Морфология соцветия у представителей рода Chionodoxa (Hyacinthaceae)
")
Автор: Седельникова Л.Л.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Сельскохозяйственные науки: Агрономия
Статья в выпуске: 2, 2015 года.
Бесплатный доступ
Представлены результаты изучения соцветия у пяти видов и форм хионодокс. Определены особенности строения соцветия. Проанализирована морфологическая изменчивость соцветия в условиях интродукции.
Хионодокса, морфология, соцветие, интродукция, западная сибирь
Короткий адрес: https://sciup.org/14084127
IDR: 14084127 | УДК: 582.573.81:581.46
Morphology of floscule in Chionodoxa (Hyacinthaceae) genus representatives
The results of the floscule study in five sorts and forms of Chionodoxa are presented in the article. The peculiarities of the floscule structure are determined. The floscule morphological variability in the introduction conditions is analyzed.
Текст научной статьи Морфология соцветия у представителей рода Chionodoxa (Hyacinthaceae)
Введение. Соцветие как одна из составных частей побеговой системы выполняет специфическую роль в формировании генеративных органов покрытосеменных растений. Известно три подхода при описании соцветий – физиономическое, ритмологическое и структурное [Кузнецова, 1985; Кузнецова и др., 1992]. Последний как наиболее перспективный и современный при исследовании строения соцветий является основополагающим и первоначально отражен в работе [Troll, 1957]. Он базируется на выявлении закономерностей расположения цветоносных осей в общей осевой системе растения, независимо от степени их сформированности. Описательная морфология физиономического характера общего строения соцветий у однодольных растений в пределах семейств представлена в трудах отечественных ученых [Федоров, Кирпичников, Артюшенко, 1979], позднее в работах [Серебрякова, Воронин, Еленевский и др. 2006] даны элементы структурного анализа. В процессе переноса видов из условий естественного местообитания наблюдается морфологическая изменчивость в строении ряда признаков, в том числе и соцветия. Поэтому частные особенности строения соцветия на меж- и внутривидовом уровне еще исследованы недостаточно. К их числу относятся ранневесенние луковичные эфемероиды – представители рода Chionodoxa Boiss. семейства Hyacinthaceae Batsch . Род состоит из 5 видов, родина которых Малая Азия, о. Крит [Артюшенко, 1977]. Нами изучен жизненный цикл развития хионодокс в условиях лесостепной зоны Западной Сибири [Седельникова, 2002, 2014]. Особенности формирования их соцветия послужили основанием выполнения данной работы.
Цель работы. Исследование морфологического строения соцветия у представителей рода Chionodoxa для выявления изменчивости при интродукции в Западной Сибири.
Объекты и методы исследований . Работа выполнена в Центральном сибирском ботаническом саду СО РАН (г. Новосибирск), где интродуцировано 5 видов и форм хионодокс: Сh. luciliae Boiss. – Хионодокса Люцилии, Ch. luciliae f. coerulea hort. – Х. Люцилии форма голубая, Ch. luciliae f. rosea – Х. Люцилии – форма розовая, Ch. gigantea Witt. – Х. гигантская, Ch. gigantea f. alba hort. – Х. гигантская – форма белая. Материал получен в 1985 г. из Всесоюзного института растениеводства (г.Санкт-Петербург). Морфологическое описание генеративного побега сделано согласно общепринятой терминологии [Кузнецова и др., 1992; Серебрякова и др., 2006]. Статистическая обработка проведена с помощью компьютерной программы Statistica 6.0.
Результаты исследований и их обсуждение. Начало отрастания хионодокс в лесостепной зоне Западной Сибири по среднемноголетним данным (1986–2014 гг.) наблюдали при переходе среднесуточных температур через 0+50 С, сразу после таяния снега, с 15 по 20 апреля. Первые этапы роста и развития генеративных особей у всех видов хионодокс проходят очень быстро. После развертывания первых двух настоящих листьев, наступало цветение (III декада апреля, I–II декады мая) (табл.1).
Таблица 1
Начало цветения хионодокс в разные вегетационные периоды в условиях Новосибирска
|
Вид |
2009 г. |
2010 г. |
2011 г. |
2012 г. |
2013 г. |
2014 г. |
|
Ch. gigantea |
10.05 |
05.05 |
28.04 |
04.05 |
03.05 |
12.05 |
|
Ch. gigantea f. alba |
11.05 |
06.05 |
30.04 |
04.05 |
06.05 |
13.05 |
|
Сh. luciliae |
12.05 |
04.05 |
29.04 |
05.06 |
10.05 |
12.05 |
|
Ch. luciliae f. coerulea |
13.05 |
05.05 |
29.04 |
06.05 |
09.05 |
10.05 |
|
Ch. luciliae f. rosea |
13.05 |
06.05 |
24.04 |
06.05 |
03.05 |
10.05 |
За последние десять лет погодные условия в течение вегетационных периодов в Приобской лесостепной климатической провинции крайне противоречивы: то засушливые, то холодные с обилием осадков, поэтому адаптация интродуцентов проходила на фоне оптимума и минимума факторов среды. Так, по гидротермическим показателям тепла и запасам влаги 2007 г. был засушливым, особенно во второй половине; 2008 г. – теплым, умеренно увлажненным; 2009 г. – прохладным, избыточно увлажненным; 2010 г. – теплым, умеренно увлажненным; 2011 г. – слабозасушливым с ранней теплой весной; 2012 г. – теплым, засушливым; 2013 г. – прохладным, избыточно увлажненным; 2014 г. – холодным, избыточно увлажненным с частыми возвратными заморозками в мае, но с ранней весной. Все виды практически зацветали одновременно. Однако отмечено, что цветение у хионодокс в условиях г. Новосибирска наступает раньше на 5–16 дней в годы с ранней теплой весной (2010–2012 гг.) по сравнению с прохладной (2009 г., 2013–2014 гг.). После цветения и плодоношения (апрель-май) наступает летний (июнь-август) период относительного покоя и далее зимний (сентябрь-март).
