Нарушения ритма речи, возникающие в результате синхронизации, индуцированной в связанных осцилляторах

Бесплатный доступ
Оказывается, что заикание, как феномен нарушения ритма речи, можно рассматривать с позиций технической акустики. Как известно, акустически голосовые звуки речи формируются голосовыми складками, периферийными резонаторами и артикуляторами речевого тракта (полость рта, носа, губы, язык, зубы и т. д). Источником энергии возникающих акустических колебаний является воздушный поток из легких. Управление речевой периферией осуществляется, в основном, со стороны мозга. Одной из функций такого управления является формирование начала и конца голосового сегмента в речи. Иными словами, управление формирует ритм речи. Основную электрическую ячейку мозга мы будем называть нейроном. Здесь мы воспользовались простейшей моделью, способной описать электрические свойства нейрона c учетом фазы его состояния: конденсатором (или интегратором) с утечкой, испускающим, по достижению некоторого порога, импульс. Это модель Leaked-Integrator-and-Fire (LIF-модель) [1]. Как известно, потенциал такого интегратора описывается осцилляциями в виде периодической последовательности импульсов с периодом следования ~1мс, поэтому LIF-модель называется также моделью осциллирующего интегратора или просто - моделью осциллятора. Электрически функционирование мозга можно представить себе как некоторое нелинейное взаимодействие таких осцилляторов. Взаимодействие осцилляторов характеризуется некоторой константой связи [2]. Если потенциал в интеграторе превышает некоторый порог, при котором происходит испускание импульса, то этот интегратор называется активным. Активный интегратор может взаимодействовать с соседними интеграторами двояким образом. 1) Активный интегратор может возбуждать временную осцилляцию потенциала в соседнем интеграторе, при этом пиковое значение потенциала превышает порог. В этом случае он называется «возбуждающим» интегратором. 2) Активный интегратор может и «затормозить» потенциал в соседнем интеграторе до уровня ниже порога. Тогда он называется «тормозящим» интегратором. Относительное число «возбуждающих» («тормозящих») в единицу времени интеграторов называется «возбуждающей» («тормозящей») активностью мозга. Активности измеряются в герцах. На периферии эти активности приводят (с помощью специальных клеток) к натяжению мышц голосовых складок, а также к натяжению других мышц речевого тракта, приводящих в движение, например, диафрагму легких, голосовых складок, губ и т. д. Таким образом, описанное управление физически реализует сегментацию речевого сигнала. Другими словами, оно формирует ритм речи. Общепринято считать, что заикание есть феномен отклонения ритма речи заикающегося пациента от ритма нормальной речи. В ходе многолетних экспериментальных исследований мы убедились, что отличаются как средние длительности сегментов, так и их коэффициенты вариации. Экспериментально установленный критический переход от ритма речи с запинками к ритму нормальной речи позволил нам принять гипотезу о том, что сегменты ритма подчиняется квадратичному отображению с управляющим параметром [3, 4]. Исследования детской фонетики, а также изучение различных форм заикания убедило нас в том, что управляющий параметр отображения имеет нейрофизиологический смысл «торможения». Это «торможение» существует в соответствующем звене иерархии нейронных структур, ответственном за ритм, который наблюдается в определенном возрасте или при определенной форме заикания. Закон квадратичного отображения имеет геометрическое место точек, соответствующих устойчивым состояниям отображения, другими словами, устойчивым продолжительностям сегментов ритма. Это геометрическое место устойчивых состояний называется сценарием ритмов, потому что переходы между состояниями соответствуют, в частности, регулярным ритмам детской «лепетной» речи и ритмам повторяющихся запинок ранней формы заикания. Под влиянием все возрастающего тормозного влияния коры мозга ритм нормальной речи становится иррегулярным. Однако при дальнейшем росте торможения появляются запинки невротического характера в виде пауз молчания, персевераций и т. д. Такая форма заикания, сопровождающаяся как запинками в виде повторов, так и невротическими запинками, носит название устойчивого, «хронического» заикания. Именно такое заикание является «камнем преткновения» для врачей-клиницистов, так как для редукции запинок-повторов требуется увеличение торможения в речевых нейронных центрах, а для редукции невротических запинок, наоборот, требуется снижение торможения в речевых центрах [3]. Однако это противоречие исчезает при учете линейной зависимости, существующей между «тормозящей» активностью и управляющим параметром в сценарии ритмов речи [4]. С другой стороны, можно показать, что при учете конечного времени распространения взаимодействия между соседними нейронами-осцилляторами только лишь «тормозящая» популяция осцилляторов способна организовывать синхронные кластеры.
