Мутация HCD у российских голштинизированных черно-пестрых коров не влияет на молочную продуктивность и содержание холестерина и триглцеридов в крови

Автор: Позовникова М.В., Лихачева Т.Е., Кудинов А.А., Лейбова В.Б., Дементьева Н.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика и селекция
Статья в выпуске: 6 т.53, 2018 года.
Бесплатный доступ
Распространение летальных и полулетальных генетических мутаций в популяциях крупного рогатого скота становится причиной эмбриональной и постэмбриональной смертности телят. Использование ограниченного числа быков-производителей создает опасность широкого распространения генетических аномалий. Рецессивный дефект голштинского скота - дефицит холестерина HCD (haplotype cholesterol deficiency) характеризуется гибелью телят в первые дни или месяцы жизни. Степень распространения этого дефекта в настоящее время очень высока (в разных странах от 6 до 17 %). Следует отметить, что в целом сведений о связи рецессивных мутаций с продуктивностью молочного скота немного, а данные о влиянии мутации HCD, впервые описанный в 2015 году, на селекционно значимые признаки крайне ограничены. Мы впервые провели генетическую оценку района гена APOB на хромосоме BTA11 в российской популяции коров и определили показатели молочной продуктивности в зависимости от статуса по HCD. Полученные результаты свидетельствуют, что в изученной нами популяции мутация HCD не снижает племенную ценность животных по молочной продуктивности и качеству молока (по жиру и белку)...
Крупный рогатый скот, генотипирование, гаплотип дефицита холестерина, летальная рецессивная мутация, аполипопротеин b, ген apob, молочная продуктивность, триглицериды, холестерин
Короткий адрес: https://sciup.org/142220062
IDR: 142220062 | УДК: 636.22/.28:575.174.015.3 | DOI: 10.15389/agrobiology.2018.6.1142rus
Cholesterol deficiency mutation HCD does not impact milk productivity and blood levels of cholesterol and triglycerides in Russian Holstein black and white cattle
The spread of lethal and semi-lethal mutations in cattle populations results in embryonic and postembryonic mortality of calves. The use of a limited number of sires creates the danger of wide spread of genetic abnormalities. Genetic markers identify carriers of a mutation in the absence of information about phenotypic manifestations of the disease. Cholesterol deficiency mutation (HCD, haplotype cholesterol deficiency), a recessive defect of Holstein cattle, is characterized by the death of calves in the first days or months of life. The extent of this genetic defect worldwide is currently very high, 6 to 17 %. In general, there is little information about the relationship of recessive mutations with dairy cattle productivity, and data on the effect of the HCD mutation, first described in 2015, on breeding traits are extremely limited. This paper is the first to report data on a genetic study of the APOB gene region on the BTA11 chromosome and milk production indices depending on the HCD status in a Russian dairy cow population...
Текст научной статьи Мутация HCD у российских голштинизированных черно-пестрых коров не влияет на молочную продуктивность и содержание холестерина и триглцеридов в крови
* Работа выполнена при финансовой поддержке ФАНО России (тема ГЗ ¹ 0600-2018-0014).
Искусственное осеменение и использование ограниченного числа производителей создает опасность распространения летальных рецессивных мутаций среди крупного рогатого скота (1). Далеко не все аномалии поддаются визуальному наблюдению. ДНК-скрининг с использованием SNP-чипов высокой плотности позволяет выявлять мутации без наличия сведений о фенотипических проявлениях заболевания (2). Этим методом определяют гаплотипы фертильности, встречающиеся у крупного рогатого скота и становящиеся причиной смертности на разных этапах развития животного. Скрининг отечественного голштинского и черно-пестрого голштинизированного скота показал, что частота встречаемости известных мутаций CVM, BLAD, DUMPS, BY, HCD, HH1, HH3, HH4, HH5 достигает 10 % у коров и 4 % у быков-производителей (3). Регулярный мониторинг распространения вредных рецессивных мутаций среди поголовья необходим и позволяет вовремя исключить из разведения их носителей, значительно сократив экономические потери хозяйств. Так, в Ленинградской области регулярная проверка голштинских быков и своевременная выбраковка носителей BLAD и CVM позволила снизить частоту встречаемости этих мутаций в отдельных хозяйствах до 1-2 % (4-6).
