Наследование цитогенетических и молекулярно-клеточных эффектов в клетках костного мозга животных при хроническом воздействии ионизирующего излучения

Автор: Башлыкова Людмила Анатольевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-3 т.19, 2017 года.
Бесплатный доступ
В экспериментах на лабораторных мышах исследованы эффекты хронического воздействия гамма-излучения в диапазоне доз от 10 до 64 сГр. Показано немонотонное изменение повреждений ДНК, оцененное на молекулярном (ДНК-кометный тест) и цитогенетическом уровнях (микроядерный тест, митотический индекс). У первого поколения мышей, родители которых были облучены в дозах 10, 20 и 30 сГр, отмечен достоверно более высокий уровень двунитевых разрывов ДНК, клеток с микроядрами, понижение митотической активности и усиление апоптоза по сравнению с контролем. У второго-третьего поколений, рожденных от родителей, облученных в дозе 30 сГр, по сравнению с контролем достоверно более низкий уровень двунитевых разрывов ДНК и клеток с микроядрами на фоне снижения митотического индекса и апоптоза. У пятого поколения различия с контролем нивелируются.
Хроническое облучение, доза, мыши линии af, наследование, геном
Короткий адрес: https://sciup.org/148205157
IDR: 148205157 | УДК: 575.1:539.16.04:611.018.41:59
Inheritance of cytogenetic and molecular-cellular effects in cells of animals bone marrow at chronic impact of ionizing radiation
In experiments on laboratory mice effects of chronic influence of gamma radiation in the range of doses from 10 to 64 сGy are investigated. The nonmonotonic change of damages of DNA estimated on molecular (the DNA-cometary test) and cytogenetic levels (micronuclear test, a mitotic index) is shown. At the first generation of mice whose parents have been irradiated in doses 10, 20 and 30 сGy, authentically higher level of double-stranded ruptures of DNA, cells with micronuclei, decrease in mitotic activity and strengthening of apoptosis in comparison with control is noted. At the second or third the generations which are given rise from the parents irradiated in a dose 30 сGy in comparison with control authentically lower level of double- stranded ruptures of DNA and cells with micronuclei against the background of decrease in a mitotic index and apoptosis. At the fifth generation of distinction with control are leveled.
Текст научной статьи Наследование цитогенетических и молекулярно-клеточных эффектов в клетках костного мозга животных при хроническом воздействии ионизирующего излучения
Актуальной проблемой современности является не только изучение изменения стабильности генома соматических клеток, выяснение механизмов его дестабилизации в условиях действия комплекса факторов окружающей среды, в том числе и радиационных, но и исследование возможности наследственной передачи этих изменений. К настоящему времени накоплен ряд экспериментальных и эпидемиологических данных, свидетельствующих о возможности повышения нестабильности генома у потомства облученных родителей [1, 5, 7, 10, 13, 22, 23]. Изучение состояния животных при хроническом облучении в диапазоне малых доз ионизирующей радиации (до 50 сГр) выявило неоднозначность геномного ответа [1, 10].
Цель исследования: с помощью цитогенетических и молекулярных методов изучить стабильность генома лабораторных мышей при хроническом воздействии ионизирующего излучения (ИИ) в диапазоне малых доз и их необлученных потомков, что позволит выявить возможность наследования радиационно-индуцированной нестабильности в ряду поколений этих животных.
Материалы и методы. Эксперименты были проведены на мышах линии Af, которые с двухмесячного возраста содержались в условиях хронического γ-излучения. Использована γ-уста-новка с двумя источниками 226Ra, содержащих 0,474 х 106 и 0,451 х 106 кБк 226Ra. Облучение животных проводили в течение от 1 до 7 месяцев при мощности экспозиционной дозы 150 мкГр/ч. Суммарные дозы облучения животных составили 10, 20, 30, 37, 45, 52 и 64 сГр, которую определяли с помощью дозиметров экспозиционной дозы ДТУ-1 на базе детекторов ДТГ-4. Для получения потомства (F 1 ) от облученных животных были использованы самцы и
самки мышей, которые подвергались облучению в течение 1, 2 и 3 мес., наколенная доза у которых составила 10, 20 и 30 сГр . От мышей, облученных в дозе 30 сГр, получены потомки до пятого поколения. Контрольные пары животных формировались одновременно с опытными парами.