Короткий адрес: https://sciup.org/14316251
IDR: 14316251
Speech rhythm disorders arising as result of synchronization induced in couple oscillators
It is possible to consider a stuttering from point of view of engineering acoustics, if the stuttering is defined as the speech rhythm disorder. Really, voice sounds of speech are formed by voice folds, peripheral resonators and articulators of the speech tract (oral cavity, nose, labiums, tongue, tooth and so on). Energy for acoustic oscillations is supplied by airflow from lungs.
Текст научной статьи Нарушения ритма речи, возникающие в результате синхронизации, индуцированной в связанных осцилляторах
Электронный журнал «Техническая акустика»
В предшествующих работах автора [1–5] было показано, что ритмы вокализаций в речи человека (V-ритмы) образуют на стадии развития фонетики целый набор, или сценарий перехода таких ритмов к хаосу в зависимости от некоторого управляющего параметра. V-ритмом мы называем (регулярное или иррегулярное) чередование длительностей вокализованных сегментов в речи. Физиологическим коррелятом таких сегментов являются эпохи, когда голосовые складки находятся в осциллирующем состоянии. Акустическим коррелятом таких сегментов являются участки непрерывности интонационного контура, то есть зависимости частоты основного тона от времени.
Многочисленные экспериментальные и теоретические обоснования позволили в качестве модели такого сценария принять сценарий ритмов точечного квадратичного отображения логистического типа в зависимости от управляющего параметра в этом отображении:
T n+1 = rT n (1– T n ), n = 1, 2,… (1)
где T n — приведенные к единичному отрезку длительности сегментов V-ритма с номером n , r — управляющий параметр.
В качестве сценария, описывающего переход каскада бифуркаций периода 2 к хаосу, фигурирует сценарий Фейгенбаума перехода к хаосу. Зона хаоса в этом сценарии нами соотносилась с ритмом нормальной речи, а левую границу между бифуркациями удвоения периода и хаосом мы рассматривали ответственной за заикание развития в детском возрасте.
Как оказалось, если рассмотреть в сценарии Фейгенбаума не только окрестность критической точки, но всю эволюцию ритмов, от простейших до сложных, переходящих в хаос, то получается картина, весьма схожая с картиной онтогенеза фонетики [6]. Этот факт позволил вскрыть нейрофизиологическую иерархию стволово-подкорково-корковых структур, и связать их взаимное влияние друг на друга со смыслом управляющего параметра в сценарии Фейгенбаума.
Оказалось, что усложнение ритма по мере роста управляющего параметра возможно связать с торможением, которое имеет место в различных звеньях иерархии нейронных структур, задействованных в процессе онтогенеза фонетики (см. рис. 1). Так, первое усложнение ритма возникает вследствие торможения, оказываемого со стороны стриатума на паллидарные подкорковые ядра, которые обеспечивают существование первичного ритма фонетики ребенка [6]. Дальнейшее, все возрастающее влияние коры на стволовые и подкорковые структуры, ведет ко все большему усложнению ритма, приводящего, в конце концов, динамику ритма в иррегулярную область, описывающую фонетику зрелых речевых процессов.