Гаплотип дефицита холестерина (HCD, haplotype cholesterol deficiency) — новый рецессивный генетический дефект голштинского скота. Об идентификации этого гаплотипа, ассоциированного с гибелью телят в ранний постнатальный период вследствие возникновения идиопатической диареи, не поддающейся терапии, впервые сообщили S. Kipp c соавторами в 2015 году на конференции Interbull в Орландо (США). Гомозиготные животные имели выраженное нарушение жирового обмена и гипохолесте-ринемию. У гетерозиготных телят при отсутствии клинических признаков отмечали низкие показатели холестерина в крови. В результате поиска геномных ассоциаций с помощью сканирования 44747SNP (чип Illumina Bo-vineSNP50 BeadChip, версия 2; 54Kv2; «Illumina, Inc.», США) у пораженных телят обнаружили гомозиготный район размером 1,01 Mb на BТА11 (c позиции 77274120 до 78290130 bp), что указывает на аутосомное моноген-ное наследование этого расстройства, действующего по рецессивному или кодоминантному типу. Значимый SNP расположен в позиции 72248536 bp на расстоянии около 5 Mb от дефектного гаплотипа (7). Поиск геномных мутаций с помощью изучения полногеномных ассоциаций GWAS (genomewide association studies) позволил выявить в общей сложности 22 SNPs (с 64367438 до 83585365 bp), которые достигли порога достоверности около дефектного гаплотипа. Изучение генома на основе генотипов 54K SNP Chip определило казуальный участок на хромосоме BTA11. Анализ родословных больных животных выявил выдающегося канадского голштинского быка Maughlin Storm как носителя такого расстройства (8, 9). Дальнейшие исследования показали, что причиной этой мутации служит вставка длинной 1299 bp в 5-м экзоне гена APOB (аполипопротеин B) на ВТА11, приводящая к сдвигу рамки считывания в области кодона для 135-го аминокислотного остатка в белке APOB. В результате происходит усечение 97 % соответствующего белка (10).
Другие авторы обнаружили на BTA11 вставку усеченного эндогенного ретровируса типа ERV2-1 в LTR (Long Terminal Repeats) в 5-м экзоне гена APOB, что привело к образованию стоп-кодона недалеко от инсер-ции. Генерация преждевременного стоп-кодона в открытой рамке считывания гена APOB вызывает усечение длины белка до 140 аминокислот. Установлено, что такие ранние укорочения вызывают неспособность выведения хиломикронов из клеток кишечника, что приводит к мальабсорб- ции холестерина (11). Белок аполипопротеин B (APOB) необходим для синтеза хиломикронов и липопротеинов очень низкой плотности в кишечнике и печени. Аполипопротеины представляют собой белковые, обычно амфифильные, компоненты липопротеинов, специфически связывающиеся с соответствующими липидами при формировании липопротеиновой частицы (12, 13).
Анализ данных о частоте встречаемости HCD в стадах голштинского скота в разных странах выявляет высокий процент носительства: в Китае (быки, n =138) — 5,07 % (14), в Германии (быки, n = 264) — 17,4 % (10), в Канаде (телки 2012 и 2016 года рождения) — соответственно 17 и 12 % (15). По результатам анализа родословных 584 быков-производителей, используемых для разведения в России, у 10,3 % (60 быков) отцы оказались скрытыми носителями мутантного аллеля гена APOB (16) . При этом происхождение отцов различалось (Канада, Америка, Австрия). По результатам генотипирования 41 быка, отцы которых были носителями HCD, 17 животных (39 %) оказались скрытыми носителями этой мутации. Своевременный скрининг популяций на наличие генетических дефектов крупного рогатого скота и правильный подбор животных снизят экономические затраты хозяйств. По данным J.B. Cole с соавт. (17), в США экономические потери от эмбриональной и постэмбриональной смертности вследствие летальных мутаций составляют порядка 11 млн долларов в год.
Мы впервые провели генетическую оценку района гена APOB и сравнили молочную продуктивность коров в одной из российских популяций в зависимости от статуса по HCD. Полученные результаты показали, что мутация HCD не снижает племенную ценность животных по удою и качеству молока по жиру и белку.
