Накопление засоляющих ионов и рост подсолнечника в условиях ограниченной освещенности

Автор: Лысенко В.С., Ефименко С.Г.
Статья в выпуске: 1 (136), 2007 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/142150789
IDR: 142150789
Текст статьи Накопление засоляющих ионов и рост подсолнечника в условиях ограниченной освещенности
Подсолнечник Helianthus annus L. относят к растениям с умеренной солеустойчивостью аккумуляторного (соленакапливающего) типа. В последнее время к этой культуре все более возрастает интерес с точки зрения эффективного использования засоленных земель. Кроме того, подсолнечник может применяться с целью рекультивации пахотного слоя. Так, например, Бхаттом и Индиракутти [1] было показано, что выращивание подсолнечника позволяет снизить засоление почвы ионами Na+ и Cl–.
Для получения новых солеустойчивых сортов и гибридов подсолнечника исключительно важно понимать механизмы и типы формирования устойчивости. Различают специфическую и неспецифическую солеустойчи-вость. Неспецифическая солеус-тойчивость развивается вне зависимости от характера раздражения и ассоциируется преимущественно с процессами, модулирующими важнейшие звенья энергетического метаболизма, включая гормональный баланс, фотосинтез и дыхание [2]. Особый интерес здесь вызывает регуляция процесса фотосинтеза.
Несмотря на очевидную в ряде случаев необходимость увеличения энергозатрат, связанную, например, с работой Na+/H+ – антипортеров (у галофитов соленакапливающего типа) или синтезом осмолитов (у галофитов исключающих соль) [3] при воздействии даже умеренного солевого стресса интенсивность фотосинтеза обычно снижается [4, 5].
В этой связи вполне логична распространенная точка зрения, согласно которой снижение интенсивности фотосинтеза, наблюдаемое в результате воздействия почвенного засоления, превышающего норму для данной культуры, представляет собой своего рода дисфункцию, последствие частичного повреждения фотосинтетического аппарата. С другой стороны, некоторые данные свидетельствуют в пользу адаптивной роли такого снижения.
Показано [6], в частности, что мутации ДНК хлоропластов подсолнечника (линии Chlorina), приводящие к потере 30-40 % хлорофилла листьев приводят к увеличению устойчивости растений к засолению и засухе. Если пла-стомная мутантная линия Chlorina при выращивании на обычной почве проявляет сниженные вегетационные характеристики и урожайность, то в условиях среднего уровня хлоридно-натриевого засоления она способна дать урожай больший, чем нормальная зеленая линия, из которой она была получена. Имеются и прямые данные, демонстрирующие зависимость устойчивости растений от условий протекания фотосинтеза. Так, например, Новиковым [7] показано, что снижение освещенности растений хлопчатника приводит к увеличению их солеустойчивости.
В то же время в известной нам научной литературе отсутствуют экспериментальные данные по непосредственному изучению влияния ограничения освещенности на солеустойчивость подсол- нечника. Результаты таких исследований могли бы прояснить вопрос, является ли индуцированное солевым стрессом снижение фотосинтетической активности этой культуры адаптивным или нет.
Задачей настоящей работы является исследование влияния разных уровней искусственного ограничения освещенности на вегетационные показатели роста и накопление засоляющих ионов растениями подсолнечника в условиях засоления питательной среды 0,5 %-ным NaCl.
Объекты и методы исследований. В экспериментах использовали сорт подсолнечника ВНИИМК 8883 селекции Всероссийского НИИ масличных культур им. В. С. Пустовойта.
Выращивание растений проводили до возраста 25 суток в водной культуре на среде Гельри-геля в соответствии с рекомендациями ВИРа [8]. Искусственное снижение освещенности достигали с помощью применения нейтральных тонированных полиэтиленовых пленок с коэффициентом поглощения 0,3. Коэффициент поглощения измерялся относительно обычной нетонированной пленки. Двойную пленку использовали в случае, если коэффициент поглощения необходимо было довести до 0,49. Светопроницаемость пленок проверялась люксметром. Контрольные растения выращивались под нетонированной прозрачной полиэтиленовой пленкой, коэффициент светопо-глощения которой составлял не менее 98 %. Измеренная освещенность контрольных растений в солнечный день (апрель) на 12.00 – 97000 лк.
