Нанайский род Самар: структура генофонда по данным маркеров Y-хромосомы

Автор: Богунов Ю.В., Мальцева О.В., Богунова А.А., Балановская Е.В.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология и палеогенетика
Статья в выпуске: 2 т.43, 2015 года.
Бесплатный доступ
Показано современное расселение нанайского рода Самар (горинская группа нанайцев, Хабаровский край). Изучен генофонд популяции по SNP-маркерам Y-хромосомы. Его основу составляет североевразийская гаплогруппа N1c1-M178 (более 83 %), представлены также еще четыре: C*-М130, I*-M170, J2ala-M47, O2-P31. Мажорная гаплогруппа Nlcl-M178 маркирует преимущественное тунгусское происхождение рода, тогда как полное секвенирование Y-хромосомы образца с минорной гаплогруппой C3*-M130 выявило и связи с коренными популяциями Амура. Рассчитанные генетические расстояния и график многомерного шкалирования демонстрируют значительную генетическую близость нанайского рода Самар с якутами, хакасами и отдельными группами бурят. Это позволяет сделать вывод об их общем происхождении. Нанайцы других регионов находятся на значительном генетическом расстоянии от горинских самагиров.
Родовая структура, род самар, генофонд, y-хромосома
Короткий адрес: https://sciup.org/145145702
IDR: 145145702 | УДК: 575.17 | DOI: 10.17746/1563-0102.2015.43.2.146-152
Текст научной статьи Нанайский род Самар: структура генофонда по данным маркеров Y-хромосомы
Положение Нижнего Приамурья на периферии Восточной Сибири и Тихоокеанского побережья стало решающим фактором в формировании генофонда населения, включившего в себя популяции и таежной, и приморской зоны. По крайней мере с неолита этот регион был зоной миграций народов Сибири и Восточной Азии – небольшие группы тунгусов, тюрок, монголов, маньчжуров смешивались с предшествующим населением. Изучение генофонда нанайцев – самого многочисленного этноса Нижнего Приамурья (ок. 10 тыс. чел.) – с опорой на знание родовой структуры и методы этнической геномики позволяет не только уточнить этногенез, но и выявить различные пласты в генофонде амурского населения, реконструировать исторические процессы на территории Дальнего Востока.
В трудах многих исследователей конца XIX – XX в. нанайская общность предстает как конгломерат разнородных групп с присваивающим типом хозяйства
Археология, этнография и антропология Евразии Том 43, № 2, 2015
без четкой социальной структуры и явных границ, отделяющих его от других сообществ долины Амура [Шренк, 1883; Лопатин, 1922; Смоляк, 1975]. До XIX в. – периода консолидации местных племен и родов в нанайский этнос – все амурское население подразделялось на несколько больших групп по территории проживания и культурно-языковому влиянию: маньчжурскому ( натки, дючеры ), монгольскому ( солоны, дауры ), тунгусскому ( манегиры, бирары, кили ) [Шренк, 1883, с. 11–93; Долгих, 1960]. В XIX в. вышеперечисленные родоплеменные образования исчезли с этнографической карты Приамурья, положив начало родовой организации нанайцев. В XX в. родовые названия закрепились в фамилиях, сохранив прежнюю территориальную привязку. Однако В.А. Туголуков, изучив общую картину этнических взаимоотношений в Приамурье за последние три столетия, заключил, что термин «род» применительно к ме стному сообществу не совсем правильный. На фоне тунгусской экспансии в регионе в XVII в., приведшей к ассимиляции коренного амурского населения и видоизменениям родовых названий, остается подходящим термин «фамилия» [Туголуков, 1972, с. 105]. Но большинство ученых при рассмотрении социальной организации нижнеамурских этносов, в т.ч. нанайцев, все же оперируют понятием «род», что находим и в источниках более раннего времени.