Методы исследования. Изучение генетических эффектов хронического излучения на молекулярном уровне проведены на клетках костного мозга с использованием ДНК-кометного теста, который основан на электрофорезе иммобилизованных в агарозу отдельных клеток. Для анализа уровня повреждения ДНК был использован «нейтральный» вариант метода ДНК-комет, детектирующий двунитевые разрывы (ДР) ДНК [26]. Препараты анализировали на микроскопе AxioSсopе А1 (Carl Zeiss, Германия) с видеосистемой на основе цифровой камеры AxioCam с программой «AxioVision, Release 4.8.2» (Carl Zeiss). На каждом слайде регистрировали по 100 комет. Для анализа и обработки микрофотоизображений ДНК-комет использовали программу СometScore Pro. Количество дву-нитевых разрывов ДНК оценивали по количеству поврежденной ДНК в хвосте кометы, выраженной в процентах (% ТDNA) [25]. Для цитогенетических исследований использован микроядерный тест клеток костного мозга, который является одним из информативных и быстрых способов индикации генотоксичности различных факторов химической и физической природы, основанный на подсчете интерфазных клеток с микроядрами [20]. Препараты КМ для подсчета микроядер (МЯ) готовили из той же взвеси клеток, которую использовали для ДНК-кометного теста. Гипотонию проводили 0,56% раствором KCL (+370C, 10 мин) и фиксировали метанол-уксусным (3:1) фиксатором [16]. Анализ клеток проведен с использованием флуоресцентной микроскопии, что позволило оценить не только частоту клеток с микроядрами, но и митотический индекс (МИ), и долю элиминирующих клеток по типу программированной гибели - апоптозу [17]. Для учета всех типов клеток в 1000 просмотренных (интерфазных, делящихся, с микроядрами, апоптоз-ных) использован лабораторный счетчик «Стимул Плюс С5». Статистическую обработку экспериментального материала проводили с использованием критерия Стьюдента, критерия малых долей, U-критерия Манна-Уитни [11].
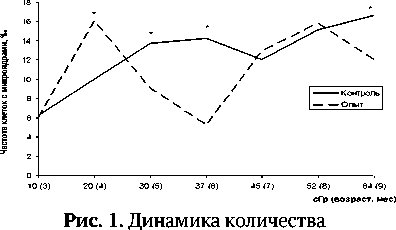
Результаты и обсуждение. В эксперименте с хроническим облучением было задействовано 135 животных. Результаты микроядерного теста, представленные на рис. 1, свидетельствуют о том, что у контрольных животных количество аберрантных клеток с возрастом (от 3 до 5 мес.) увеличивается от 6 до 14‰ и продолжительное время остается на одном уровне (14±2%о). У мышей, испытывающих хроническое облучение, отмечены периоды резкого увеличения и уменьшения количества клеток с микроядрами (от 5,2 до16,2%о), что достоверно выше (при дозе 20 сГр) или ниже (при дозах 30, 37 и 64 сГр) физиологической нормы. То есть у животных в условиях хронического низкодозового воздействия ионизирующего излучения мы наблюдаем немонотонное изменение количества повреждений ДНК, которое свидетельствует о гиперчувствительности при меньших дозах (20 сГр) и радиорезистентности при более высоких дозах (30, 37 и 64 сГр).
Оценка количества двунитевых разрывов ДНК у животных, облученных в дозах 10, 20 и 30 сГр проведена через 4 месяца после окончания облучения (животные использовались для получения потомства). Сравнение количества поврежденной ДНК (в процентах, % TDNA), содержащейся в «хвосте» кометы, показало, что у животных, облученных в дозах 10 и 20 сГр, различий с контролем по количеству ДР не обнаружено (табл. 1). Животные, облученные в большей дозе (30 сГр), имели достоверно меньше ДР, чем контрольные. Сравнение ранжированных данных (табл. 1) по процентному содержанию ДНК в хвосте кометы показало, что у животных, облученных в дозе 10 сГр, по сравнению с контролем понижено количество клеток с неповрежденной ДНК (р<0,001), а у облученных в дозе 30 сГр этот показатель достоверно выше, чем в контроле (5,6 и 2,1%, соответственно, р<0,01). Таким образом снижение количества ДР ДНК у животных облученных в дозе 30 сГр связано с достоверно более высоким содержанием неповрежденных клее-ток (рис. 4), что может свидетельствовать об активации систем репарации ДНК. У мышей, подвергнутых облучению в дозах 10 и 20 сГр, меньше клеток с большим количеством поврежденной ДНК (>50%): у облученных в дозе 20 сГр различие с контролем достоверно (р<0,001), у облученных в дозе 10 сГр -имеет только тенденцию (р=0,06). Клетки с большой долей поврежденной ДНК относят к апоптозным [10, 17]. Поэтому можно предположить, что снижение количества клеток с высокофрагменти-рованной ДНК связано со снижением чувствительности клеток к апоптозу. Несмотря на отсутствие отличий по средним значениям ДР ДНК у животных, облученных в дозах 10 и 20 сГр, по сравнению с контролем, мы наблюдаем различия по уровню клеток с неповрежденной и высоко фрагментированной ДНК.