Известна модель для торможения в нейронных образованиях типа коры головного мозга, предложенная в цикле работ В. И. Сбитнева и В. Г. Яхно [7, 8]. В этих работах исследуется распространение областей повышенной импульсной активности в таких нейронных образованиях. В этой модели в качестве одной динамической переменной используется среднее число ρ т активных волокон, находящихся в «аксонных» деревьях тормозных нейронов, приходящееся на единицу объема и времени, а в качестве другой динамической переменной используется среднее число ρ в активных волокон, находящихся в «аксонных» деревьях возбуждающих нейронов, приходящееся на единицу объема и времени. Оказалось, что динамика этих динамических переменных такова, что допускает существование различных динамических режимов, которые описывают распространение в нейронной структуре зон повышенной активности в триггерном, ждущем и автоколебательном режиме. В автоколебательном режиме Яхно установил наличие волн как тормозной, так и возбуждающей активности с характерным временным размером в 160–140 мсек [8]. Эта величина хорошо согласуется со средними значениями длительностей элементов V-ритма нормальной речи, установленными автором данной работы экспериментально [1–5]. Динамика тормозной и возбуждающей активностей, определяемых по формулам r т = ρ т / P т и r в = ρ в / P в , где плотности тормозных P т и возбуждающих нейронов P в равны соответственно P т ≅ 10-7 мкм-3 и P в ≅ 10-5 мкм-3 [7], была описана с помощью хорошо себя зарекомендовавшей так называемой аппроксимации «среднего поля» [9] для уравнений нейрона Ходжкина-Хаксли [10], или для их упрощенной модели — уравнений ФицХью [11]. Зависимость r т от r в для уравнений ФицХью в приближении «среднего поля» представима в виде предельного цикла, показанного для различных ситуаций на рис. 1. Мы условились называть величину r т уровнем «торможения» в нейронной системе, соответствующей определенному участку в сценарии ритмов, а r в — уровнем «возбуждения» в такой нейронной системе.
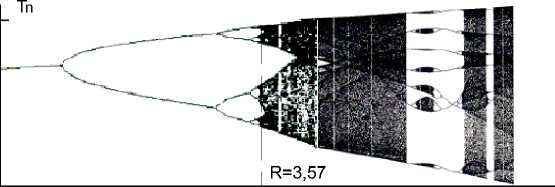
Рис. 1.
rТ
r
В
Вверху: сценарий возникновения речевого иррегулярного ритма в результате усложнения регулярных ритмов при росте торможения в системе. По оси абсцисс отложен управляющий параметр r т (активность «торможения» в нейронной системе, порождающей ритм — подробнее см. текст), по оси ординат отложены возможные в теории нормированные на 1 длительности элементов V-ритма. Управляющий параметр R = 3,57 соответствует границе перехода бифуркаций, ответственных за возникновение регулярных ритмов, в иррегулярный ритм речи. Справа в зоне иррегулярных ритмов видна довольно обширная лакуна, в которой опять возникают бифуркации, но другой природы.
Внизу: в координатах r т (активность «торможения») и r в (активность «возбуждения») представлены предельные циклы, к которым стремится динамика в порождающей ритм нейронной системе. Изображающая точка { r т ; r в } движется против часовой стрелки с течением времени. Жирной линией отмечен предельный цикл для речи в норме; пунктирными линиями отмечены предельные циклы для преходящего заикания у детей (слева) и для невротической компоненты стойкого заикания у взрослых (справа). Два цикла вместе символизируют стойкую фазу заикания, представляющего собой смешанную форму дизритмий тоно-клонического типа и дизритмий невротического типа заикания в переходном и взрослом периоде.
Однако физиологический смысл уровня «возбуждения», в отличие от смысла уровня «торможения» в иерархии нейронных структур, ответственных за возрастную фонетику, оставался неясен.
Здесь следует обратить внимание на то, что вхождение в хаос в сценарии, изображенном на рис. 1, идет по двум полостям, или крыльям:
-
1) по верхнему крылу, «выросшему» из так называемой фазы «гуления», и приводящему к языкам с сингармоническим ритмом (китайский, японский и т. д.); в нейрофизиологии это тоническое крыло принято ассоциировать с правым полушарием;
-
2) по нижнему крылу, «выросшему» из фазы криков, соответствует языкам с силлабическим ритмом (практически все европейские языки); такие ритмы принято соотносить с доминантным, обычно левым полушарием.
0.0
2.0
0.5
Очевидно (см. рис. 2а), что ритм полноценной речи в центре зоны хаоса (соответствующий предельный цикл помечен цифрой 1) соответствует силлабо-тоническим ритмам, которые обеспечиваются взаимодействием обоих полушарий. Рис. 2а можно представить себе как результат поворота нижней части рис. 1, на которой изображены предельные циклы «торможение-возбуждение» вокруг оси абсцисс до тех пор, пока они не «лягут» на сценарий Фейгенбаума, изображенный на верхней части рис. 1.