Наша цель заключалась в оценке распространенности мутации HCD в выборке российских голштинизированных черно-пестрых коров и их сравнении по признакам молочной продуктивности и показателям липидного обмена.
Методика. Анализируемая выборка ( n = 451) голштинизированных черно-пестрых коров ( Bos taurus taurus ) одного из племенных хозяйств Ленинградской области (2017 год) была случайной, в нее вошли животные с 2009 по 2015 год рождения. Выборка телят ( n = 7) включала особей с клиническими признаками диареи, имеющих в родословной (отцы, отцы отцов) подтвержденных носителей HCD.
ДНК выделяли фенольным методом (18) из крови, полученной из хвостовой вены.
Для генотипирования методом ПЦР использовали праймеры: прямой общий — 5´-GGTGACCATCCTCTCTCTGC-3´; обратный для определения дикого аллеля — 5´-AGTGGAACCCAGCTCCATTA-3´ (обеспечивает амплификацию фрагмента размером 249 bp); прямой для определения мутантного аллеля гена APOB (indel-полиморфизм) — 5´-CACCTTCC-GCTATTCGAGAG-3´ (обеспечивает амплификацию фрагмента размером 436 bp) (19). ПЦР проводили по следующей схеме: 1 мин при 95 °С (первоначальная денатурация); 30 с при 94 °С, 30 с при 60 °С, 30 с при 72 °С (35 циклов); 10 мин при 72 °C (амплификатор Thermal Cycler T1000, «Bio-Rad Laboratories, Inc.», США). В состав реакционной смеси входили 67 мМ Трис-HCl (pH 8,6), 1,5 мМ MgCl 2 , 16,6 мМ NHOH, 0,125 мМ каждого из дезоксирибонуклеозидтрифосфатов (dATP, dGTP, dCTP, dTTP), 0,5 мкМ каждого праймера, 50-100 нг геномной ДНК и 2,5 ед. Taq ДНК- полимеразы (ООО «Сибэнзим», Россия). Полученные фрагменты ДНК разделяли методом горизонтального электрофореза при 10 В/см в 1½ ТВЕ бу-1144
фере с бромистым этидием (0,1 мкг/мл) в 2,0 % агарозном геле Agarosa LE 2 («Helicon», Россия). Размеры ампликонов определяли относительно маркера молекулярных масс ThermoScientific Gene Ruler Ultra Low Range DNA Ladder («Fermentas», Литва). Документирование и обработку результатов осуществляли с помощью видеосистемы Gel Imager 2 («Helicon», Россия).
Неравновесие по сцеплению (LD) анализировали на выборке дочерей быков-носителей HCD (Ленинградская обл.). Генотипирование с помощью чипа BovineSNP50 BeadChip v. 3 («Illumina, Inc.», США) проводили согласно рекомендациям производителя. Исследования генотипов ограничили районом гена APOB на хромосоме ВТА11, включающим 33 SNPs, длиной около 2000 kbр. Расчеты LD (R2) проводили в программе PLINK 1.9 (20).
Содержание холестерина и триглицеридов в крови определяли у животных-носителей (HCD+, n = 3) и условно здоровых животных 2-4-й лактации (HCD - , n = 14), находящихся в первой половине сухостойного периода. Кровь для биохимических исследований отбирали из хвостовой вены с помощью вакуумной системы Vacuette («Greiner Bio-One», Австрия) через 2 ч после раздачи корма (с 10.00 до 11.00). Через 30-40 мин пробирки с кровью центрифугировали 20 мин при 3000 об/мин. Концентрацию триглицеридов и холестерина измеряли на автоматическом биохимическом анализаторе RX Daytona («Randox Laboratories», Великобритания) с использованием реагентов фирмы «Сormay» (Польша).
Данные по молочной продуктивности коров 2012-2014 годов рождения были взяты из племенных карточек (форма 2МОЛ). Учитывали удой по 1-й и 2-й лактации, выход молочного жира и выход молочного белка.