микронуклеированных клеток костного мозга в зависимости от дозы Y-излучения и возраста лабораторных мышей.* - различия с контролем достоверны при р^ 0,05
Таблица 1. Уровень двунитевых разрывов ДНК по результатам ДНК-кометного теста клеток костного мозга мышей линии Af, подвергнутых хроническому облучению в разных дозах, и их необлученных потомков
|
Покол ения |
Варианты облучения |
N |
% TDNA |
Клетки с неповрежд енной ДНК, % |
Клетки с высоко-фрагментир ованной (>50%) ДНК, % |
|
Fо |
0 |
10 |
23,3±0,3 |
2,1±0,3 |
1,7±0,3 |
|
10 сГр |
5 |
24,6±0,4 |
0,4±0,2*** |
0,7±0,4 |
|
|
20 сГр |
6 |
23,2±0,4 |
1,4±0,5 |
0,5±0,3*** |
|
|
30 сГр |
6 |
19,6±0,5* |
5,6±1,0** |
1,2±0,5 |
|
|
F 1 |
0 |
5 |
15,6±0,6 |
10,2±1,6 |
1,9±0,07 |
|
10 сГр |
9 |
19,3±0,4* |
1,0±0,3** |
2,1±0,04* |
|
|
20 сГр |
8 |
24,7±0,8* |
1,2±0,3*** |
10,9±0,9*** |
|
|
30 сГр |
9 |
25,6±0,5* |
2,1±0,5*** |
4,6±0,7* |
Примечание : * - достоверность отличий от контроля (0) при p<0,05; ** - p<0,01; *** -p<0,001.
Митотическая активность клеток костного мозга контрольных животных с возрастом увеличилась от 1,0 до 4,8%о, а к 8-9 мес. снизилась до 3^ (рис. 2), что соответствует физиологической норме возрастного изменения пролиферативной активности. У мышей, подвергнутых хроническому низкодозовому облучению, данный показатель имел фазовый характер изменений (отмечено два пика и два минимума митотической активности), и амплитуда колебаний была больше, чем у животных из контрольной группы - от 0,8 до 5,4%о. Отметим, что достоверное повышение митотического индекса (в точке 20 сГр) сопровождается увеличением количества клеток с микроядрами, а при снижении этого показателя в точках 37 и 64 сГр наблюдается достоверное снижение доли клеток с МЯ (рис. 1). Снижение пролиферации связывают с остановкой клеточного цикла, необходимой для успешной репарации ДНК и активацией систем контрольных точек клеточного цикла (сверочных точек) [6]. Замедление деления клеток способствует увеличению времени, необходимому для репарации хромосомных аберраций, что в последующем приводит к снижению повреждений ДНК, зарегистрированных нами на клеточном (снижение клеток с микроядрами в точках 37 сГр и 64 сГр) и молекулярном (ДР ДНК) уровнях. То есть, исследуемые показатели - частота клеток с МЯ и митотический индекс клеток костного мозга у облучаемых животных в течение времени резко меняют свои величины вплоть до изменения знака биологического эффекта малых доз ИИ. Это указывает на отсутствие линейной зависимости между дозой хронического низкодозового облучения и его эффектом в клетках костного мозга. Наблюдаемые эффекты свидетельствуют об изменении чувствительности животных в процессе облучения, которая меняется от гиперчувствительности (достоверное повышение частоты МЯ, точка 20 сГр), до радиорезистентности (достоверно более низкая частота клеток с МЯ при дозах 30, 37, и 64 сГр, достоверно более низкий уровень ДР ДНК при дозе 30 сГр).