Вливание в инокультурную среду совершалось через институт доха, когда мелкие роды с целью выживания заключали союз между собой. Отношения в образованной патронимии базировались на трудовом и правовом партнерстве между ее членами, предполагали совместные действия в религиозных церемониях [Каргер, 1929, с. 4–8; Смоляк, 1970, с. 266, 288– 289; 2001, с. 14]. В.А. Туголуков, изучая удэгейские и орочские роды, предположил, что институт доха в нижнеамурской среде выполнял функцию связи дочерних родов, имевших общего предка. Его возникновение связано с тунгусской экспансией, буквально перекроившей местную матрилинейную систему родства, которая прослеживалась у нивхов и айнов. Так, материнский (нивхский) род вступал в связь с отцовским (тунгусским). Дети от таких браков, согласно нивхским нормам, не могут жениться в отцовском роде и, согласно тунгусским нормам, – в материнском. Компромиссным вариантом могла стать билинейность (совмещение патрилинейной или матрилинейной) системы родства или поиск третьего рода. У тунгусо-маньчжурских народов как ассимилированного сообщества отцовская родовая организация уживалась с принципами матрилинейности, имевшей местные корни. Выдача вдовы в соседний род, открывавшая путь к экзогамным отношениям в доха, знаменовала переход ее детей в новый отцовский род с дальнейшим запретом для них на брачные связи внутри рода отчима. Вступление в доха даже групп, проживавших далеко друг от друга, указывает на процесс дробления больших локальных общностей на мелкие, их перемешивание и перемещение, вызванное миграцией тунгусов. В результате соседями могли оказаться кровнородственные группы, сблизившиеся через институт доха. Также предполагается, что созданное родовое объединение Самар могло базироваться на изначальном генетиче ском родстве его представителей [Туголуков, 1972].
В настоящей работе показано современное расселение рода Самар в Хабаровском крае, изучен его генофонд по SNP-маркерам Y-хромосомы, а также определена степень генетического родства с другими популяциями нанайцев и народами соседних территорий.
Специфика геногеографических исследований
В последние десятилетия изучение генетического разнообразия популяций человека стало одним из самых актуальных направлений науки. Это обусловлено появлением в арсенале популяционной генетики т.н. однородительских нерекомбинирующих маркеров, локализованных на митохондриальной ДНК и Y-хромосоме человека. Поскольку первые имеют материнский тип наследования, а вторые – отцовский, в совокупности они дают раздельное понятие о вкладе представителей разных полов в историю становления генофонда. В настоящее время в фи-логеографических исследованиях населения маркеры Y-хромосомы исследуются более интенсивно, чем мтДНК, потому что по Y-хромосоме выявляется большее генетическое разнообразие и обнаруживается четкая корреляция с географическим положением и этнической историей населения [Балановская, Балановский, 2007]. Поскольку данная работа основана на использовании маркеров Y-хромосомы, остановимся более подробно на особенностях ее генома и характеристике типов ее полиморфизма.
Протяженно сть Y-хромосомы составляет ок. 60 млн пар нуклеотидов, ее концевые (теломерные) участки вступают в кроссинговер с гомологичными участками X-хромосомы. Все остальное пространство (более 50 млн пар нуклеотидов) занимает группа сцепленных генов, которая не разрушается рекомбинацией и передается из поколения в поколение в виде единого гаплотипа. Новые гаплотипы образуются только путем мутагенеза. Такой механизм возникновения изменчивости позволяет анализировать родство индивидов со сходными последовательностями нерекомбинирующей части Y-хромосомы, что невозможно для рекомбинирующих аутосомных маркеров.
При изучении разнообразия Y-хромосомы исследуют два типа генетического полиморфизма (соответственно два типа маркеров). Однонуклеотидные полиморфизмы (SNP – single nucleotide polymorphism) возникают в Y-хромосоме достаточно часто. Точечные мутации почти никогда не происходят в одной и той же позиции дважды. Исследование SNP позволяет отнести анализируемый образец к той или иной ветви родословного древа Y-хромосомы – гаплогруппе – или ее субветви. Таким образом, мутация является «меткой» для всех последующих потомков и позволяет надежно отличать один «род» от другого. Как правило, у обозначения гаплогруппы (например, N1c1) указывают наименование маркера (индекс), ее определяющего (например, для N1c1 индекс М172). Это связано с тем, что Y-хромосомное древо гаплогрупп постоянно совершенствуется и происходят изменения в их названии, однако маркеры конкретных мутаций не меняются.