В период цветения наблюдения проводили за одновозрастными особями с морфометрическими показателями длины листовой пластинки, генеративного побега и луковицы, представленными в таблице 2.
Таблица 2
Морфометрические показатели вегетативных и генеративных органов (min-max, см, шт.) интродуцентов рода Chionodoxa в Новосибирске
|
Вид |
1 |
2 |
3 |
4 |
5 |
6 |
|
Ch. luciliae f. rosea |
15,5-16,0 |
12,0-13,5 |
8,5-15,5 |
3-6 |
2,0-2,5 |
9-10 |
|
Ch. gigantea f. alba |
9,5-13,5 |
10,0-13,5 |
5,2-8,0 |
1-4 |
2,0-2,5 |
6-7 |
|
Сh. luciliae |
15,5-16,5 |
10,0-13,0 |
8,0-10,5 |
3-7 |
1,5-2,0 |
5-6 |
|
Ch. gigantea |
11,5-14,0 |
11,5-14,0 |
6,5-12,5 |
3-5 |
1,5-1,7 |
5-6 |
|
Ch. luciliae f. coerulea |
6,5-10,5 |
2,5 – 8,5 |
8,5-15,5 |
2-5 |
2,0-2,3 |
7-8 |
Примечание: 1 – длина первого листа, см; 2 – длина второго листа, см; 3 – длина генеративного побега, см; 4 – число цветков в соцветии, шт.; 5 – высота луковицы, см; 6 – число запасающих чешуй, шт.
В период цветения особое место уделяли особенности формирования соцветия. Его развитие рассмотрено в рамках структурного подхода морфологического описания как более современного, сочетающего в себе элементы физиономического и ритмологического. Установлено, что для всех видов хионодокс характерно соцветие простая открытая кисть. По определению [Федоров и др., 1979, с. 24], для простого соцветия характерно то, что пазушные меристемы полностью расходуются на формирование боковых цветков с цветоножками. Известно, что кисть – это соцветие с удлиненными междоузлиями главной оси и удлиненными боковыми осями (цветоножками) [Кузнецова и др., 1992, с.11]. Зачаточные цветки у хионодокс во внутрипочечном развитии формируются акропетально, и верхний (последний) цветок всегда расположен в боковом направлении к оси, по- этому верхушечная часть соцветия нарастает меристематической тканью, и ее терминальная зона открытая [Седельникова, 2014]. В соцветии всегда имеется боковой цветок, меристематическая ткань верхушечной части монокарпического побега в период цветения сильно паранхематизирует-ся, но оно открытое, что присуще для представителей семейства Hyacinthaceae и Liliaceae. Кроющие листья у всех видов видоизменены и представляют брактеи или прицветники размером 0,05– 0,1 мм с зелено-синеватым оттенком, поэтому соцветие брактеозное.
Относительно длины междоузлий главной оси соцветия установлена фенотипическая изменчивость. У Ch. gigantea , Ch. gigantea f. alba (рис., а, б, в) все междоузлия главной оси соцветия были удлиненные. Длина их главной оси составляла 7,2–12,5 см с междоузлиями 1,5–2,0 см. У Сh. luciliae , Ch. luciliae f. coerulea и Ch. luciliae f. rosea главная ось соцветия была 8,5–15,5 см и имела как удлиненные (1,5–2,0 см), так и укороченные (0,5–1,0 см) междоузлия (рис., д, ж, з). Причем чередование междоузлий в соцветии от длинных до коротких было выражено акропетально только у Сh. luciliae (рис., д). У форм Ch. luciliae f. coerulea и Ch. luciliae f. rosea наблюдали укороченные междоузлия в базальной части соцветия, средней – длинные, верхней – короткие (рис., д, ж). Расположение цветоножек с цветками очередное, с двух сторон либо с одной стороны оси соцветия. Наблюдали сильно укороченное междоузлие (0,2–0,3 см) у Ch. gigantea f. alba (рис., в).