Связь между генотипами коров и анализируемыми признаками выявляли на основе оценки достоверности различий между средними значениями. Найденную величину td сравнивали с таблицей Стьюдента (21). Дисперсионный анализ ANOVA и подсчет средних проводили в программе RStudio (22) на основании модели с одним фиксированным эффектом, имеющей вид: yij = µ + HCDj + eij, где yij — расчетная племенная ценность (РПЦ) коровы i по признакам продуктивности, µ — среднее, HCDj — фиксированный эффект гаплотипа, eij — остаточное неизвестное. Статистическую обработку данных выполняли в программах Microsoft Excel и AtteS- tat . Представлены средние значения (М) и стандартные ошибки средних (±SEM).
Расчетную племенную ценность (РПЦ) по признакам удой, жир и белок вычисляли методом BLUP Animal Model по данным фенотипического учета 2016 года (23). Эффект гаплотипа представляли в виде кодировки 0 (HCD - ) и 1 (носитель HCD+).
Результаты. На рисунке 1 представлена типичная электрофореграмма продуктов ПЦР-амплификации на наличие indel полиморфизма в гене APOB .
Всего мы протестировали 55,7 % маточного поголовья коров с подозрением на носительство мутантного аллеля HCD гена АРОВ. По результатам исследования 35 коров (7,76 %) и одна телочка ока-
М 1 2 3 4 5
700 500 400 300 200 150 100
Рис. 1. Электрофореграмма продуктов ПЦР-амплифи-кации на наличие indel полиморфизма в гене APOB у голштинизированных черно-пестрых коров: М — маркер молекулярных масс; 1, 3 — здоровые животные (присутствует фрагмент длиной 249 bp), 2, 4 — гетерозиготные носители (фрагменты 249 и 436 bp), 5 — гомозигота (фрагмент 436 bp) (Ленинградская обл., 2017 год).

Рис. 2. Телка голштинизированной черно-пестрой породы, гомозиготная по мутантному аллелю гена АРОВ , с типичными клиническими проявлениями синдрома дефицита холестерина (отставание в росте и развитии, истощение, диарея) (Ленинградская обл., 2017 год).
предков матери был бык Maughlin
зались гетерозиготными носителями мутации HCD. Только у одной телочки определили гомозиготный генотип по HCD гена АРОВ . У этого животного отмечали резкое отставание в росте и развитии, истощение, диарея, не поддающаяся медикаментозному лечению (рис. 2). Масса при рождении составила 39 кг, в 2 мес — 49 кг, в 3 мес — 49 кг; среднесуточный прирост массы от рождения до 2-месячного возраста — 166 г. Согласно результатам генотипирования, статус матери по HCD был положительным, при этом отец также нес мутантный аллель . Анализ родословных родителей показал, что среди Storm 5457798 среди предков отца —
Breadale Goldwyn 10705608. Эти быки и их потомки используются в системе искусственного осеменения в России уже несколько лет и отмечены как скрытые носители HCD.
1. Показатели молочной продуктивности у коров голштинизированной ч е рнопестрой породы по вариантам генотипов с indel полиморфизмом в гене APOB (мутация дефицита холестерина HCD) ( M± SEM, Ленинградская обл., 2017 год)
|
Год рождения Статус по HCD |
Число животных, гол. |
Удой, кг |
Жир, кг |
Белок, кг |
|
2012 HCD+ |
1-я лактация 3 |
8750±440 |
342,6±28,6 |
281,7±20,4 |
|
HCD - |
22 |
8894±256 |
333,7±10,1 |
280,8±7,8 |
|
2013 HCD+ |
8 |
9471±261a |
353,8±7,8 |
292,0±8,5c |
|
HCD - |
73 |
8252±142b |
340,0±5,9 |
272,1±4,5d |
|
2014 HCD+ |
10 |
8780±378 |
346,5±10,0 |
268,9±9,1 |
|
HCD - |
101 |
8646±126 |
347,0±5,4 |
271,0±4,1 |
|
2012 HCD+ |
2-я лактация 3 |
10339±787 |
390,6±76,9 |
330,8±43,8 |
|
HCD - |
22 |
9596±385 |
371,9±16,4 |
300,1±11,3 |
|
2013 HCD+ |
6 |
10872±346е |
428,7±20,1g |
339,7±12,2i |
|
HCD - |
49 |
9480±233f |
384,7±9,0h |
299,9±6,9j |
П р и м еч а ни е. Буквами отмечены статистически значимые различия между группами HCD+ и HCD-: a, b — при p < 0,01; c, d — при p < 0,05; e, f — при p < 0,001; g, h — при p < 0,05; i, j — при p < 0,01.