Tn
r в
1.0
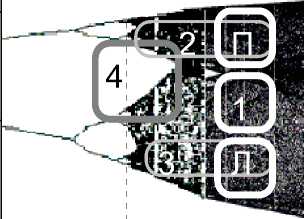
R=3.57
r т
b)
Рис. 2.
a)
-
a) Окрестность границы перехода ритма к иррегулярным ритмам в сценарии V-ритмов, соответствующая дизритмиям тоно-клонического типа и изображенная в координатах: управляющий параметр («торможение») по оси абсцисс; нормированные на единицу длительности элементов V-ритма по оси ординат. Здесь же в координатах: «торможение» по оси абсцисс — «возбуждение» по оси ординат представлены различные предельные циклы «торможения-возбуждения». Белый предельный цикл, помеченный цифрой 1, отвечает ритму нормальной речи. Темно-серый предельный цикл (4) отвечает запиночной речи с запинками тоно-клонического типа. Светло-серый верхний предельный цикл (2), переходящий в белый цикл «П», отвечает тоническим (сингармоническим) иррегулярным ритмам; нижний цикл (3), переходящий в белый цикл «Л», отвечает иррегулярным силлабическим ритмам.
-
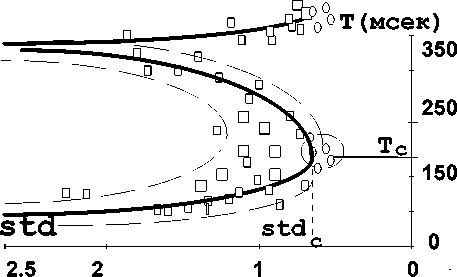
b) Экспериментальные результаты V-сегментации. Значения T c и std c соответствуют нормальной речи. Точки { T, std }, полученные при чтении у взрослых с нормальной речью, отмечены «пустыми» кружками внутри компактного круга в вершине диаграммы при нормальном темпе, и отмечены «пустыми» кружками на конце третьей ветви — при замедленном «распевном» темпе. Серыми квадратами помечены точки, полученные при чтении у заик в процессе обычного курса коррекции (псевдобифуркационная диаграмма) и в процессе коррекции по методу Е. С. Лопотко (третья ветвь диаграммы). На рисунке приведены результаты за один из месяцев, взятый в период исследований, длившихся на протяжении 1993–2003 годов. Более подробные пояснения даны в тексте; различные размеры серых квадратов отражают различные экспериментальные погрешности при экспериментальном вычислении точек для речи различных пациентов.
Допустимость такой трактовки V-ритмов заставила нас провести дополнительные исследования фонетических проявлений межполушарных взаимодействий. Принято считать, что во время пения «задействовано» только лишь правое, сингармоническое полушарие. В НИИ ЛОР (г. Санкт-Петербург) широко используется методика коррекции заикания, предложенная Е. С. Лопотко [12]. Эта методика основана на использовании навыков так называемой «распевной» речи при темпе, несколько замедленным по сравнению с нормой. Такие навыки, как известно, практически целиком редуцируют запинки в речи заикающегося. Наличие такого клинического опыта в институте позволило нам провести сравнительное исследование «распевной» речи с замедленным темпом и речи в обычном темпе как в процессе коррекции речи у заикающихся, так и в контрольной группе лиц без признаков явной речевой патологии.
МЕТОДИКА
Экспериментальную группу составила подгруппа из 22 взрослых заикающихся, успешно прошедших обычный курс коррекции, и подгруппа из 11 взрослых заикающихся, также успешно прошедших курс коррекции, но по методике Е. С. Лопотко. Все пациенты проходили курс коррекции заикания на речевой клинике НИИ ЛОР. Контрольная группа состояла из 8 человек, говоривших как в нормальном темпе, так и в темпе, используемом заикающимися. Пациенты читали стандартный текст объемом 120 слогов. Регистрация речи проводилась трижды за месячный курс коррекции: в начале, в середине и в конце курса.
Речь испытуемых фиксировалась на магнитную ленту с помощью электродинамического микрофона. Процедура перевода электрического сигнала с выхода микрофона в цифровое представление описана в работе [15].
