Морфологические и цитологические исследования искусственных полиплоидов кориандра Coriandrum sativum L

Автор: Мошненко Е.В., Зеленцов С.В., Пасменко Т.В., Лунва В.Б.
Статья в выпуске: 2 (141), 2009 года.
Бесплатный доступ
Изучены морфологические особенности тетраплоидов кориандра. Установлено, что тетраплоидные растения отличаются увеличенными размерами листовых пластинок прикорневых листьев и увеличенной длиной черешков. Репродуктивные органы ранних (С1-С2) тетраплоидов характеризуются частичной стерильностью цветков, уменьшенным количеством пыльцы и её сниженной фертильностью. Завязываемость плодов в зонтиках тетраплоидов понижена. Тетраплоидные плоды имеют более крупные размеры по сравнению с диплоидными формами. После восстановления и стабилизации фертильности, сохранившие крупнолистность и крупноплодность тетраплоидные, либо реплоидные формы кориандра могут быть использованы в качестве исходного материала для селекции кориандра на эти признаки.
Кориандр, полиплоидия, пыльца, колхицин
Короткий адрес: https://sciup.org/142150895
IDR: 142150895
Morphological and cytological researches of artificial polyploids of coriander Coriandrum sativum L
Morphological features of coriander tetraploids are studied. It is established that tetraploid plants have the increased sizes of leaf blades of composite leaves and increased petioles length. Reproductive organs of early (С1-С2) tetraploids are characterized by partial flower sterility, reduced quantity of pollen and its lowered fertility. Seedsetting rate in tetraploid umbels is lowered. Tetraploid fruits have larger sizes in comparison with diploid forms. After restoration and stabilization of fertility, tetraploid and reploid coriander forms kept large-leaves and large-fruitiness can be used as a parental material for coriander breeding on these traits.
Текст научной статьи Морфологические и цитологические исследования искусственных полиплоидов кориандра Coriandrum sativum L
Введение . Кориандр ( Coriandrum sativum L. (2n = 22) – одна из древнейших эфиромасличных культур, которая относится к семейству зонтичных – Umbelliferae (син.: Apeaceae). Ценность кориандра заключается в высоком содержании эфирного и жирного масел в плодах, которые широко применяются в парфюмерно-косметической и пищевой промышленности. Название «кориандр» происходит от греческого слова " koris ", что означает «клоп», так как в незрелом состоянии растение издаёт запах клопа. Однако к созреванию дециловый альдегид, который и определяет этот запах и содержание которого составляет до 60-80 % в эфирном масле зелёных частей растения, почти полностью выветривается, и зрелые плоды приобретают приятный пряный аромат [Столетова, 1931; Порембский, 2000].
Кориандровое эфирное масло содержит свыше 20 компонентов, среди них терпеновые спирты: линалоол (60-80 %), гераниол (3-5 %), борнеол (1-4 %), а также геранилацетат (до 50 %), уксуснокислые эфиры, альдегиды и терпены. Линалоол служит исходным продуктом для получения ряда душистых веществ, обладающих запахами лимона, апельсина, розы, фиалки и др. Среднее содержание эфирного масла в плодах кориандра, выращенного в европейских странах, варьирует от 0,5 до 0,9 %, по другим сведениям содержание эфирного масла в плодах может варьировать от 0,03 до 2,6 и даже 3,0 %. Велика также ценность кориандра как медоносного растения. В связи с таким многообразием использования кориандра на протяжении длительного времени к нему сохраняется устойчивый интерес в нашей стране и за рубежом. Посевная площадь, занятая под этой культурой в мире, в настоящее время охватывает 300-320 тыс. га. Из этой площади около 15-20 тыс. га засеваются для получения зелени, а 280-300 тыс. га на – зерно [10, 13, 16, 11].
Основным направлением возделывания этой культуры является выращивание для получения плодов, которые используются в пищевой промышленности и для производства кориандрового масла. По данным британского Каталога эфирных масел, среди экспортируемых стран на международном рынке высоко ценится российский кориандр вследствие большого содержания в нём эфирного масла, которое добывают из плодов. Современные отечественные сорта кориандра способны формировать урожай плодов до 2,0 т/га и накапливать до 3 % эфирного масла, обеспечивая его выход до 45-50 кг/га [8, 4, 6].
