Наследование длины и диаметра венчика у трубчатых цветков подсолнечника

Автор: Демурин Я.Н., Рубанова О.А.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 1 (181), 2020 года.
Бесплатный доступ
Некоторые морфологические признаки трубчатых цветков подсолнечника влияют на процесс пчёлоопыления во время цветения корзинки, что делает их значимыми для селекции на высокую семенную продуктивность. К ним относится длина венчика, определяющая доступность нектара, и диаметр венчика, потенциально влияющий на количество нектара. В 2017-2019 гг. изучали изменчивость и наследование этих признаков в линиях генетической коллекции ВНИИМК, г. Краснодар. Длину венчика измеряли от его основания до конца зубчиков. В генетической коллекции подсолнечника содержатся линии с наследственно контролируемыми различиями по длине венчика от 6,3 до 9,2 мм и его диаметру от 1,4 до 3,6 мм. Изменчивость длины венчика оценивается как слабая (CV = 9 %) с долей влияния фактора генотипа 96 %, а диаметра - как средняя (CV = 13 %), с долей влияния генотипа 94 %. Между длиной и диаметром венчика в линиях генетической коллекции не установлено достоверной корреляции (r = 0,12
Морфологический признак, трубчатый цветок, наследование, аддитивность, олигогены
Короткий адрес: https://sciup.org/142223398
IDR: 142223398 | УДК: 633.854.78:575 | DOI: 10.25230/2412-608X-2020-1-181-12-20
Inheritance of the corolla length and diameter in tubular florets of sunflower
Some morphological features of tubular sunflower florets affect the process of bee pollination during flowering, which makes them significant for breeding for high seed productivity. These include the length of the corolla, which determines the availability of nectar and the diameter of the corolla circle, potentially affecting the amount of nectar. In 2017-2019 the variability and inheritance of these traits in the lines from the genetic collection of VNIIMK, Krasnodar were studied. The corolla length was measured from the base to the end of the cloves. The genetic collection of sunflower contains lines with hereditarily controlled differences in corolla length from 6.3 to 9.2 mm and circle diameter from 1.4 to 3.6 mm. Variation in the length of the corolla is estimated to be low (CV = 9%) with a share of the influence of the genotype factor 96%, and the diameter of the circle - as moderate (CV = 13%) with a share of the influence of the genotype 94%. We established no reliable correlation between the length and the diameter of the corolla in the lines from the genetic collection (r = 0...
Текст научной статьи Наследование длины и диаметра венчика у трубчатых цветков подсолнечника
Введение . Подсолнечник относится к типичным перекрёстноопыляемым видам растений. Наличие насекомых-опылителей необходимо для реализации потенциальной продуктивности растений. При недостаточном опылении подсолнечника потери урожайности могут достигать более 25 % за счет снижения завязываемо-сти семян [1].
При селекционно-семеноводческой работе с использованием системы ЦМС-Rf у подсолнечника существует проблема переноса пыльцы на стерильную форму с фертильных отцовских (получение гибридных семян) или фертильных материнских (размножение линии) растений. В этих случаях процесс завязывания семян практически полностью зависит от наличия насекомых-опылителей [2].
Некоторые морфологические признаки трубчатых цветков подсолнечника влияют на процесс пчёлоопыления во время цветения корзинки, что делает их значимыми для селекции на высокую семенную продуктивность. Прежде всего, это длина венчика, коэволюционно связанная с длиной пчелиного хоботка (доступность нектара), а также, вероятно, диаметр кольцеобразного вздутия в нижней части венчика, потенциально определяющий размер нектароносной ткани (количество нектара).
При изучении селекционных линий ВНИИМК установлено, что размеры венчика, измеряемые от его основания до нижней кромки зубчиков, изменялись от 3,5 до 4,8 мм. При этом длина венчика трубчатого цветка оказывала наиболее существенное влияние на привлекатель- ность отдельных генотипов для насекомых. Так, линии с укороченным венчиком (в интервале 3,5–4,0 мм) наиболее охотно посещались пчёлами. При увеличении размеров венчика свыше 4,2 мм пчёлопо-сещаемость снижалась, а минимальная аттрактивность растений к пчёлам (до 10 особей на корзинке в час) отмечена у линии, имеющей венчик с максимальной длинной 4,8 мм. Коэффициент корреляции между пчёлопосещаемостью и размером цветка составил -0,78 [3].