К настоящему времени известно, что при «слабых» радиационных воздействиях может происходить не только повышение, но и снижение выхода генетических нарушений [4, 18, 19]. Подобные эффекты малых доз ИИ отмечены в натурных исследованиях и экспериментах на разных объектах -клеточных культурах, млекопитающих, у работников атомной промышленности, у пострадавших от инцидентов на объектах ядерной промышленности. В работе [12] отмечено, что у сотрудников Сибирского химического комбината, которые в процессе профессиональной деятельности подвергались хроническому радиационному воздействию, дозовая зависимость частоты хромосомных аберраций (ХА) имела нелинейный характер. При облучении в дозе до 10 мЗв наблюдали достоверное уменьшение частоты ХА по сравнению с контролем, что соответствует известному явлению радиационного гормезиса. Проявление волнообразных изменений генетических эффектов у животных (кур), испытавших воздействие хронического облучения в течение 30 сут., отмечены в работе В.А. Будакова [2]. Установлено, что воспроизводительная способность после введения курам 131I в количестве 0,11 МБк/кг существенно не изменялась, при воздействии 1,1 МБк/кг - повышалась, при 2,1 МБк/кг - после кратковременного повышения отмечено снижение, при 4,6 МБк/кг - угнетение вплоть до необратимого прекращения. Снижение выраженности радиационного эффекта и изменение знака эффекта малых доз ИИ может быть связано с изменением в системе защиты генома - повышение эффективности системы детоксикации свободных радикалов, активации процессов репарации ДНК, апоптоза клеток [3, 6,9].
Проявление радиационно-индуцированных эффектов у потомков облученных животных. Анализ клеток костного мозга показал, что у потомков F 1 , родителей облученных в дозах 10, 20 и 30 сГр, доля клеток с МЯ достоверно выше контроля (рис. 3).
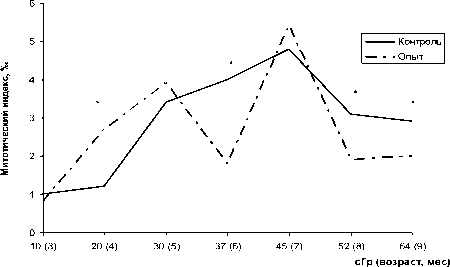
Рис. 2. Изменение митотической активности клеток костного мозга при хроническом низкодозовом облучении лабораторных мышей. * - различия с контролем достоверны при р^ 0,05
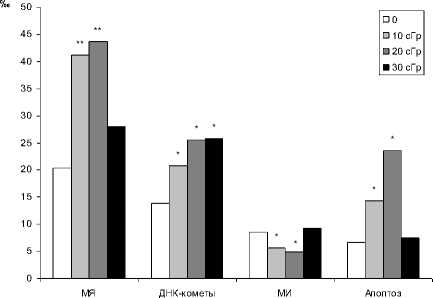
Рис. 3. Результаты исследования эффектов нестабильности генома клеток костного мозга потомков F 1 животных, испытавших воздействие хронического Y-излучения в разных дозах
Более выраженный эффект в отношении нестабильности генома в условиях проведенного эксперимента прослежен у потомков родителей, облученных в меньших дозах (10 и 20 сГр): частота клеток с МЯ в костном мозге в этих группах превышала норму в 2 раза (р<0,01), у потомков родителей, получивших дозу 30 сГр - в 1,3 раза (р<0,05). Исследования на молекулярном уровне показали, что у всех потомков F1 облученных животных наблюдается достоверно более высокий уровень ДР ДНК (рис. 3). По сравнению с контролем у F1 облученных животных происходит снижение в 5-10 раз количества клеток с неповрежденной ДНК и повышение в 2-4 раза клеток с высоко фрагментированной ДНК (табл. 1). Следует отметить, что у потомства облученных родителей в дозах 10 и 20 сГр наблюдается достоверное снижение МИ, что может свидетельствовать об активации репарации ДНК (рис. 3). У первого поколения мышей, облученных в дозах 10 и 20 сГр, отмечен достоверно более высокий уровень апоптоза, обнаруженный на клеточном (рис. 3) и молекулярном уровнях (увеличение клеток с высоко фрагментированной ДНК, табл. 1). C одной стороны, у первого пост-облученного поколения наблюдается повышенный уровень повреждений ядерного материала, определенный на цитогенетическом (МЯ) и молекулярном (ДР ДНК) уровнях, а с другой стороны – усиление защиты генома путем активации процесссов репарации ДНК (замедление пролиферации) и повышение апоптоза (элиминации клеток с нере-парируемыми повреждениями ДНК) [15]. Но их активация недостаточна для поддержания спонтанного уровня повреждений у первого поколения облученных животных. На основе данных литературы и полученных нами результатов можно предположить, что постоянное формирование МЯ и двунитевых разрывов ДНК является следствием состояния повышенного уровня нестабильности генома соматических клеток потомков первого поколения облученных родителей [7, 13, 27]. Предполагают, что повышенный уровень нестабильности генома в соматических клетках потомства является следствием нерепарированных или ошибочно репарированных повреждений генома половых клеток родителей в некий тип изменений, стимулирующих повышенную вариабельность в целом [7, 27].