Наиболее распространённым методом создания исходного материала у кориандра является внутривидовая гибридизация с последующим индивидуальным отбором элитных растений и оценкой по потомству. Для селекции кориандра на скороспелость, высокую продуктивность, повышенное содержание эфирного и жирного масел, устойчивость к болезням, полеганию растений и осыпанию плодов широко используется мировой генофонд культуры [12, 16, 14].
Следует отметить, что и в отечественной, и в зарубежной селекции кориандра ранее практически не использовался такой метод получения исходного материала, как полиплоидия. При этом общепринятым является представление о полиплоидии, как самой стремительной форме эволюции, обеспечивающей наиболее быстрые наследственные изменения в геноме. Ещё в 70-е годы прошлого века многие генетики и селекционеры высказывали единодушное мнение о целесообразности использования искусственных полиплоидов, предоставляющих богатый исходный материал для селекционной работы [18, 17]. Кроме этого, известно, что автополиплоиды, как правило, нестабильны, и в последующих поколениях из них часто выщепляются растения с более низким уровнем плоидно-сти – анеуплоиды и возвратные диплоиды (реплоиды), в том числе несущие изменённые или даже новые хозяйственно ценные признаки [2]. При этом выделенные фертильные реплоиды также могут быть использованы в практической работе по дальнейшему расширению полиморфизма признаков, например, на основе разработанной во ВНИИМК теории полиплоидной рекомбинации генома высших цветковых растений [3, 5].
Однако на кориандре известны лишь отдельные работы, посвящённые некоторым аспектам полиплоидии [15]. В связи с этим целью нашей работы было изучение возможности искусственной полиплоидизации кориандра на Алексеевской ОС для получения исходного материала с улучшенными хозяйственно ценными признаками и их дальнейшего применения в селекционной работе.
Материал и методы . Исследования проводили в 2007-2009 гг. в полевых условиях и в камере искусственного климата. Фотопериод в камере искусственного климата составлял 16 ч с интенсивностью освещения ≈8000 лк.
Материалом для искусственной полиплоидизации служили 6 сортов кориандра Янтарь, А-1820, А-1366, А-413, А-190, ЭВА-1. Плоидность сортов увеличивали двукратной обработкой верхушечных меристем вегетирующих растений в фазу образования центрального зонтика водным раствором колхицина с концентрацией 0,05 % [1]. Контролем служили необработанные растения. Плоды с морфологически изменённых растений проращивали и в корешках подсчитывали количество хромосом. Проростки с удвоенной плоидностью высаживали в сосуды и выращивали до созревания. Цитологический анализ плоидности, а также фертильности пыльцы диплоидных и полиплоидных форм кориандра проводили при увеличении X500-1000 на световом бинокулярном микроскопе. Простые зонтики фиксировали в упрощённом фиксаторе Карнуа (3:1). Для приготовления и окрашивания временных давленых препаратов использовали стандартную методику [7]. Временные препараты окрашивали ацетолакмоидом.
Результаты и обсуждение . Ювенильные растения кориандра были обработаны растворами колхицина в полевом сезоне 2007 г. Рост и развитие колхицинированных меристем были замедленными. Общая высота таких растений после завершения цветения зонтиков оказалась в 2 раза ниже по сравнению с исходными необработанными формами. Симметрия лучей зонтиков на них была нарушенной, а фертильность цветков – сниженной. Эти морфологические отличия от контрольных растений и послужили критерием для их отбора и последующего изучения.
Визуальный анализ позволил установить, что тетраплоидные растения С 1 всех исследуемых сортов отличались более тёмной окраской розеточных листьев, а также более широкими листовыми пластинками и утолщёнными черешками по сравнению с исходными диплоидными формами (рис. 1).