При втором типе полиморфизма различия между индивидами определяются по числу аллелей – коротких тандемных повторов (STR – short tandem repeats) в определенных локусах Y-хромосомы. В ряду поколений этот показатель сохраняется, т.е. у ребенка-мальчика, его отца, деда и т.д. будет одинаковый гаплотип Y-хромосомы. Однако все же происходят изменения числа аллелей – возникают новые мутации, причем гораздо чаще, чем SNP, примерно одна на 22 поколения (500 лет).
Обратим внимание на то, что, не зная, к какой гап-логруппе принадлежит исследуемый образец, точно невозможно сказать, действительно ли данный индивид является родственником другого с похожим набором STR-маркеров. Чтобы это проверить, нужно проанализировать SNP-маркеры. Именно поэтому наиболее эффективно параллельное исследование обеих генетических систем (SNP и STR). Их образно сравнивают с часовой и минутной стрелкой: первая (SNP) показывает позицию образца на родословном древе в «глобальном» масштабе, а вторая (STR) – его более точное положение. Исследование STR на фоне данных об SNP позволяет датировать возникновение отдельных кластеров гаплотипов (т.е. ветвей родословного древа Y-хромосомы). Существуют два подхода к анализу SNP-маркеров Y-хромосомы. При первом (традиционном) анализируются отдельные, наиболее информативные маркеры. Второй подход (ставший технически возможным только в самые последние годы) заключается в полной расшифровке нуклеотидной последовательности Y-хромосомы, что позволяет выявить все SNP-маркеры Y-хромосомы, присутствующие у данного образца. В связи с высокой стоимостью полногеномного секвенирования этот подход в нашей работе реализован для одного образца (представителя гаплогруппы С3*-М130), а первый – для всех.
Таким образом, человек обладает набором генов, содержащих информацию о его предках. Весь набор изменений в генофонде популяции отражает очередность генетических преобразований в ряду поколений. Зная среднюю частоту мутаций STR-маркеров Y-хромосомы, можно сравнить спектры гаплотипов разных популяций (или родов), выявить степень их близости и проследить историю изменения Y-гаплотипов. Таким образом, определенные мутации в Y-хромосоме выполняют роль свидетеля исторических процессов: они могут зафиксировать время, миграции, ассимиляцию, метисацию.
Общая структура геногеографических исследований такова: собирают выборку биологических образцов мужчин, не связанных между собой родством по крайней мере на три поколения, но принадлежащих к одной популяции; генотипируют STR- и SNP-маркеры Y-хромосомы; проводят анализ частот гаплогрупп и рассчитывают генетические расстояния; анализируют разнообразие гаплогрупп с последующим фи-логеографическим анализом STR-гаплотипов внутри каждой.
История рода Самар хранится в его генофонде. В ряду множества поколений ассимилировались различные по своему этническому происхождению ветви. Однако с позиции описанного выше методического аппарата современной популяционной генетики изучаемый генофонд не является смесью исторических генофондов. По отцовским линиям Y-хромосомы он хорошо структурируется, что позволяет надеяться на выявление следов не только тунгусской экспансии, но и последующих напластований.
Материал и методы исследований
Материалом для демографических исследований служили данные похозяйственных книг населенных пунктов – мест традиционного проживания нанайцев – трех административных районов Хабаровского края (Солнечный, Комсомольский, Нанайский); для генетических – образцы венозной крови 37 мужчин рода Самар из сельских поселений Кондон ( N = 19), Нижние Халбы ( N = 10), Бельго ( N = 5), Верхняя Эконь ( N = 1), Черный Мыс ( N = 1), Нижняя Там-бовка ( N = 1). В выборку включены индивиды, не состоящие в кровном родстве на глубину не менее трех поколений.