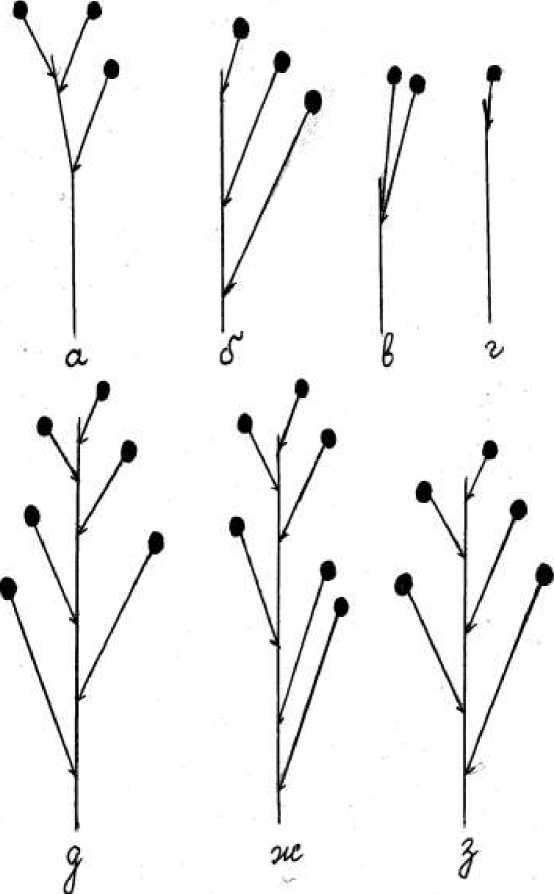
Схема соцветий у Chionodoxa gigantea (а, б), Ch. gigantea f. alba (в, г), Сh.luciliae (д), Ch. luciliae f. rosea (ж), Ch. luciliae f. coerulea (з)
Относительно длины боковых осей (цветоножек) установлена изменчивость этого признака от длинных до коротких. Длинные цветоножки (от 2,5 до 4,1 см) наблюдали у цветков от первого до третьего порядка. Верхние цветки имели короткую цветоножку – от 0,5 до 0,8 см. Чередование длинных и коротких цветоножек отмечено у Сh. luciliae , Ch. luciliae f. rosea (рис., д, ж). Таким образом, длина цветоножек варьировала в пределах соцветия, но физиономически она всегда уменьшалась снизу вверх от 4,0 до 0,8 см.
Следует отметить, что у однодольных растений, особенно у близкородственных таксонов ( Scilla L., Tulipa L., Crocus L.), очень часто наблюдаются редукционные явления в открытой кисти, в которой формируется только один цветок. Такая же редукция обнаружена нами у Ch. gigantea f. alba , где одновременно с формированием в кисти по два-три цветка у отдельных особей формируется только один цветок (рис., г) независимо от их возрастного состояния. Такие соцветия рассматриваются как открытые. Поэтому одиночный цветок у Ch. gigantea f. alba – это единственно сохранившейся элемент флоральной единицы (ФЕ) – открытой кисти. Всего цветков в соцветии у данных интродуцентов от 1 до 7 шт. (см. табл.2).
Относительно универсальной схемы строения цветоносного побега у хионодокс за флораль-ную единицу (ФЕ) следует принимать годичный побег, т.е. открытое рацемозное соцветие без пара-кладиев. Понятие синфлоресценции, по В.Троллю (1957), включает совокупность цветоносных осей годичного побега с главной верхушечной ФЕ и все паракладии. Поэтому в нашем случае соцветие у хионодокс в физиономическом смысле оказывается отдельным элементом синфлоресценции, а именно ФЕ. Установленные нами структурные типы соцветия у хионодокс не обладают строгой дискретностью и связаны между собой переходными формами. Однако каждая ось соцветия формируется за счет деятельности одной апикальной меристемы, является побегом одного порядка. По классификации структурных типов, соцветия представителей рода Chionodoxa отнесены к монопо-диальным, рацемозным, с акропетальным порядком формирования и распускания цветков.
Таким образом, у ранневесенних эфемероидов из рода Chionodoxa формирование соцветия и их фенотипическая изменчивость обусловлены высокоспециализированной адаптацией видов к внешним факторам в процессе исторического становления жизненной формы, связанной с укороченным ритмом надземного развития и длительным периодом внутрипочечного подземного развития в период относительного летнего и зимнего покоя.
Выводы
-
1. У представителей рода Chionodoxa ( Сh. luciliae Boiss. – Хионодокса Люцилии, Ch. luciliae f. coerulea hort. – Х. Люцилии – форма голубая, Ch. luciliae f. rosea – Х. Люцилии – форма розовая, Ch. gigantea Witt. – Х. гигантская, Ch. gigantea f. alba hort. – Х. гигантская – форма белая) соцветие кисть – простое, брактеозное, с моноподиальным нарастанием главной оси рацемозного типа.
-
2. В условиях интродукции отмечена редукция открытой кисти, выраженная в формировании одиночного цветка.
-
3. Выделена фенотипическая изменчивость морфологических признаков с удлиненными и укороченными междоузлиями главной оси соцветия, с акропетальным развитием цветка и длиной цветоножки от 0,5 до 4,1 см.
-
4. Установлено межвидовое и внутривидовое морфологическое разнообразие в строении простой кисти как годичного побега без паракладиев, идентичного элементу синфлоресценции.