Вегетационные показатели растений оценивали по высоте, сухой массе надземной части и площади листьев. Рассчитывали процент гибели растений.
Надземные части растений измельчали, высушивали в термостате при 150 оС до постоянной массы и взвешивали. Отбирали навески сухого материала по 0,5 г и экстрагировали их в течение 1 часа, порциями по 5 мл дистиллированной воды на водяной бане при 90 С. В экстрактах определяли Na+ и Cl– потенциометрическим методом с исполь-зованиием ионселективных электродов ЭЛИС-142 Na и ЭЛИС-131 Cl.
Определяли коэффициент влияния экспериментального фактора Т| , как отношение величины показателя в опыте к соответствующей контрольной величине. В выборках рассчитывали стандартное отклонение для уровня значимости 0,05 и достоверность различий между выборками по Стьюденту.
Результаты экспериментов . Проведенные эксперименты показали, что различные исследуемые факторы (засоление среды NaCl, снижение освещенности) в большинстве случаев способны оказывать воздействие на выживаемость 25-суточных растений подсолнечника.
Так, добавление к жидкой среде выращивания 0,5 %-ного хлористого натрия привело к гибели 4 растений из 30 (13,3 %), тогда как в контрольной выборке такой же численности погибло только одно растение (таблица). Различия в данном случае недостоверны. Снижение освещенности в 30 % (CО 30 %) также приводило к увеличению процента гибели растений (до 16,6 %), причем это уже достоверное отличие (Р = 0,05). Однако причины гибели, вызываемые обоими экспериментальными факторами, оказались неодинаковы; в первом случае (0,5 % NaCl) погибшие растения останавливали рост на самых ранних стадиях прорастания, во втором (СО 30 %) растения поги- бали в интервале с 20 по 25 сутки в основном в результате полегания.
Комбинированное воздействие обоих факторов (NaCl 0,5 % + СО 30 %) приводило к парадоксальному результату; гибель растений сокращалась до уровня контроля.
В то же время более значительное ограничение освещенности (CO 49 %) как само по себе, так и в сочетании с действием 0,5 % NaCl, вызывало значительный рост гибели (см. таблицу).
Следовательно, засоление питательной среды 0,5 %-ным NaCl в условиях эксперимента способно предотвращать гибель растений подсолнечника, вызываемую умеренным снижением уровня освещенности.
Воздействия на растения 0,5 %-ным хлористым натрием и снижением освещенности, если эти факторы влияли по отдельно-
Таблица – Гибель растений* подсолнечника до возраста 25 суток в условиях комбинированного воздействия хлоридно-натриевого засоления и снижения освещенности
|
Вариант опыта |
Гибель растений, % |
|
Контроль |
3,3 |
|
NaCl 0,5% |
13,3 |
|
СО 30% |
16,6 |
|
CO 49% |
50,0 |
|
NaCl 0,5% + СО 30% |
3,3 |
|
NaCl 0,5% + СО 49% |
53,3 |
*Достоверность межгрупповых отличий (уровни значимости Р)
|
Контроль |
NaCl 0,5% |
СО 30% |
CO 49% |
NaCl 0,5% + СО 30% |
NaCl 0,5% + СО 49% |
|
|
Контроль |
- |
>0,05 не достоверно |
0,05 |
<0,001 |
нет отличий |
<0,001 |
|
NaCl 0,5% |
- |
>0,05 не достоверно |
<0,01 |
>0,05 не достоверно |
<0,01 |
|
|
СО 30% |
- |
<0,01 |
0,05 |
<0,01 |
||
|
CO 49% |
- |
<0,001 |
>0,05 не достоверно |
|||
|
NaCl 0,5% + СО 30% |
- |
<0,001 |
NaCl 0,5% – засоление питательной среды хлористым натрием СО 30% и СО 49% – выращивание в условиях сниженной освещенности соответственно на 30 и 49%
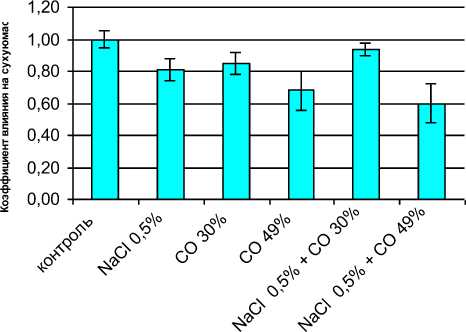
сти, приводили к достоверному (P=0,05) и приблизительно одина- ковому снижению сухой массы растений (рис. 1) по сравнению с контролем.