ДНК выделяли фенол-хлороформенной экстракцией. Концентрация ДНК определялась методом ПЦР в реальном времени с использованием набора Quantifiler® (Applied Biosystems). С помощью набора Y-filer™ PCR Amplification Kit (Applied Biosystems) и генетического анализатора 3130×l (Applied Biosystems) анализировали 17 STR-локусов
Y-хромосомы. На основе установленных гаплотипов проводилась предикция гаплогрупп Y-хромосомы с последующим подтверждением генотипированием SNP-маркеров. Такой подход позволял надежно определять гаплогруппу. Дополнительно для образца с гаплогруппой С3*-М130 (×М48) была проведена полная расшифровка нуклеотидной последовательности Y-хромосомы с использованием технологии BigY, разработанной американской генетико-генеалогической компанией FamilyTreeDNA, и определено положение этого образца в общей филогенетической структуре гаплогруппы.
Сравнение генофонда рода Самар с прочими популяциями нанайцев и этносами из других регионов проведено на основе базы данных Y-base (www. . По частотам гаплогрупп с помощью программы DJgenetic рассчитывали генетические расстояния [Nei, 1975], на основе матрицы которых в программе Statistica о суще ствляли многомерное шкалирование, графически визуализирующее степень сходства изученных популяций.
Результаты исследований
Современное расселение рода Самар в Хабаровском крае. Наибольшая численность зарегистрирована в районе исторического проживания – пос. Кондон Солнечного р-на: в 2012 г. 93 мужчины и 111 женщин – 38,3 % от населения поселка. За последнее столетие представители рода Самар расселились и по долине Амура, главным образом в пределах территории Комсомольского р-на: Верхняя Эконь – 3 %, Бельго – 8,76, Нижние Халбы – 18,9 %. В населенных пунктах Нанайского р-на индивиды с фамилией Самар отсутствовали или их доля в родовой (фамильной) структуре населения не превышала 1 %.
«Генетический портрет» рода Самар. В результате генотипирования Y-хромосомы установлено, что «генетический портрет» ныне живущих представителей рода характеризуется наличием пяти гаплогрупп: N1c1-M178, С*-М130, I*-M170, J2a1а-M47, O2-P31. Основу генофонда составляет гаплогруппа N1с1 (более 83 %), значительное преобладание которой можно рассматривать как характерную родовую черту, выделяющую данную популяцию среди других нанайских родов.
Североевразийская гаплогруппа N1с1 с высокой частотой встречается в генофондах народов Сибири: якутов (ок. 90 %), бурят (в отдельных популяциях до 78 %), эвенков (34 %), а также дальневосточных чукчей (61 %), коряков (24 %) [Степанов и др., 2001; Харьков, Степанов, 2005; Харьков и др., 2014]. Накопление данной гаплогруппы у представителей рода Самар свидетельствует о наличии генетической связи изучаемой популяции с северными народами и подтверждает исторические реконструкции миграционных потоков самагиров на Амур.
Высокая частота гаплогруппы J2a1a-М47 (8,1 %) – вторая характерная особенность генофонда рода Самар. По нашим данным, ближневосточная линия J2 не встречается в генофондах остальных родов нанайцев Хабаровского края (неопубликованные данные). Она отмечена у эвенков и эвенов с частотой менее 2 % и не характерна для других народов Сибири. У этносов Средней Азии достигает 12 %.
Гаплогруппа С3*-М130, являясь одним из минорных компонентов в генофонде рода Самар, может маркировать связи с очень широким кругом народов, у которых она зафиксирована. Поэтому для уточнения было проведено полное секвенирование образца с этой гаплогруппой. Обнаружено большое сходство с ее носителями в других популяциях нанайцев и у нивхов и значительное отличие от таковых у иных народов Сибири и Центральной Азии. Таким образом, наличие гаплогруппы С3*-М130 маркирует пусть минорные по частоте, но четко прослеживаемые генетические связи рода Самар с другими популяциями Амура.