Оценка нестабильности генома у отдаленных потомков была проведена у второго-пятого поколения (F 2 -F 5 ) мышей, родители которых были облучены в дозе 30 сГр. Обращает на себя внимание то, что выявленный у F 1 достоверно более высокий уровень аберрантных клеток с МЯ и ДР ДНК (рис. 3), в последующих поколениях становится ниже контрольного уровня (табл. 2). У потомков F 2 – F 4 обнаружено достоверное понижение частоты двунитевых разрывов ДНК (%TDNA, табл. 2).
Таблица 2. Цитогенетические и молекулярно-клеточные эффекты в клетках костного мозга потомков (F 2 -F 5 ) мышей линии Af, облученных в дозе 30 сГр
|
Поколения |
N |
МЯ, ‰ |
Апоптоз, ‰ |
МИ, ‰ |
%ТDNA |
|
|
F 2 |
контроль |
9 |
33,7±2,2 |
8,3±1,1 |
2,6±0,6 |
14,3±1,1 |
|
опыт |
10 |
20,1±1,4* |
3,8±0,6* |
1,3±0,4* |
10,9±0,9* |
|
|
F 3 |
контроль |
13 |
40,3±2,5 |
7,2±0,11 |
2,3±0,6 |
16,3±1,3 |
|
опыт |
12 |
34,8±2,9* |
4,8±0,12* |
0,1±0,1* |
12,6±0,9* |
|
|
F 4 |
контроль |
10 |
53,9±2,3 |
12,7±1,1 |
6±0,77 |
20,1±1,2 |
|
опыт |
10 |
51,8±2,26 |
11±1,0 |
8,5±0,91* |
16,4±1,2* |
|
|
F 5 |
контроль |
5 |
16,6±1,8 |
1,4±0,53 |
3,2±0,8 |
14,9±1,4 |
|
опыт |
6 |
15,3±1,6 |
1,3±0,46 |
3,3±0,74 |
14,2±1,4 |
|
Примечание: * - достоверность различий от контроля при p<0,05.
Снижение средних значений, характеризующих величину двунитевых разрывов ДНК, обусловлено достоверным повышением количества клеток с малоповрежденной ДНК, у которых в хвосте кометы фрагментов ДНК меньше 10% (рис. 4). При цитогенетическом исследовании клеток костного мозга F 2 –F 3 выявлено снижение количества клеток с микроядрами и митотического индекса, что может быть обусловлено усиленной репарацией ДНК на фоне снижения пролиферации [3, 6, 9]. По принципу обратной связи снижение повреждений ДНК (ДР ДНК и МЯ в F 2 –F 3 ) приводит к понижению количества клеток, подверженных элиминации (апоптозу) (табл. 2). У пятого поколения мы не обнаружили различий ни на клеточном, ни на молекулярном уровнях исследований.