2. Дисперсионный анализ расчетной пле- Коровы HCD не усту- менной ценности голштинизированных пали сверстницам, а иногда черно-пестрых коров, гетерозиготных по даже превосходили их по пока-мутации дефицита холестерина HCD (Ле- зателям молочной продуктив-нинградская обл., 2017 год) ности (табл. 1). Так, скрытые
Признак продуктивности ] Эффект HCD p-value носители мутантного аллеля Удой 4,076 0,0442 гена APOB 2013 года рождения
Жир 6,617 0,0105
Белок 1,905 0,1680 достоверно превосходили здо ровых животных: в 1-ю лактацию по удою на 1219 кг (p < 0,01), по выходу молочного жира — на 13,8 кг, по выходу молочного белка — на 19,9 кг (p < 0,05); во 2-ю лактацию — соответственно на 1392 кг (p < 0,001), 44 кг (p < 0,05) и 39,8 кг (p < 0,01). Между группами коров 2012 и 2014 года рождения не было выявлено достоверных различий.
Средняя РПЦ по признакам удой, жир и белок для носителей
HCD составила соответственно 1100,8; 42,5 и 28,3 кг, а для животных
HCD- — 1030,5; 39,3 и 27,1 кг. То есть у первых она была выше по удою на 6,8 %, по жиру — на 8,1 %, по белку — на 4,8 %. Дисперсионный ана- лиз (табл. 2) показал стойкий положительный эффект влияния носительства по HCD на продуктивность.
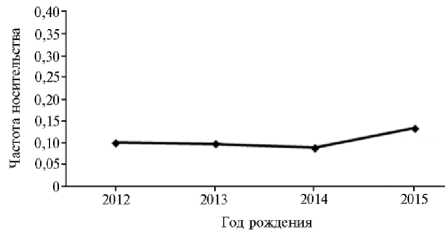
Рис. 3. Частота носительства мутации дефицита холестерина HCD среди голштинизированных черно-пестрых коров разных лет рождения (Ленинградская обл., 2017 год).
Среди протестированных животных разных лет рождения частота HCD составляла около 10 % (рис. 3) при тенденции к повышению в потомстве, рожденном в 2015 году, что указывает на необходимость постоянно контролировать поголовье в племенных хозяйствах для сокращения числа носителей мутации HCD.
Мы не выявили достоверных различий между группами носителей HCD+ и ко- ров HCD- по содержанию в сыворотке крови холестерина (соответственно 3,04±0,31 и 3,33±0,12 ммоль/л) и триглицеридов (соответственно 0,197±0,01 и 0,170±0,01 ммоль/л). Для обеих групп концентрация этих метаболитов была в пределах референсных значений, которые составляют 1,5-4,5 ммоль/л для холестерина и 0,05-0,3 ммоль/л для триглицеридов (24). Молочная продуктивность за 305 сут последней завершенной лактации по группам не различалась и составляла 10302±791 и 10191±453 кг.
Для более полной оценки влияния отбора по молочной продуктивности на распространение носителей гаплотипа холестеринового дефицита HCD мы проанализировали неравновесие по сцеплению (LD) между SNP в районе гена APOB на расстоянии около 1000 bp от гена в обе стороны. Среднее расстояние между SNPs на чипе в изучаемой области составило около 55 kbp. Расчеты показали отсутствие гаплоблоков на этом участке нуклеотидной последовательности в геноме у потомков быков — носителей HCD в популяции голштинского скота в Ленинградской области. Средний показатель LD был низким (R2 = 0,077±0,008).
