1hy практически сливается с задней ямкой, отделяющей осевой гребень гипоконуса от дистального краевого. Задняя ямка очень узкая и имеет вид глубокой борозды, прерванной дополнительным дистальным бугорком (баллы 3–4 ASUDAS). Фиссура, разделяющая гипоконус и протоконус, в лингвальном отделе коронки не выходит на лингвальную поверхность, как это обычно бывает, а заканчивается на поверхности краевого гребня. В месте пересечения окклюзивной и лингвальной поверхностей зуба сохраняется тонкая непрерывная перемычка между гипоконусом и протоконусом. Ее можно отождествить с рудиментом архаичного гребня энтокристы, который имеется на зубах приматов, а у современного человека – только на закладках молочных вторых моляров [Зубов, Халдеева, 1989].
Корневая система зуба разрушена, но рельеф сохранившихся фрагментов позволяет сделать предположение о наличии трех корней – лингвального, дистального и мезиального.
О таксономическом статусе одонтологического комплекса верхних моляров денисовца
Оба постоянных моляра из Денисовой пещеры характеризуются рядом общих признаков, что позволяет говорить о едином таксономическом статусе находок. Наиболее ярко их специфику отражают крупные размеры коронок с округлыми углами; отсутствие выраженной редукции дистальных зубов в ряду моляров; исключительно сильная морфологическая выраженность борозд первого и второго порядка, часто выходящих на стенки коронки или формирующих дополнительные бугорки; наличие дополнительных борозд третьего порядка, отсутствующих у современного человека. На обоих зубах имеются цингулярные образования, представленные множественными энто-стилями в комплексе Карабелли, дополнительными маргинальными бугорками, мезиостилем верхних моляров. К этому же ряду признаков следует отнести выраженную тенденцию к гребнеобразованию на окклюзивных поверхностях о сновных бугорков. Она проявляется в формировании широкой и сложной плагиокристы, в дроблении метаконуса с выделением гребня, параллельного плагиокристе; в сегментации основных бугорков, сохранении энтокристы и элементов эокристы.
Согласно результатам сравнительного анализа, наблюдаемая комбинация признаков не соответствует характеристикам ни одного из европейских таксонов, привлеченных для сравнения. Большинство признаков в ее составе – маркеры глубокой архаики. При этом в данном одонтологическом комплексе отсутствуют неандертальские признаки [Халдеева, Харламова, Зубов, 2010; Bailey, 2002], которые могли бы указывать на близость изучаемой группы к таксону H. heidelber-gensis, морфологическую специфику которого определяют повышенные частоты архаичных признаков в сочетании с неандертальскими чертами [Martinón-Torres et al., 2012]. В рассматриваемой комбинации не представлены такие маркеры сапиентной линии, как сильная редукция гипоконуса второго верхнего моляра и редукция борозд третьего порядка. Из от-но сительно прогрессивных характеристик зубов де- нисовца можно отметить только заметную редукцию гипоконуса на верхнем третьем моляре. О наличии апоморфных признаков пока судить преждевременно.
В целом морфологические особенности моляров из Денисовой пещеры представляют модель, эволюционно соответствующую стадии архантропа. Результаты сравнительного анализа позволяют говорить о ее наибольшем сходстве с одонтологическими комплексами, распространенными у H. erectus ЮгоВосточной Азии, точнее, у эректусов сангиранской группы. Их полиморфизм чрезвычайно высок, однако в Сангиране встречаются мегадонтные образцы, сопоставимые по размерам с зубами денисовских гомини-нов [Lovejoy, 1970, tab. 2; Orban-Segebarth, Procureur, 1983, tab. 2; Tyler, 2001]. На сангиранских образцах зафиксирован почти полный набор архаичных признаков, характерный для денисовцев. У сангиранских гомининов повышена частота цингулярных дериватов – зубцов эктостилей и эктостилидов вестибулярной и лингвальной поверхности верхних и нижних моляров и дополнительных бугорков – NG 8503 [Kaifu, Aziz, Baba, 2005], Sangiran 5, 6, 7, 9 [Schwartz, Tattersall, 2003, Vol. II], Sangiran 8 [Kaifu, Aziz, Baba, 2005], Sangiran 22, 27, 33 [Ibid.], Bpg 2001.4 [Zaim et al., 2011]. Во многих случаях у гомининов из Сангирана так же ярко, как у денисовцев, выражена тенденция к гребнеобразованию, хотя по опубликованным данным невозможно судить о сохранении у яванских находок энтокристы и эокристы, а также о присутствии архаичных одонтоглифических вариантов. На образце Bpg 2001.4 отмечена задняя ямка в виде Т-образной фиссуры [Ibid.]. У Sangiran 4, 27, Bpg 2001.04 отмечены похожие трапециевидные с округленными углами очертания коронок верхних моляров [Schwartz, Tattersall, 2003, vol. II; Zaim et al., 2011].