Получены экспериментальные данные о значимости количества, качества и доступности нектара как факторов аттрак-тивности подсолнечника по отношению к опылителям. При этом признаки трубчатого цветка, включая длину венчика, зависят как от генотипа, так и от факторов окружающей среды [4; 5].
В ряде исследований установлено, что генотипы подсолнечника, имеющие меньшую длину трубчатого цветка, наиболее привлекательны для опылителей. При средней длине хоботка рабочей пчелы в 6,5 мм, предпочтение в виде более частого посещения пчёлами отдавалось генотипам подсолнечника, венчики цветков которых имели длину 4,8–5,5 мм. Также сообщается о значительной отрицательной корреляции (-0,65) между этим морфологическим признаком и частотой пчёлопосещений [6; 7].
С другой стороны, в некоторых опытах ВНИИМК не было обнаружено влияния длины венчика цветка на привлекательность пчёлам. Так, например, цветки линии ВК678 с укороченным венчиком 3,7 мм хорошо посещались пчёлами, однако у линии 57R с такой же длиной венчика отмечена минимальная посещаемость [8], что указывает на влияние других факторов.
У 100 инбредных линий наблюдали значительную изменчивость длины венчика от 6,8 до 9,9 мм. При этом уменьшение длины на 2 мм более чем удваивало посещаемость опылителями [9]. Имеются данные о высокой наследуемости призна- ка длины венчика трубчатого цветка и отмечено частичное доминирование в F1 родителя с меньшими значениями признака [4; 10].
В целом, в отличие от краевых ложноязычковых цветков, наблюдается недостаточное количество данных о генотипической изменчивости и, особенно, закономерностях наследования морфологических признаков венчика трубчатых цветков у подсолнечника.
Материалы и методы . Исследования проводили в 2017–2019 гг. на ЦЭБ ВНИИМК, г. Краснодар. Объектом исследования были растения культурного подсолнечника H. annuus L. Всего изучали 90 линий генетической коллекции различного происхождения: К223, К225, К254, К370, К562, К581, К651-3, К752, К912, К1459, К1506, К1594, К1687, К2068, К2086, К2125, К2235, К2238, К2257, К2462, К2479, К2645, К3035, К3059, К3159, К3350, К3376, Сл1721, Сл1787, Сл1790, Сл1813, Сл2039, Сл2950, ВА1, ВА4, ВИР130-1, ВИР130-3, ВИР172, ВИР391, ВИР369, ВК15, ВК30, ВК98-2, ВК102, ВК206, ВК268, ВК310, ВК416, ВК428, ВК464, ВК474, ВК475, ВК519, ЛГ3, ЛГ8-2, ЛГ10, ЛГ26, ЛГ27, ЛГ28, КГ7, КГ15, КГ16, КГ19, КГ21, КГ32, КГ48, КГ49, КГ104, КГ111, КГ113, КГ115, Л1392, Л2090, Л2138, Л2532, Л2543, Л2563, Л2582, Л2586, Л2595, Л3376, Л7247, И7-235, И7-249, МВГ-3, МВГ-8, № 424924, № 577083, № 577432 и № 577433.
Растения выращивали на однорядных делянках при схеме посева 70 × 35 см по 25 растений в ряду на селекционном поле лаборатории генетики отдела подсолнечника.
Сбор трубчатых цветков одного дня цветения, проводили из средней зоны корзинки через 3–4 дня после начала её цветения. Для каждой линии собирали 25 цветков (по 5 штук с 5 корзинок). Цветки собирали в герметично закрываемые пластиковые ёмкости объемом 30 мл. Измерение признаков венчика осуществляли в 14
день сбора с помощью штангенциркуля. Полную длину венчика определяли от его основания (в месте прикрепления к завязи и расположения нектарника) до дистальной части с учетом размеров зубчиков после раскрытия цветка, т.е. в пчёлопосещаемую фазу его развития. Диаметр венчика измеряли в его самой широкой нижней кольцеобразной части (рис. 1).
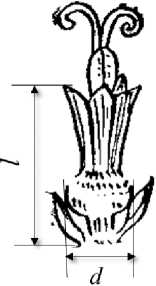
Рисунок 1 – Трубчатый цветок подсолнечника l – длина венчика от основания до дистальной части лепестков;
d – диаметр венчика
В осенне-зимний период 2017–2018 гг. в тепличном комплексе проведены реципрокные скрещивания отобранных линий с контрастными значениями признаков с использованием ручной кастрации. В 2018 г. семена F 1 и родительских линий высевали в поле, для получения семян F 2 проводили принудительное самоопыление корзинок под индивидуальными сетчатыми изоляторами.