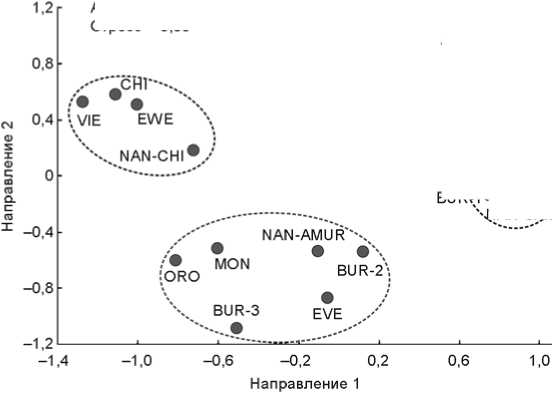
Генетические взаимоотношения между популяциями. Были рассчитаны генетические расстояния между генофондами народов Сибири и Восточной Азии. Использована панель наиболее характерных для изучаемых популяций 13 крупных гаплогрупп, данные по которым присутствуют в литературных источниках. В анализ включены три нанайские выборки: рода Самар (NAN-SAM), суммарной популяции остальных нанайцев Амура (NAN-AMUR) и нанайцев Китая (NAN-CHI). В генетическом пространстве графика многомерного шкалирования выявились три кластера (см. рисунок ), каждый из которых включил одну из нанайских популяций. Это указывает на значительные генетические различия между ними. Нанайцы Китая вошли в единый кластер с эвенами (генетическое расстояние до них d = 0,09), китайцами (0,10 < d < 0,12), вьетнамцами ( d = 0,11). Однако их генофонд обнаруживает меньшее, но выраженное сходство с генофондами монголов ( d = 0,21) и орочей ( d = 0,22). А нанайцы Амура вошли в единый кластер с эвенками ( d = 0,05), монголами ( d = 0,09), бурятами Забайкальского края (0,04 < d < 0,13) и орочами ( d = 0,15).
Генофонд рода Самар наиболее своеобразен: генетическое расстояние между ним и остальными нанайцами Амура ( d = 0,60) почти в 2 раза больше, чем между по следними и нанайцами Китая ( d = 0,36). Столь значительные различия между генофондами нанайцев Хабаровского края связаны с преобладанием у рода Самар североевразийской гаплогруппы N1c1-М217 и низкой частотой С3с-М48. Именно поэ-
ALT
KH-KACI
^IAN-S£M
Алиенация = 0,1
Стресс = 0,08
/'ESKe .-КН-SAG^ : КН-SUM e
Hf ' )»YM
Положение популяций нанайцев в генетическом пространстве народов Сибири и Восточной Азии: график многомерного шкалирования.
ALT – алтайцы; BUR-1 – буряты (Забайкальский край); BUR-2 – буряты (Селенгинский и Хорин-ский р-ны, Республика Бурятия); BUR-3 – буряты (Закаменский р-н, Республика Бурятия); VIE – вьетнамцы; CHI – китайцы [Zhong et al., 2010; Cai et al., 2011]; NAN-SAM – нанайцы рода Самар; NAN-AMUR – нанайцы Амура (Хабаровский край); NAN-CHI – нанайцы Китая [Chen et al., 2006]; MON – монголы [Kim et al., 2011]; ORO – орочи [Xue, 2006]; KH-SUM – хакасы, суммарная популяция; KH-SAG – сагайцы; KH-KACH – качинцы; EWE – эвены; EVE – эвенки [Zhong et al., 2010]; ESK – эскимосы [Lell et al., 2002];
YAK – якуты.
тому, вопреки огромным географическим расстояниям, его генофонд чрезвычайно похож на таковой бурят Забайкальского края ( d = 0,01), якутов ( d = 0,01) и хакасов (0,01 < d < 0,15). Сходство «генетических портретов» этих этносов может указывать на общность некоторых этапов их этногенеза. Неожиданной является относительная близость генофондов рода Самар и эскимосов ( d = 0,18). Указанные популяции образовали единый кластер в генетическом пространстве.