Отметим, что, согласно родословным, в хозяйстве, где проводились исследования, для разведения использовались голштинские быки разного происхождения (Нидерланды, США, Канада, Россия). Некоторые из них имели подтвержденный статус скрытых носителей HCD. В литературе содержится недостаточно информации о связи рецессивных мутаций с продуктивностью молочного скота. Однако, как показывает практика, быки — носители рецессивных мутаций зачастую служат улучшателями, а широкому распространению генетических дефектов способствует то, что они, как правило, сцеплены с генами хозяйственно полезных признаков (25). Так, анализ родословных сыновей и внуков Skokie Sensation Ned, родоначальника расстройства DUMPS, показал, что гетерозиготы имеют значительно более высокий генетический потенциал молочной продуктивности (26). S. Saleem с соавт. (27) отмечают, что качественные показатели спермы голштинских быков не изменяются в зависимости от статуса по HCD. В предыдущих исследованиях мы выяснили, что статус по HCD не оказывает существенного влияния на такие репродуктивные показатели коров, как возраст первого осеменения, первого отела, число осеменений до плодотворного, длительность сервис-периода и межотельного периода (28).
В ряде работ (29, 30) установлено, что быки и телята — скрытые носители мутации HCD имеют более низкое содержание триглицеридов и холестерина в крови по сравнению с животными, у которых отсутствует мутантный аллель гена APOB . Однако на содержание холестерина в крови (как и на концентрацию триглицеридов) влияют такие факторы, как питание, физиологическое состояние животного и заболевания различной этиологии. Поэтому диагноз на наличие дефекта HCD можно поставить только с помощью молекулярно-генетического тестирования. В своем исследовании мы не выявили достоверных отклонений от нормы в биохимических показателях сыворотки крови у носителей этого генетического дефекта, что может быть связано как с малой выборкой, так и с наличием у некоторых животных генетически детерминированных компенсаторных механизмов (11, 31).
Отслеживание селекционно-генетических характеристик в породе и определение генеалогической принадлежности быка необходимы для контроля генетических дефектов (32, 33).
Интересно отметить, что носители мутации HCD не снижают продуктивность в стаде. Более того, в изученной популяции средняя расчетная племенная ценность по признакам удой, жир и белок у носителей HCD оказалась выше, чем у интактных животных. Дисперсионный анализ выявил стойкий положительный эффект влияния носительства HCD на продуктивность. Такое превосходство не имеет четкого объяснения, но может быть обусловлено нахождением гена APOB в участке генома, отвечающем за высокую молочную продуктивность. В то же время анализ неравновесия по сцеплению между SNP в этом районе показал отсутствие гаплоблоков в популяции голштинского скота из Ленинградской области. Следовательно, селекция на повышение продуктивности не приведет к значительному росту частоты носителей мутации HCD.
Таким образом, мы не выявили различий по концентрации холестерина и триглицеридов в сыворотке крови в первую фазу сухостойного периода в изученной популяции коров голштинизированной черно-пестрой породы в зависимости от статуса по рецессивной мутации HCD (гаплотип дефицита холестрина). Использование коров — носителей мутации HCD не снижает продуктивность в стаде. Тем не менее мониторинг популяций на носительство указанного генетического дефекта необходим, поскольку использование гетерозиготных быков может приводить к рождению больного и нежизнеспособного потомства и, как следствие, — к экономическим потерям в хозяйствах.
Список литературы Мутация HCD у российских голштинизированных черно-пестрых коров не влияет на молочную продуктивность и содержание холестерина и триглцеридов в крови
- Атлас генетических болезней и признаков продуктивности крупного рогатого скота/Под ред. А.А. Кудинова, К.В. Племяшова, П.И. Уколова, Г.В. Ширяева. СПб, 2017.
- Charlier C., Coppieters W., Rollin F., Desmecht D., Agerholm J., Carta E., Dardano S., Dive M., Fasquelle C., Frennet J.-C., Hanset R., Hubin X., Jorgensen C., Karim L., Kent M., Harvey K., Pearce B.R., Simon P., Tama N., Nie H., Vandeputte S., Lien S., Longeri M., Fredholm M., Harvey R.J., Georges M. Highly effective SNP-based association mapping and management of recessive defects in livestock. Nat. Genet., 2008, 40(4): 449 ( ) DOI: 10.1038/ng.96
- Зиновьева Н.А. Гаплотипы фертильности голштинского скота. Сельскохозяйственная биология, 2016, 51(4): 423-435 ( ) DOI: 10.15389/agrobiology.2016.4.423rus
- Дементьева Н.В., Митрофанова О.В., Кудинов А.А. Анализ частоты встречаемости трех рецессивных летальных мутаций у коров. Известия Санкт-Петербургского государственного аграрного университета, 2015, 39: 136-143.