В 2019 г. в поле были высеяны родительские линии, гибриды F 1 и F 2 . Для линий и гибридов F 1 измеряли по 25 трубчатых цветков. Для оценки длины и диаметра венчика у гибридов F 2 отбирали по 10 трубчатых цветков с 40 корзинок в пределах 4-рядных делянок.
Значение степени доминирования признака в F1 рассчитывали по формуле отношения h/d, где h – отклонение фенотипа F1 от среднего (m) между ранжированными родителями P1 и P2, а d – половина разности между родителями P1 и P2 или модуль разности любого родителя и m (рис. 2).
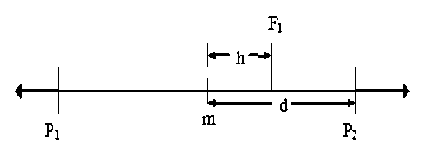
Рисунок 2 – Степень доминирования признака h/d в F 1
Погодные условия в 2017–2019 гг. были в целом благоприятными для развития растений подсолнечника.
Статистическую обработку данных проводили с использованием пакета программ Excel (анализ вариационного ряда, дисперсионный и корреляционный анализы).
Результаты и обсуждение . Изученные в 2017 г. линии генетической коллекции подсолнечника показали варьирование длины венчика трубчатого цветка от 6,3 у КГ49 до 9,2 мм у Л2138, т.е. с различием в 1,5 раза (рис. 3). Среднее значение при этом составило 7,5 мм, размах варьирования – 2,9 мм с невысоким коэффициентом вариации CV = 9 %.
9 >9,2 •
-
8,7- 8,9 _„_|
-
8,4- 8,6 _j
-
8.1- 8,3 ,
3 7,8-8,0 ЙЙЙЙҺніННЯВ
37,5-7,7 еняя^яецбеняняв
7,2-7,4 ■■^■■■■■■■■^■■В
6,9-7,! бййбйййяййййГ
6,6-6.8
6,3-6,5!
’ ’и
О 5 10 15 2025
Рисунок 3 – Изменчивость длины венчика трубчатых цветков у линий генетической коллекции (n = 90), ЦЭБ ВНИИМК, г. Краснодар, 2017 г.
Трехлетнее наблюдение в 2017–2019 гг. за пятью отобранными линиями с контра- стными значениями длины венчика Л2138, К581, ВК905 (max) и ЛД102, КГ49 (min) показало сильное влияние генотипа на варьирование признака без смены ежегодных фенотипических рангов линий. В среднем за три года длина венчика по двум группам линий (max и min) составила от 8,4 до 8,9 мм и от 6,3 до 6,7 мм соответственно (табл. 1). Доля влияния фактора генотипа по результатам дисперсионного анализа оценивается в 96 % (табл. 2).
Таблица 1
Длина венчика трубчатого цветка у линий генетической коллекции подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2017–2019 гг.
|
Лимит |
Линия |
Год |
Среднее по годам |
НСР 05 |
||
|
2017 |
2018 |
2019 |
||||
|
Л2138 |
9,2 |
8,7 |
8,7 |
8,9 |
0,1 |
|
|
Мax |
К581 |
8,8 |
8,6 |
8,6 |
8,7 |
0,1 |
|
ВК905 |
8,3 |
8,3 |
8,5 |
8,4 |
0,1 |
|
|
Мin |
ЛД102 |
6,7 |
6,7 |
6,7 |
6,7 |
0,1 |
|
КГ49 |
6,3 |
6,3 |
6,3 |
6,3 |
0,1 |
|
|
НСР 05 |
0,2 |
0,1 |
0,1 |
|||
Таблица 2
Результаты двухфакторного дисперсионного анализа длины венчика трубчатого цветка у линий генетической коллекции подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2017–2019 гг.
|
Источник вариации |
SS |
df |
MS |
F |
F кри-тиче-ское |
Доля влия ния, % |
|
Год |
2,40 |
2 |
1,20 |
30,38 |
3,03 |
0,6 |
|
Генотип |
394,37 |
3 |
131,46 |
3327,52 |
2,64 |
96,0 |
|
Взаимодействие |
2,29 |
6 |
0,38 |
9,68 |
2,13 |
0,6 |
|
Внутри |
11,38 |
288 |
0,04 |
2,8 |
||
|
Итого |
410,44 |
299 |
100 |
Примечание: SS – сумма квадратов;
df – степени свободы, MS – средний квадрат;
F – критерий Фишера
При реципрокном скрещивании линий с максимальными значениями длины венчика в 2018 г. max × max (табл. 3) наблюдали наследование в F1 по типу отрицательного сверхдоминирования (h/d от -3,00 до -2,18), отрицательного доминирования (-0,50) и промежуточного наследования (от 0,00 до 0,33). В 2019 г. установлено отрицательное сверхдомини-15
рование (h/d от -7,00 до -2,00) и отрицательное доминирование (-1,00) (табл. 4).