В то же время одновременное влияние этих факторов позволяло получать растения, сухая масса которых мало и недостоверно отличалась от контроля. Использование 49 % го ограничения освещенности во всех вариантах опытов (с NaCl и без NaCl) давало существенное (на 36-40 %) снижение сухой массы, достоверное как по отношению к контролю, так и по отношению ко всем другим вариантам опыта.
Таким образом, засоление питательной среды 0,5 % NaCl, сочетающееся с 30 %-ным снижением освещенности, обусловливает взаимное ослабление отрицательного действия этих факторов и на показатель сухой массы растений.
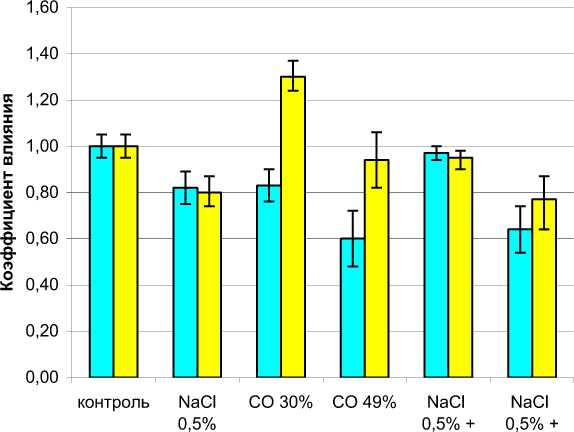
Раздельное воздействие факторов засоления 0,5 %-ным NaCl и снижения освещенности на
30 % приблизительно одинаково снижало площадь листьев расте- ний (рис. 2). Однако на высоту растений эти факторы влияли совершенно противоположным образом. Добавление 0,5 % NaCl в жидкую питательную среду вызывало резкое и достоверное уменьшение высоты растений, а CО 30 % – ее увеличение. Полученные результаты в целом согласуются с известными экспериментальными работами по влиянию почвенного засоления на подсолнечник, в которых показано, что NaCl в сопоставимых концентрациях может снижать высоту растений на 20-30 % [1, 10].
Снижение освещенности, действующее в качестве единственного фактора, обусловливало в
Вариант опыта
Рисунок 1 – Комбинированное воздействие хлоридно-натриевого
данном случае механическую слабость стебля и, как следствие, склонность к полеганию – наблюдаемый эффект также хорошо известен для светолюбивых растений и обусловлен работой фитохромной системы регуляции
СО 30% СО 49%
засоления и снижения освещенности на сухую массу 25-суточных растений подсолнечника, оцениваемую по коэффициенту влияния Т| .
Сухая масса контрольных растений принята за 100%; л = 1,0
NaCl 0,5% – засоление питательной среды хлористым натрием
СО 30% и СО 49% – выращивание в условиях сниженной освещенности соответственно на 30 и 49%
В то же время растения, подвергшиеся комбинированному воздействию 0,5 % NaCl и СО 30 % практически не отличались от контрольных по площади листьев и высоте (см. рис. 2).
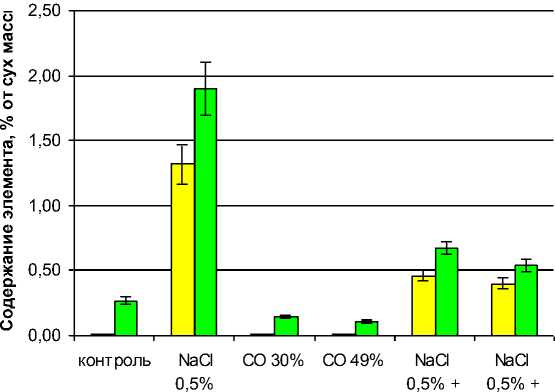
Как следует из данных, приведенных на рис. 3, содержание ионов Na+ и Cl– в листьях растений существенно зависело от их содержания в жидкой питательной среде и от уровня освещенности. Содержание Na в листьях контрольных растений, питательная среда которых не засолялась, составило около 0,01 %, что близко к порогу его обнаружения данным типом ионоселективного электрода. Вероятно, эти остаточные количества натрия поступали в листья в процессе развития из се-
Вариант опыта
-
□ площадь листьев □ высота
Рисунок 2 – Комбинированное воздействие хлоридно-натриевого засоления и снижения освещенности на площадь листьев и высоту 25-суточных растений подсолнечника, оцениваемое по коэффициенту влияния Т| .