Рисунок 1 – Морфологические особенности первых розеточных листьев тетраплоидного кориандра на 30-й день после всходов, сорт А-1366
(камера искусственного климата)
При формировании последующих прикорневых листьев было обнаружено, что у тетрапло-идных растений они состоят из увеличенного количества листовых пластинок. Кроме этого, по мере роста у них заметно удлинялись черешки. В результате у тетраплоидов общая длина листа вместе с черешками возросла в полтора раза по сравнению с исходной формой (рис. 2 а и б ).

Рисунок 2 – Морфологические особенности тетраплоидов кориандра С 1 в фазе розетки, 50-е сутки после всходов, сорт А-1366
(камера искусственного климата) а – исходная диплоидная форма кориандра, 2n = 22; б – тетраплоидная форма кориандра, 2n = 44.
В период цветения на контрольных и полиплоидных растениях практически все цветки в соцветиях имели типичные размеры и строение без видимых морфологических изменений элементов цветка. Однако при цитологическом анализе было обнаружено, пыльники тетраплоидных цветков содержали меньше пыльцевых зёрен уменьшенного размера. Во многих пыльниках тетраплоидных цветков присутствовали как фертильные, так и стерильные пыльцевые зёрна. Кроме этого в цветках тетраплоидных зонтиков встречались сегментарно-стерильные пыльники, а также пыльники с полным отсутствием пыльцевых зёрен.
Также у диплоидных и тетраплоидных форм кориандра обнаружены различия в размерах и форме пыльцевых зерён. Обычные гаплоидные пыльцевые зёрна имели отчётливо выраженную капсуловидную форму с небольшим утолщением в средней части поперёк продольной оси пыльцевого зерна. Размеры пыльцевых зёрен в пыльниках тетраплоидов были несколько меньше по сравнению с исходной формой, причём большинство из них имели округло-угловатую, реже укороченную капсуловидную формы (рис. 3 а и б ). Предполагается, что эти явления могут быть связаны с асинхронностью митотического деления при морфогенезе тычинок и нарушениями мей-оза при микрогаметогенезе тетраплоидов.
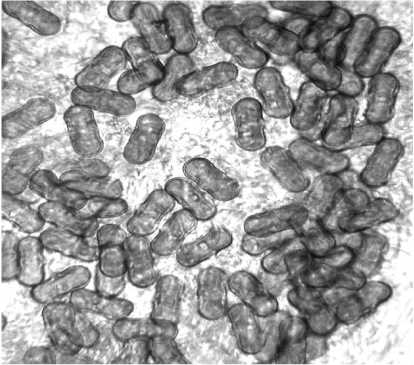
а
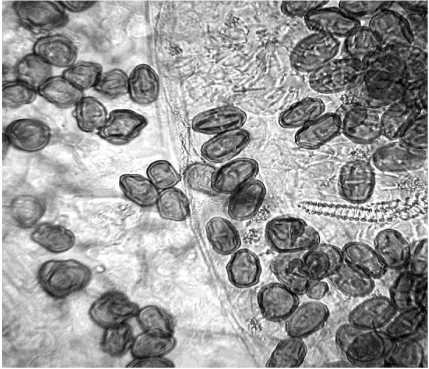
б
Рисунок 3 – Сравнительная морфология пыльцевых зёрен диплоидных и тетраплоидных (С 2 ) растений кориандра, сорт Янтарь, X 500.
а – пыльцевые зёрна диплоидного растения (видовая норма);
б – пыльцевые зёрна тетраплоидного растения.
Удвоенная плоидность растений кориандра С 2 оказала заметное влияние на формирование их репродуктивных органов. Размеры соцветий у тетраплоидов за счёт удлинения лучей зонтиков оказались крупнее, чем у контрольных растений. Как это уже отмечалось у предыдущих поколений колхицинированных форм кориандра, цветки у тетраплоидов в С 2 отличались пониженной фертильностью. В результате в тетраплоидных зонтиках за счёт повышенной абортивности цветков или прекращения развития завязей на ранних этапах формировалось меньшее количество плодов (рис. 4 а и б ).

Рисунок 4 – Сравнительная морфология зонтиков диплоидных и тетраплоидных
(С 2 ) растений кориандра, сорт Янтарь а – зонтик диплоидного растения; б – зонтик тетраплоидного растения.
Полностью сформировавшиеся и созревшие на тетраплоидных растениях плоды имели повышенную по сравнению с диплоидными формами крупность. Так например, если у сорта Янтарь оценки выборочных средних и их ошибок по длине и ширине плодов исходной диплоидной формы в среднем составляли 3,14 ± 0,16 и 2,76 ± 0,23 мм, то у тетраплоидной формы этого же сорта их размеры увеличились до 4,50 ± 0,21 и 3,74 ± 0,19 мм, или в среднем на 43 и 74 % соответственно (рис. 5а и б).

а
б
Рисунок 5 – Сравнительные размеры плодов диплоидных и тетраплоидных растений кориандра, сорт Янтарь а – плоды диплоидного растения;
б – плоды тетраплоидного растения.
Увеличенная крупность тетраплоидных плодов кориандра может быть объяснена двояко. С одной стороны, это можно отнести к известному на других культурах эффекту полиплоидного гигантизма, определяемого увеличением размеров соматических клеток. С другой стороны, увеличенная крупность плодов тетраплоидных растений может быть объяснена простым перераспределением неиспользованных пластических веществ между меньшим количеством завязавшихся плодов.
Выводы . Применение водных растворов колхицина в концентрации 0,05 % позволяет получать полиплоидные растения кориандра. Тетраплоиды кориандра отличаются увеличенными размерами листовых пластинок прикорневых листьев и увеличенной длиной черешков. Репродуктивные органы ранних (С1-С2) тетраплоидов характеризуются частичной стерильностью цветков, уменьшенным количеством пыльцы и её сниженной фертильностью. Завязываемость плодов в зонтиках тетраплоидов понижена. Сформировавшиеся плоды отличаются более крупными размерами по сравнению с диплоидными формами.
В целом в результате искусственной полиплоидизации у потомства колхицинированных растений обнаружены востребованные потребителем хозяйственно ценные признаки крупнолист-ности и крупноплодности.
После восстановления и стабилизации фертильности сохранившие крупнолистность и крупноплодность тетраплоидные либо возвратно-диплоидные формы кориандра могут быть использованы в качестве исходного материала для селекции кориандра на эти признаки.