- Дементьева Н.В., Митрофанова О.В., Тыщенко В.И., Никиткина Е.В., Терлецкий В.П., Яковлев А.Ф. Встречаемость и значение мутации CVM у племенных животных Ленинградской области. Молочное и мясное скотоводство, 2014, 6: 7-9.
- Яковлев А.Ф., Терлецкий В.П., Митрофанова О.В., Дементьева Н.В. Определение носителей генетических дефектов среди быков-производителей. Молочное и мясное скотоводство, 2004, 6: 31-32.
- Kipp S., Segelke D., Schierenbeck S., Reinhardt F., Reents R., Wurmser C., Pausch Н., Fries R., Thaller G., Tetens J., Pott J., Haas D., Raddatz B.B., Hewicker-Trautwein M., Proios I., Schmicke M., Grünberg W. Identification of a haplotype associated with cholesterol deficiency and increased juvenile mortality in Holstein cattle. J. Dairy Sci., 2016, 99(11): 8915-8931 ( )
- DOI: 10.3168/jds.2016-11118
- Jung S., Pausch H., Langenmayer M.C., Schwarzenbacher H., Majzoub-Altweck M., Gol-lnick N.S., Fries R. A nonsense mutation in PLD4 is associated with a zinc deficiency-like syndrome in Fleckvieh cattle. BMC Genomics, 2014, 15: 623 ( )
- DOI: 10.1186/1471-2164-15-623
- Kipp S., Segelke D., Schierenbeck S., Reinhardt F., Reents R., Wurmser C., Pausch H., Fries R., Thaller G., Tetens J., Pott J., Piechotta M., Grünberg W. A new Holstein haplotype affecting calf survival. Interbull Bulletin, 2015, 49: 49-53.
- Menzi F., Besuchet-Schmutz N., Fragnière M., Hofstetter S., Jagannathan V., Mock T., Raemy A., Studer E., Mehinagic K., Regenscheit N., Meylan M., Schmitz-Hsu F., Drögemüller C. A transposable element insertion in APOB causes cholesterol deficiency in Holstein cattle. Anim. Genet., 2016, 47(2): 253-257 ( )
- DOI: 10.1111/age.12410
- Schütz E., Wehrhahn C., Wanjek M., Bortfeld R., Wemheuer W.E., Beck J., Brenig B. The Holstein Friesian lethal haplotype 5 (HH5) results from a complete deletion of TBF1M and cholesterol deficiency (CDH) from an ERV-(LTR) insertion into the coding region of APOB. PLoS ONE, 2016, 11(4): e0154602 ( )
- DOI: 10.1371/journal.pone.0157618
- Hebbachi A.M., Gibbons G.F. Microsomal membrane-associated apoB is the direct precursor of secreted VLDL in primary cultures of rat hepatocytes. J. Lipid Res., 2001, 42(10): 1609-1617.
- Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека. М., 1993.
- Li Y., Fang L., Liu L., Zhang S., Ma Z., Sun D. The cholesterol deficiency-associated mutation in APOB segregates at low frequency in Chinese Holstein cattle. Canadian Journal of Animal Science (в печати, published on the web 16 October 2018) ( )
- DOI: 10.1139/CJAS-2017-0108
- Van Doormaal B., Beavers L. HCD: haplotype associated with cholesterol deficiency. Canadian Dairy Network (CDN), 2015. Режим доступа: https://www.cdn.ca/images/uploaded/fi-le/HCD%20Update%20Article%20-%20December%202015.pdf. Дата обращения: 31.01.2018.
- Зиновьева Н.А., Костюнина О.В., Волкова В.В., Ермилов А.Н., Янчуков И.Н. Дефицит холестерина -новый рецессивный дефект голштинского скота. Молочное и мясное скотоводство, 2016, 2: 5-8.