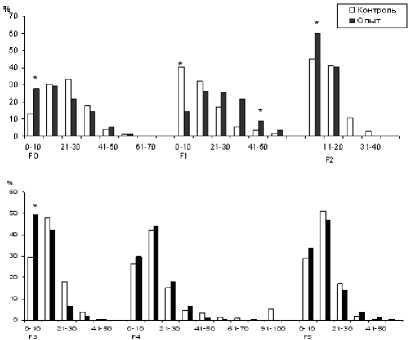
Рис. 4. Гистограмма распределения ДНК в хвосте «кометы» (%ТDNA) клеток костного мозга мышей линии Af после хронического γ-облучения в дозе 30 сГр и их необлученных потомков (F 1 -F 5 )
Выводы: можно сделать заключение об эффективности хронического воздействия ИИ в малых дозах. Наблюдаемые в области низких доз генетические эффекты обусловлены не столько повреждающим действием ионизирующего излучения, сколько особенностями реализации ответной реакции клетки на слабые внешние воздействия [8]. В условиях хронического низкодозового воздействия ионизирующего излучения наблюдается немонотонные изменения количества повреждений ДНК в виде двунитевых разрывов ДНК и клеток с микроядрами, а также непостоянство пролиферативной активности (митотический индекс). Это свидетельствует об отсутствии линейной зависимости между дозой хронического низкодозового излучения и его эффектом в клетках костного мозга. Обнаруженные эффекты свидетельствуют об изменении чувствии-тельности животных к ИИ в процессе облучения, которая меняется от гиперчувствительности до радиорезистентности. Изменение знака эффекта малых доз ИИ может быть связано с изменением в системе защиты генома - повышение эффективности системы детоксикации свободных радикалов, активации процессов репарации ДНК, апоптоза клеток [3, 6, 9]. У потомков F1 облученных родителей выявлено наследование нестабильности генома, о чем свидетельствует увеличение числа клеток с МЯ и ДР ДНК. Зависимость эффектов радиационно-индуцированной нестабильности генома (РИНГ) от дозы облучения на клеточном и молекулярном уровнях имеют нелинейный вид - большую эффективность в первом поколении проявили более низкие дозы (10 и 20 сГр по сравнению с 30 сГр). Полагают, что феномен РИНГ является следствием неточной репарации повреждений структуры генома [7, 27]. Особенностью этого эффекта являя-ется эпигенетический характер их наследования, то есть наследование измененного состояния генной экспрессии [9, 21]. Изменение таких показателей клеточного гомеостаза как митотический индекс и апоптоз направлены на поддержание целостности генома. В отличие от первого поколения, у отдаленных потомков облученных мышей отмечено снижение частоты ДР ДНК (F2-F4) и клеток с микроядрами (F2-F3) ниже спонтанного уровня. У этих животных понижены также показатели, свидетельствующие об изменении пролиферативной активности и элиминации клеток, что может свидетельствовать о перестройке работы системы защиты клеток, которая к тому же передается последующим поколениям. Вероятно, переданная по наследству нестабильность генома потомкам облученных животных, выражается в активации механизмов защиты генома – процессов репарации и апоптоза.
Работа выполнена при поддержке проекта Президиума УрО РАН (№ 15-4-4-20).
Список литературы Наследование цитогенетических и молекулярно-клеточных эффектов в клетках костного мозга животных при хроническом воздействии ионизирующего излучения
- Ахматулина, Н.Б. Отдаленные последствия радиации и индуцированная нестабильность генома//Радиационная биология. Радиоэкология. 2005. Т. 45, №6. С. 680-687.
- Будаков, В.А. Оценка репродуктивного здоровья кур и их потомков при хроническом действии 131I, введенного в организм родителей//Радиационная биология. Радиоэкология. 2015. Т. 55, №3. С. 267-281.
- Бурлакова, Е.Б. Особенности действия сверхмалых доз биологически активных веществ и физических факторов низкой интенсивности//Российский химический журнал. 1999. Т. 43, №5. С. 3-11.
- Бычковская, И.Б. Немутагенные немишенные радиационные эффекты. Наследуемое снижение жизнеспособности клеток, индуцированное лучевыми воздействиями в малых дозах//Радиационная биология. Радиоэкология. 2013. Т. 53, №3. С. 246-258.
- Воробцова, И.Е. Трансгенерационная передача радиационно-индуцированной нестабильности генома//Радиационная биология. Радиоэкология. 2006. Т. 46, №4. С. 441-446.
- Газиев, А.И. Низкая эффективность репарации критических повреждений ДНК вызываемых малыми дозами радиации//Радиационная биология. Радиоэкология. 2011. Т. 51, №5. С. 512-529.
- Дуброва, Ю.Е. Нестабильность генома среди потомков, облученных родителей. Факты и интерпретация//Генетика. 2006. Т. 42, №10. С. 1335-1347.
- Евсеева, Т.И. Сочетанное действие факторов радиационной и нерадиационной природы на традесканцию. Монография/Т.И. Дуброва, С.А. Гераськин. -Екатеринбург: УрО РАН, 2001. 156 с.
- Жижина, Г.П. Влияние малых доз низкоинтенсивной ионизирующей радиации на структуру и функции ДНК//Радиационная биология. Радиоэкология. 2011. Т. 51, №2. С. 218-228.