Таблица 3
Наследование в F 1 длины венчика при скрещивании линий подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2018 г.
|
Комбинация скрещивания |
Генотип |
Длина венчика, мм |
НСР 05 |
h/d |
|||
|
Р 1 |
Р 2 |
F 1 |
F 1 по группе |
||||
|
К581 × Л2138 |
8,6 |
8,7 |
8,5 |
0,09 |
-3,00 |
||
|
Л2138 × К581 |
8,7 |
8,6 |
8,5 |
0,09 |
-3,00 |
||
|
ВК905 Б × К581 |
8,3 |
8,6 |
8,1 |
8,4 |
0,01 |
-2,18 |
|
|
К581 × ВК905 Б |
8,6 |
8,3 |
8,5 |
0,10 |
0,33 |
||
|
Max × max |
ВК905 Б × Л2138 |
8,3 |
8,7 |
8,5 |
0,10 |
0,00 |
|
|
Л2138 × ВК905 Б |
8,7 |
8,3 |
8,4 |
0,13 |
-0,50 |
||
|
Л2138 × ЛД102 |
8,7 |
6,7 |
8,0 |
0,11 |
0,30 |
||
|
Л2138 × КГ49 |
8,7 |
6,3 |
7,3 |
0,10 |
0,00 |
||
|
Max × min |
ВК905 Б × ЛД102 К581 × ЛД102 |
8,3 8,6 |
6,7 6,7 |
7,6 7,7 |
7,6 |
0,10 0,10 |
0,13 0,05 |
|
К581 × КГ49 |
8,6 |
6,3 |
7,3 |
0,12 |
-0,13 |
||
|
ВК905 Б × КГ49 |
8,3 |
6,3 |
7,8 |
0,12 |
0,50 |
||
|
ЛД102 × ВК905 Б |
6,7 |
8,3 |
8,0 |
0,12 |
0,00 |
||
|
ЛД102 × Л2138 |
6,7 |
8,7 |
8,1 |
0,10 |
0,40 |
||
|
Min × max |
ЛД102 × К581 КГ49 × ВК905 Б |
6,7 6,3 |
8,6 8,3 |
7,8 7,7 |
7,7 |
0,11 0,12 |
0,16 0,40 |
|
КГ49 × Л2138 |
6,3 |
8,7 |
7,4 |
0,11 |
0,24 |
||
|
КГ49 × К581 |
6,3 |
8,6 |
7,4 |
0,10 |
-0,04 |
||
|
Min × min |
ЛД102 × КГ49 |
6,7 |
6,3 |
6,6 |
6,8 |
0,09 |
0,50 |
|
КГ49 × ЛД102 |
6,3 |
6,7 |
6,9 |
0,10 |
2,00 |
||
В 12 комбинациях реципрокных скрещиваний линий с максимальным и минимальным значениями длины венчика в 2018 г. (табл. 3) наблюдали в 11 случаях наследование в F 1 по промежуточному типу (h/d от -0,13 до 0,40), а в одном случае (ВК905 Б × КГ49) отмечено неполное доминирование (0,50). В 2019 г. (табл. 4) во всех комбинациях установлено только промежуточное наследование (h/d от -0,13 до 0,46).
При реципрокном скрещивании линий с минимальными значениями длины венчика в 2018 г. min × min (табл. 3) наблюдали наследование в F 1 по типу неполного доминирования (h/d = 0,50) и сверхдоминирования (2,00). В 2019 г. (табл. 4) в двух комбинациях скрещиваний установлено неполное доминирование (h/d = 0,50).
Средние значения длины венчика в F 1 за два года по группам скрещиваний составили 8,5 (max × max), 7,7 (max × min), 7,7 (min × max) и 6,7 мм (min × min) при
НСР 05 = 0,2 мм, что также подтверждает существенный генетический контроль признака.