Второй ареал комплексов, сходных по составу морфологических характеристик с денисовским, был зафиксирован на территории Северного Китая – местонахождение Суйцзиайяо, датируемое концом среднего – началом верхнего плейстоцена [Xing et al., 2015]. Для верхних постоянных моляров из Суйцзи-айяо характерны очень крупные размеры, подквадратная форма коронки, дополнительные мезиальные и дистальные маргинальные бугорки, сложная форма бугорка Карабелли, дифференцированный одонтогли-фический узор, у которого фиксируется экстремальная выраженность главных туберкулярных борозд, и тенденция к дроблению метаконуса и гипоконуса [Ibid., fig. 2]. На верхних молярах РА 1481 и РА 1500 отмечено исключительно сильное расхождение вестибулярных корней с лингвальным [Ibid., fig. 3], подобное наблюдается у индивида Денисова 4. Антропологические находки из Суйцзиайяо характеризуются мозаичностью морфологического облика, имеют значительные отличия от китайских одонтологических образцов, относящихся к ранним современным людям, и сохраняют архаичный компонент, сближающий их со среднеплейстоценовыми гомининами Восточной Азии – Сангиран, Чжоукоудянь, Лонтандун, Чао-сан. Из маркеров неандертальского комплекса на зубах из Суйцзиайяо отмечены только неспецифические признаки, довольно широко представленные в сериях среднеплейстоценовых гомининов [Ibid., р. 237–238].
Таксономический статус находок из Суйцзиайяо по отношению к H. sapiens и H. neanderthalensis пока не определен. При этом значительное сходство в строении верхних моляров из Суйцзиайяо и Денисовой пещеры позволяет предположить, что их обладатели относятся к одному таксону, характеризующемуся длительным сохранением эректоидных признаков. Если дальнейшие исследования подтвердят это сходство, то оно станет серьезным аргументом в пользу гипотезы о широком распространении денисовцев на территории Восточной Азии [Reich et al., 2010; Деревянко, 2011]. Вместе с тем необходимо отметить, что на других палеоантропологических находках из Китая комплекс архаичных признаков, объединяющий сангиранских эректусов и денисовцев, заметно редуцирован или отсутствует [Turner, Manabe, Hawkey, 2000; Wu, Poirier, 1995; Schwartz, Tattersall, 2003, Vol. II; Xing, Zhou, Liu, 2009; Liu et al., 2010].
О возможности длительной консервации морфологической специфики эректоидных форм свидетельствуют останки H. floresiensis [Brown et al., 2004] – вида, сформировавшегося под влиянием длительной изоляции в островных условиях. Морфологическая специфика денисовских находок и гомини-нов из Суйцзиайяо позволяет предполагать наличие на территории Азии еще одного очага консервативного морфогенеза.
Наблюдаемое одонтологическое сходство между верхнепалеолитической популяцией Алтая, среднеплейстоценовыми гомининами Китая и нижнепалеолитическим населением Юго-Восточной Азии не противоречит результатам палеогенетических исследований. Период, в который произошло расхождение предков денисовского человека с общим предком H. sapiens и H. neanderthalensis , совпадает с наиболее поздними датами, полученными для находок H. erectus на Яве [Pope, Cronin, 1984]. На территории Алтая близкий возраст имеет стоянка Карама [Bolikhovskaya, Derevianko, Shunkov, 2006]. Таким образом, миграция, обусловившая сходство денисовских гомининов и H. erectus Явы, допустима. В этой связи следует отметить, что комплекс архаичных морфологических особенностей в строении зубочелюстного аппарата более отчетливо выражен у поздних H. erectus Явы, чем у ранних [Kaifu et al., 2005].