Особенности генофонда рода Самар в целом подтверждают гипотезу историков о происхождении са-магиров от более северных народов Сибири. Согласно этой гипотезе, они являются потомками эвенков, испытавших влияние сначала якутов, а затем (в XVII в.) бурят. Несмотря на то, что доля гаплогруппы N1c1 в генофонде современных эвенков не превышает 34 %, у якутов и бурят наблюдается более значительное ее накопление – 90 и 47 % соответственно (у «восточных» бурят может достигать 78 %) [Харьков, Степанов, Медведева и др., 2008; Харьков, Ха-мина, Медведева и др., 2014]. В изученной нами популяции эвенков (Забайкальский край) частота этой гаплогруппы также невелика (23 %). Однако классическими генетическими маркерами ранее был выявлен чрезвычайно высокий уровень генетического разнообразия данного этноса при огромном ареале и крайне малой численности населения. Это приводит к дрейфу генов, в результате которого возникают значительные различия между генофондами популяций эвенков. Поэтому можно предположить, что истоком генофонда рода Самар была популяция эвенков, отличающаяся более высокими частотами гаплогруппы N1c1. Хотя даже и это не обязательно – основателем рода мог стать один из носителей данной гаплогруп-пы. Относительный вклад эвенков, якутов и бурят можно будет оценить позднее, после анализа вариан- тов гаплогруппы N1c1 – STR-гаплотипов, характерных для генофондов каждого из этих этносов.
Заключение
Усилившийся в XX в. распад родовой организации и установление территориально-соседских отношений народов Амура способствовали размыванию границ внутриэтнических структур, а межэтнические контакты только ускорили ассимиляцию. На этом фоне, а также в условиях невысокой численности приамурских этносов проводить исследования их этногенеза крайне затруднительно. Современная генетика, опираясь на маркеры Y-хромосомы и внутриэтнический уровень структурирования генофонда, открывает новые источники информации, а следовательно, и новые возможности в изучении этносов.
Значительная доля современных нанайцев в настоящее время проживает в условиях иноязычного окружения и имеет тесные контакты с ним, главным образом с русскими. Однако нанайский род Самар (район современного пос. Кондон) в этом отношении уникален. Значительная удаленность от основного ареала нанайцев, а также труднодоступность территории способствовали длительному практически изолированному существованию популяции, а значит, и сохранению исходного генофонда. Напрямую об этом свидетельствуют низкое разнообразие гаплогрупп и преимущественное накопление только одной линии Y-хромосомы.
Установленный в работе «генетический портрет» нанайского рода Самар позволяет говорить о его тунгусских истоках. За длительный период алдано-ленско-амурской миграции к р. Девятке исходный генофонд испытал многократные влияния извне, о чем свидетельствует сходство с генофондами современ- ных якутов и бурят. Археологические, этнографические материалы, письменные источники также указывают на участие народов северо-восточных районов Сибири в этногенезе рода Самар [Малявин, 1998; Медведев, 2005; Медведев, Филатова, 2014, с. 148– 149]. О степени такого влияния можно будет судить в дальнейшем после филогеографического анализа выявленных гаплотипов Y-хромосомы.
Список литературы Нанайский род Самар: структура генофонда по данным маркеров Y-хромосомы
- Балановская Е.В., Балановский О.П. Русский генофонд на русской равнине. - М.: Луч, 2007. - 416 с.
- Долгих Б. О. Родовой и племенной состав народов Сибири в XVII в. // Албазинский уезд. - М.: Изд-во АН СССР, 1960. - С. 579-614.
- Каргер Н.Г. Отчет об исследовании родового состава населения бассейна р. Гарина // Гарино-Амгунская экспедиция 1926 г. - Л.: [б.и.], 1929. - С. 3-24.
- Лопатин И.А. Гольды амурские, уссурийские и сунгарийские. - Владивосток: [б.и.], 1922. - 371 с.
- Малявин А.В. Древние памятники р. Девятки: явление археологической локалии // Project Amur. - Tsubkuba: University of Tsubkuba, 1998. - Р. 53-68.