С помощью сертифицированной Минздравом РФ программы сегментации [16], мы определяли длительности голосовых (Voice- или V-) и неголосовых (Unvoice- или U-) сегментов в речи испытуемых заикающихся и вычисляли среднее значение длительностей Т и безразмерный коэффициент вариации std указанных длительностей (отношение среднеквадратичного отклонения к среднему). Точка с координатами { T, std } наносилась на планшет с соответствующими координатными осями (см. рис. 2b). На этом планшете предварительно наносилась компактная область точек { T, std }, соответствующая речи лиц без явной речевой патологии, читавших стандартный текст в нормальном темпе. Заметим здесь в скобках, что приведение теоретического сценария Фейгенбаума, изображенного на рис. 2а, к величинам, фиксируемым нами экспериментально (значение длительностей Т и безразмерный коэффициент вариации std указанных длительностей), приводит к картине, изображенной на рис. 2b (см., например, [13, 14]).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Из рис. 2b видно, что точки { T, std }, полученные при обработке речи пациентов, которые успешно прошли обычный курс коррекции, образуют некоторое множество точек. Среднеквадратичная аппроксимация этого множества представима кривой линией, похожей на бифуркационную линию (так называемая «пуля» Марковица). По мере завершения курса коррекции точки приближаются к области нормы, очерченной окружностью. Как видим, точки для здоровых лиц группируются в вершине указанной кривой. Точки, соответствующие речи пациентов, говорящих в нормальном темпе (серые квадраты — 22 человека) разделяются на две ветви. Пунктирными линиями отмечены области 95% доверительной вероятности (при условии нормального распределения экспериментальных данных). Одна ветвь отвечает речи пациентов со средними длительностями V-сегментов Т , несколько большими длительностей сегментов для нормальной речи. Другая ветвь, наоборот, соответствует средним длительностям, меньшим нормы. Случаи больших коэффициентов вариации std соответствовали, как правило, большей «тяжести» заикания, определенной экспертами. Таким образом, при моделировании процесса ритма необходимо учесть, что заикание должно характеризоваться наличием двух типов V-ритма с большими и меньшими средними длительностями Т для V-сегментов по сравнению с нормальной речью. Первый тип соответствует речи заикающихся, изобилующей запинками тонического типа, например, «со…о…о…о…о…нце»; второй тип соответствует речи заикающихся, изобилующей запинками клонического типа, например, «ку…ку…ку…ку…сты». Назовем эти два типа сингармоническим или тоническим и силлабическим или клоническим типом соответственно. Как мы отметили выше, принято соотносить сингармонический тип вокализаций с правом полушарием, а силлабический тип — с левым, как правило, доминантным полушарием.
Кроме того, на рис 2b, мы видим третью ветвь, полученную в группе, проходившей курс коррекции по методу Лопотко (метод «распевной» речи), причем по мере завершения курса соответствующие точки продвигались к правой оконечности этой ветви. Видим, что эти точки характеризуются средними длительностями V-сегментов, значительно большими, чем в норме, и стремятся, по мере успеха коррекции, к средним значениям длительностей Т для V-сегментов, несколько больших средних значений длительностей Т с для нормы, а соответствующие коэффициенты вариации std к значениям std с , соответствующим норме. При взгляде на рис. 2а видно, что такая ситуация возможна, когда из сценария Фейгенбаума выключается нижняя, силлабическая ветвь сценария, за которую ответственно левое полушарие. В этом случае речи без запинок соответствует белый предельный цикл «торможение-возбуждение», помеченный буквой «П» — правое полушарие.
Вероятно, в условиях, когда из сценария «выключена» верхняя сингармоническая ветвь, за которую ответственно правое полушарие, речи без запинок будет соответствовать белый предельный цикл «торможение-возбуждение», помеченный буквой «Л» — левое полушарие. «Выключение» верхней ветви сценария соответствует, скорее всего, речи, произносимой шепотом. Такой речи, действительно, соответствуют крошечные «островки» фонации в «море» шума. Из такой речи может развиться О. П. Скляров
Сценарий акустических ритмов речи как инструмент исследования работы мозга скандированная, силлабическая речь. Как известно, привитие навыков такой речи также резко редуцирует запинки.