Наибольший вклад денисовского человека в генофонд современных популяций прослеживается также на юго-востоке Азии, в Меланезии [Reich et al., 2010], Папуа – Новой Гвинее, Полинезии, на Фиджи [Reich et al., 2011]. Исследования показали невозможность смешения базовых геномов современных групп с денисовским на северо-западе или западе азиатского континента. Скорее всего, очаг смешения находился непосредственно на территории Юго-Восточной Азии [Ibid., p. 523]. Палеогенетические данные позволяют также предполагать, что в этом регионе до появления предков современного населения денисовские гены были распространены достаточно широко.
На основе полученных результатов можно сделать два вывода. Первый – при формировании H. altaiensis определяющее значение имела консервация архаичных компонентов без вытеснения их прогрессивными трансформациями. Морфологическую специфику таксона определяет комплекс характеристик эректоидно-го облика, который полностью представлен на обоих молярах, несмотря на значительную давность расхождения геномов индивидов, которым они принадлежали. Согласно палеогенетическим данным, денисовская популяция характеризовалась крайне низким уровнем генетического разнообразия [Meyer et al., 2012; Slon et al., 2015], что отличает денисовский таксон от современного человека, особенностями которого являются максимально широкая адаптивная радиация и высокий уровень генетического разнообразия. В этом отношении денисовцы ближе к неандертальцам, которые представляли собой специализированную форму с более низким, чем у современного человека, уровнем генетического разнообразия [Reich et al., 2010, p. 1055].
Второй вывод – специфический морфологический комплекс денисовских моляров находит аналоги только на территории Азии. На европейских образцах подобные комплексы не обнаружены. Таким образом, генезис H. altaiensis , вероятнее всего, связан с азиатскими эректоидными формами, что соответствует предположениям, выдвинутым на основании изучения археологических материалов [Деревянко, 2011].
Доминирование эректоидных признаков в морфологическом комплексе моляров из Денисовой пещеры позволяет полагать, что часть генома денисов-цев, принадлежавшая гомининам неустановленного вида [Krause et al., 2010], маркирует таксон H. erectus. К этому предположению, несомненно, следует относиться с большой осторожностью, тем более что в своем большинстве архаичные признаки в морфологии сангиранских эректусов не являются для них апо-морфными, они унаследованы от более древних форм, относящихся к таксонам H. habilis и H. rudolfensis . Однако морфологические свидетельства сохранения наследия азиатских представителей H. erectus в антропологии алтайских гомининов сегодня выглядят вполне убедительными.
Заключение
Результаты проведенного исследования еще раз показали высокую значимость одонтологических признаков для диагностики межвидовых различий в пределах рода Homo . Они подтвердили существование на территории Алтая особой популяции, обозначенной как H. altaien-sis , которая отличается от H. sapiens и H. neandertha-lensis не только генетически, но и морфологически. Одонтологическая специфика этой группы заключается в мегадонтии и длительной консервации маркеров, сближающих денисовца со среднеплейстоценовыми гомининами Северного Китая и эректусами Сангирана.
В эволюционном отношении H. altaiensis, таким образом, характеризуется крайне консервативной моделью морфогенеза.
Анализ морфологии верхних моляров денисовца позволил не только подтвердить равную значимость генетических и морфологических критериев при выделении новых форм гомининов, что является предметом активной дискуссии в палеоантропологической науке, но и сделать морфологически обоснованное предположение о таксономической принадлежности неидентифицированной части денисовского генома представителям вида H. erectus s.l . Это предположение подчеркивает самостоятельную ценность одонтологической информации, получаемой классическими методами исследования, при доступности методов молекулярно-генетического анализа.
Список литературы Морфологическая характеристика постоянных моляров из палеолитических слоев Денисовой пещеры
- Деревянко А.П. Верхний палеолит в Африке и Евразии и формирование человека современного анатомического типа. -Новосибирск: Изд-во ИАЭТ СО РАН, 2011. -560 с.
- Зубов А.А. Одонтология. Методика антропологических исследований. -М.: Наука, 1968. -199 с.
- Зубов А.А. Одонтоглифика//Расогенетические процессы в этнической истории. -М.: Наука, 1974. -С. 11-42.
- Зубов А.А. Морфологическое исследование зубов детей из Сунгирского погребения 2//HOMO SUNGIRENSIS. Верхнепалеолитический человек: экологические и эволюционные аспекты исследования. -М.: Науч. мир, 2000. -С. 256-268.
- Зубов А.А. Методическое пособие по антропологическому анализу одонтологических материалов. -М.: Этно-Онлайн, 2006. -72 с.