- Кузнецова, E.А. Индукция редко-и плотноионизирующими излучениями повреждений ДНК в лейкоцитах крови и цитогенетических повреждений в полихроматофильных эритроцитах костного мозга мышей и их потомков/Е.А. Кузнецова, С.И. Заичкина, Н.П. Сирота и др.//Радиационная биология. Радиоэкология. 2014. Т. 54, №4. С. 341-349.
- Лакин, Г. Ф. Биометрия. 3-е изд. -М.: Высшая школа, 1980. 352 с.
- Литвяков, Н.В. Частота и спектр цитогенетических нарушений у работников сибирского химического комбината/Н.В. Литвяков, М.Б. Фрейдин, М.В. Халюзова и др.//Радиационная биология. Радиоэкология. 2014. Т. 54, №3. С. 283-296.
- Ломаева, М.Г. Влияние ионизирующей радиации на уровень полиморфизма ДНК в разных тканях у потомства облученных мышей. Автореф. дисс… к.б.н. -М.: 2008. 24 с.
- Ломаева, М.Г. Повышенная вариабельность генома в соматических клетках у потомства самок мышей, подвергнутых острому рентгеновскому облучению в преконцептивный период/М.Г. Ломаева, Г.В. Васильева, Л.А. Фоменко и др.//Генетика. 2011. Т. 47, №10. С. 1371-1377.
- Михайлов, B.Ф. Молекулярные проявления радиационно-индуцированной нестабильности генома: возможность химической модификации/В.Ф. Михайлов, В.К. Мазурик, Е.Б. Бурлакова и др.//Радиационная биология. Радиоэкология. 2005. Т. 45, №5. С. 561-570.
- Орлов, В.Н. Сравнительная цитогенетика и карио-систематика млекопитающих/В.Н. Орлов, Н.Ш. Булатова. -М.: Наука, 1983. С. 21-28.
- Тронов, В.А. Спонтанная гибель мононуклеарных клеток, полученных от здоровых доноров и больных системной красной волчанкой/В.А. Тронов, Т.А. Никольская, М.А. Конопляников и др.//Цитология. 1999. Т. 41, №5. С. 400-404.
- Шмакова, Н.Л. Индукция хромосомных аберраций и микроядер в лимфоцитах периферической крови человека при действии малых доз облучения/Н.Л. Шмакова, Е.А.Насонова, Е.А. Красавин и др.//Радиационная биология. Радиоэкология. 2006. Т. 46, №4. С. 480-487.
- Grigorkina, E. Radioadaptation of rodents in the zone of local radioactive contamination (Kyshtim Accident, Russia): 50 years on/E. Grigorkina, G. Olenev//Radioprotection. 2009. V. 44, No 5. P. 129-134.
- Hedlle, J.A. A rapid in vivo test for chromosomal damage//Mutation Research. 1973. V. 18. Р. 187-190.
- Lorimore, S. A. Radiation-induced genomic instability and bystander effects: inter-related nontargeted effects of exposure to ionizing radiation/S.A. Lorimore, P.J. Coates, E.G. Wright//Oncogene. 2003. V. 22. P. 7058-7069.
- Maxwell, С.А. Targeted and nontargeted effects of ionizing radiation that impact genomic instability/S.A. Maxwell, M.C. Fleisch, S.V. Costes et al.//Cancer Res. 2008. V. 68, №20. P. 8304-8311.
- Mohr, U. Possible carcinogenic effect of γ-rays in a transgenerational study СВА mice/U. Mohr, T. Tillmann, K. Kamino et al.//Carcinogenesis. 1999. V. 20, №2. С. 325-332.
- Mothersill, C. Radiation_induced bystander effects: do they provide evidence for an adaptive response?/C. Mothersill, C. Seymour//Int. J. Low Radiat. 2006. V.2. №1/2. P. 119-127.
- Olive, P.L. Induction and rejoining of radiation induced DNA single-strand breaks: «tail moment» as a function of position in the cell cycle/P.L. Olive, J.P. Banath//Mutat Res. (DNA Repair). 1993. V. 294. P. 275-283.
- Singh, N.P. A simple technique for quantitation of low levels of DNA damage in individual cell/N.P. Singh, M.T. McCoy, E.L. Schneider//Exp. Cell Res. 1988. V. 175. P. 184-191.
- Skinner, M.K. What is an epigenetic transgenerational phenotype? F3 or F2//Reproductive Toxicology. 2008. V. 25. P. 2-6.