Площадь листьев и высота контрольных растений принята за 100 %;
л = 1,0. NaCl 0,5 % – засоление питательной среды хлористым натрием СО 30 % и СО 49 % – выращивание в условиях сниженной освещенности соответственно на 30 и 49 % мени.
Содержание хлора в листьях контрольных растений оказалось достаточно заметным – 0,27 %, что может быть объяснено присутствием этого элемента в жидкой питательной среде в качестве стандартного компонента в концентрации 0,0036 %.
Засоление питательной среды 0,5 %-ным NaCl (эквивалентно
СО 30% СО 49%
Вариант опыта
□ натрий □ хлор
Рисунок 3 – Комбинированное воздействие хлоридно-натриевого засоления и снижения освещенности на накопление ионов Na+ и Cl– 25-суточными растениями подсолнечника NaCl 0,5% – засоление питательной среды хлористым натрием СО 30% и СО 49% – выращивание в условиях сниженной освещенности соответственно на 30 и 49%
В контроле в жидкой питательной среде хлор присутствует в концентрации 0,0036% как компонент питательной среды Гельригеля (KCl); натрий отсутствует
0,195 % Na+ + 0,305 % Cl) приводило к увеличению содержания в листьях натрия до 1,32 и хлора до 1,9 %. Коэффициенты аккумуляции этих ионов составили соответственно 6,80 и 6,16. Следовательно, в условиях эксперимента подсолнечник ведет себя как умеренный галофит аккумуляторного типа, что согласуется с известными данными [10]. Для сравнения следует отметить, что более активные аккумулирующие галофиты способны обнаруживать коэффициент аккумуляции более чем 20 [11].
В то же время снижение освещенности как на 30 %, так и на 49 % вызывало резкое снижение аккумуляции натрия и хлора, причем снижение аккумуляции хлора наблюдалось даже в условиях незасоленного контроля, где этого элемента мало (0,0036 %).
Таким образом, умеренное снижение фотоэкспозиции растений подсолнечника способствовало увеличению его устойчивос- ти к засолению 0,5 %-ным NaCl, оцениваемую по показателям выживаемости, накопления сухой массы, площади листьев и поступления засоляющих ионов в листья.
Характерно, что если каждое из экспериментальных воздействий – засоление и снижение освещенности по отдельности приводит к ухудшению вегетационных показателей, то их комбинированное применение позволяет получать растения, близкие к контрольным.
Обнаруженный синергизм может быть интерпретирован с точки зрения энергозависимого характера развития солевого стресса и солеустойчивости. Так, поглощение корневой системой и ксилемный транспорт ионов Cl– и и Na+ является активным процессом, требующим значительных затрат АТФ. Очевидно, что в случае энергодефицита, определяемого снижением освещенности и, соответственно, снижени- ем уровня фотосинтетически вырабатываемой АТФ, следует ожидать меньшую аккумуляцию засоляющих ионов растениями, что и наблюдается в эксперименте.
Полученные результаты хорошо согласуются с известным фактом существования солеустой-чивости у некоторых пластом-ных мутантных линий подсолнечника Chlorina с пониженным содержанием хлорофилла, на основании чего высказывалось предположение о фотозависимом характере формирования такой солеустойчивости [12]. Это предположение сводится к следующему механизму: снижение уровня фотосинтетически продуцируемых НАДФН и АТФ приводит к зависимому снижению уровня десатурации мембранных липидов, соответствующему увеличению температурной точки фазового перехода мембраны, которое, в свою очередь, может изменять проницаемость клеточных мембран для засоляющих ионов.
Полученные в настоящей работе данные с высокой вероятностью свидетельствуют об адаптивном характере подавления фотосинтеза растений подсолнечника, вызванного умеренным засо-лениием питательной среды. Получено также прямое подтверждение возможности увеличения хлоридно-натриевой солеустойчи-вости подсолнечника путем регулирования фотоэкспозиции.