И, наконец, ритму нормальной речи соответствует взаимодействие обеих ветвей: «сингармонической» ветви и «силлабической» ветви с синтетическим «силлабо-тоническим» ритмом нормальной речи. В этом случае, при паритетном взаимодействии обоих полушарий, нормальной речи без запинок будет соответствовать на рис. 2a белый предельный цикл «торможение-возбуждение», помеченный цифрой 1.
Соответствующая модель для описания третьей ветви на рис. 2b была предложена автором еще в ранних работах [2, 3, 13, 14]. Эта модель описывала V-ритм как результат конкуренции двух ритмических процессов, описываемых двумя взаимосвязанными логистическими отображениями типа (1), но с разными, вообще говоря, управляющими параметрами, ответственными за процесс «возбуждения» и «торможения» фонации. Компьютерный расчет показал [2, 3, 13], что асимметрия указанных процессов ведет к возникновению третьей ветви, согласующейся с третьей ветвью на экспериментальном графике 2b. Как мы уже указывали, эта третья ветвь, полученная экспериментально, описывает поведение V-ритма в процессе коррекции методом «распевной» речи.
Таким образом, мы получили как экспериментальное, так и теоретическое свидетельство в пользу наличия асимметрии взаимодействия, скорее всего, межполушарного взаимодействия, в процессе формирования V-ритма «распевной» речи.
ЗАКЛЮЧЕНИЕ
Таким образом, мы видим, что разворачивание сценария Фейгенбаума для V-ритмов речи (см. рис. 1а) как по оси абсцисс, то есть по линии возрастной фонетики, связанной с усложнением ритмической структуры, так и по оси ординат, то есть по линии силлабо-тонического взаимодействия, обусловлено взаимосвязанными реципрокными процессами типа «торможение-возбуждение». Различие лишь в том, что в возрастном аспекте (по оси абсцисс) это торможение обусловлено всей иерархией нейронных структур, и реализуется для каждого участка сценария наиболее высоким представителем иерархии на данной стадии развития по отношению ко всем представителям иерархии, расположенным ниже. Например, первое усложнение ритма возникает в результате торможения, оказываемого стриатумом на паллидарные ядра. Дальнейший переход от регулярных ритмов к иррегулярным ритмам, свойственным, как мы показали ранее в работах [1–5], нормальной речи, происходит в результате торможения со стороны коры на подкорковые структуры и стволовые образования.
Взаимное же влияние нейронных структур при разворачивании сценария Фейгенбаума вдоль оси ординат обеспечивается, по всей видимости, механизмом взаимодействия между структурами одного уровня иерархии. Указанные взаимодействия проявляются в сфере эмоциональных состояний говорящего, например, на ранних стадиях возрастной фонетики в виде появления мажорных или минорных тембров с возникновением триады Якобсона основных гласных звуков речи [а], [и], [у] [6]. В случае иррегулярных V-ритмов силлабо-тонического типа, характерных для развитой речи, таким механизмом взаимодействия является, по всей видимости, механизм межполушарных взаимодействий. Рассмотренные нами экспериментальные данные и предложенная трактовка этих данных оставляют нас в убеждении, что исследование V-ритмов речи является доступным и эффективным инструментом исследования нейрофизиологических механизмов работы мозга при нормальной и нарушенной речи.
Список литературы Нарушения ритма речи, возникающие в результате синхронизации, индуцированной в связанных осцилляторах
- Турбин А. А. Редукция к одномерному уравнению Фоккера-Планка электрической активности ансамбля нейронов. Автореф. дисс....канд. физ.-мат. наук, СПб Политехнический институт, 2005.
- Ernst U., Pawelzik K., Geizel T. Synchronization Induced by Temporal Delay in PulseCoupled Oscillators. Phys. Rev. Lett., vol. 74, №9, 1995. pp. 1570-1573.
- Скляров О. П. Онтогенез речи и сценарий развития ее V-ритмов. Электронный журнал "Техническая акустика", http://www.ejta.org >, 2004, 16.
- Скляров О. П. Элементы теории ритма речи на основе физической феноменологии его нарушений. Автореф. дисс....доктора физ.-мат. наук, СПб Университет, 1999.
- Шеперд Г. Нейробиология. В двух томах. Т.2. (ред. Д. А. Сахаров). М.: "Мир", 1987.