- Cole J.B., Null D.J., VanRaden P.M. Phenotypic and genetic effects of recessive haplotypes on yield, longevity and fertility. J. Dairy Sci., 2016, 99: 7274-7288 ( )
- DOI: 10.3168/jds.2015-10777
- Терлецкий В.П., Племяшов К.В., Тыщенко В.И., Дементьева Н.В. Использование современных молекулярно-генетических методов в генотипировании сельскохозяйственных животных. СПб-Пушкин, 2014.
- Kamiński S., Ruść A. Cholesterol deficiency -new genetic defect transmitted to Polish Holstein-Friesian cattle. Pol. J. Vet. Sci., 2016, 19(4): 885-887 ( )
- DOI: 10.1515/pjvs-2016-0110
- Chang C.C., Chow C.C., Tellier L.C., Vattikuti S.S., Purcell S.M., Lee J.J. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience, 2015, 4: 7 ( )
- DOI: 10.1186/s13742-015-0047-8
- Меркурьева Е.К. Биометрия в животноводстве. М., 1977.
- RStudio Team. RStudio: integrated development for R. RStudio, Inc., Boston MA, 2015. Режим доступа: http://www.rstudio.com. Без даты.
- Kudinov A.A., Juga J., Uimari P., Mantysaari E.A., Stranden I., Plemyashov K.V., Saksa E.I., Smaragdov M.G. Upgrading dairy cattle evaluation system in Russian Federation. Interbull Bulletin, 2017, 51: 67-74.
- Кондрахин И.П., Архипов А.В., Левченко В.И., Таланов Г.А., Фролова Л.А., Новиков В.Э. Методы ветеринарной клинической лабораторной диагностики. М., 2004.
- Зиновьева Н.А., Стрекозов Н.И., Ескин Г.В., Турбина И.С., Янчуков И.Н., Ермилов А.Н. Моногенные наследственные дефекты и их роль в воспроизводстве. Животноводство России, 2015, 6: 30.
- Shanks R.D., Greiner M.M. Relationship between genetic merit of Holstein bulls and deficiency of uridine-5´-monophosphate synthase. J. Dairy Sci., 1992, 75(7): 2023-2029 ( )
- DOI: 10.3168/jds.S0022-0302(92)77962-4
- Saleem S., Heuer C., Sun C., Kendall D., Moreno J., Vishwanath R. The role of circulating low-density lipoprotein levels as a phenotypic marker for Holstein cholesterol deficiency in dairy cattle. J. Dairy Sci., 2016, 99(7): 5545-5550 ( )
- DOI: 10.3168/jds.2015-10805
- Позовникова М.В., Лихачева Т.Е., Ширяев Г.В. Влияние мутации дефицит холестерина на репродуктивные качества коров голштинизированной черно-пестрой породы. Генетика и разведение животных, 2018, 2: 61-66.
- Gross J.J., Schwinn A.C., Schmitz-Hsu F., Menzi F., Drögemüller C., Albrecht C., Bruckmaier R.M. Rapid Communication: Cholesterol deficiency-associated APOB mutation impacts lipid metabolism in Holstein calves and breeding bulls. J. Аnim. Sci., 2016, 94(4): 1761-1766 ( )
- DOI: 10.2527/jas.2016-0439
- Mock T., Mehinagic K., Menzi F., Studer E., Oevermann A., Stoffel M. H., Drögemüller С., Meylan M., Regenscheit N. Clinicopathological phenotype of autosomal recessive cholesterol deficiency in Holstein cattle. J. Vet. Intern. Med., 2016, 30(4): 1369-1375 ( )
- DOI: 10.1111/jvim.13976
- Inokuma H., Horiuchi N., Watanabe K.I., Kobayashi Y. Retrospective study of clinical and laboratory findings of autosomal recessive cholesterol deficiency in Holstein calves in Japan. Jpn. J. Vet. Res., 2017, 65(2): 107-112 ( )
- DOI: 10.14943/jjvr.65.2.107
- Племяшов К.В., Сакса Е.И., Барсукова О.Е. Селекция голштинского скота при чистопородном разведении. Генетика и разведение животных, 2016, 1: 8-16.
- Сакса Е.И., Барсукова О.Е. Результаты использования и генеалогические схемы быков-производителей голштинской породы. СПб, 2012.