Таблица 4
Наследование в F 1 длины венчика при скрещивании линий подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2019 г.
|
Комбинация скрещивания |
Генотип |
Длина венчика, мм |
НСР 05 |
h/d |
|||
|
Р 1 |
Р 2 |
F 1 |
F 1 по группе |
||||
|
К581 × Л2138 |
8,6 |
8,7 |
8,6 |
0,13 |
-1,00 |
||
|
Л2138 × К581 |
8,7 |
8,6 |
8,6 |
0,12 |
-1,00 |
||
|
ВК905 Б × К581 |
8,5 |
8,6 |
8,2 |
8,5 |
0,10 |
-7,00 |
|
|
К581 × ВК905 Б |
8,6 |
8,5 |
8,5 |
0,14 |
-1,00 |
||
|
Max × max |
ВК905 Б × Л2138 |
8,5 |
8,7 |
8,6 |
0,10 |
-1,00 |
|
|
Л2138 × ВК905 Б |
8,7 |
8,5 |
8,4 |
0,13 |
-2,00 |
||
|
Л2138 × ЛД102 |
8,7 |
6,7 |
8,1 |
0,11 |
0,40 |
||
|
Л2138 × КГ49 |
8,7 |
6,3 |
7,4 |
0,10 |
-0,08 |
||
|
Max × min |
ВК905 Б × ЛД102 |
8,5 |
6,7 |
7,9 |
7,7 |
0,10 |
0,33 |
|
К581 × ЛД102 |
8,6 |
6,7 |
7,6 |
0,10 |
-0,05 |
||
|
К581× КГ49 |
8,6 |
6,3 |
7,3 |
0,12 |
-0,13 |
||
|
ВК905 Б × КГ49 |
8,5 |
6,3 |
7,9 |
0,12 |
0,46 |
||
|
ЛД102 × ВК905 Б |
6,7 |
8,5 |
7,9 |
0,12 |
0,33 |
||
|
ЛД102 × Л2138 |
6,7 |
8,7 |
8,0 |
0,10 |
0,30 |
||
|
Min × max |
ЛД102 × К581 |
6,7 |
8,6 |
7,8 |
7,7 |
0,11 |
0,16 |
|
КГ49 × ВК905 Б |
6,3 |
8,5 |
7,6 |
0,12 |
0,18 |
||
|
КГ49 × Л2138 |
6,3 |
8,7 |
7,5 |
0,11 |
0,00 |
||
|
КГ49 × К581 |
6,3 |
8,6 |
7,5 |
0,10 |
0,04 |
||
|
Min × min |
ЛД102 × КГ49 КГ49 × ЛД102 |
6,7 6,3 |
6,3 6,7 |
6,6 6,6 |
6,6 |
0,09 0,10 |
0,50 0,50 |
Наследование длины венчика в F 2 при скрещивании линий в пределах одной контрастной группы между собой, т.е. max × max (Л2138 × ВК905 Б) и min × min (ЛД102 × КГ49), носило континуальный характер с незначительными двусторонними трансгрессиями. Это говорит об отсутствии факта потенциальной рекомбинации генов с сильным фенотипическим эффектом, контролирующих контрастные проявления признака у линий одной группы. Морфологические отличия по размеру венчика в F 2 этих двух контрастных групп были очевидны (рис. 5).
С другой стороны, наследование длины венчика в F2 при скрещивании линий с контрастными проявлениями признака между собой ВК905 Б × ЛД102, т.е. max × min, также характеризовалось континуальной изменчивостью, однако без трансгрессий (рис. 4). При этом среднее значение F1 (7,9 мм), F2 (7,7 мм) и среднеродительское значение (7,6 мм) были близки, что предполагает наличие адди- тивного действия полигенов. Лимиты распределения значений в популяции F2 – 6,9 и 8,4 мм – соответствовали родителям с минимальной и максимальной величиной признака. Размах варьирования при этом составил 1,5 мм, а CV = 6 %.
Рисунок 4 – Наследование длины венчика трубчатого цветка в скрещивании ВК905 Б × ЛД102 (max × min)
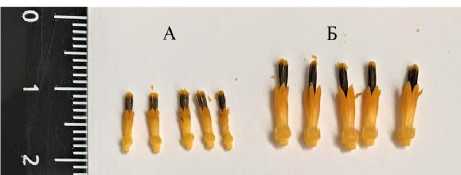
Рисунок 5 – Трубчатые цветки подсолнечника:
А – мелкие (корзинка F 2 ЛД102 × КГ49;
Б – крупные (корзинка F 2 Л2138 × ВК905 Б)
Изученные в 2017 г. линии генетической коллекции подсолнечника показали широкое варьирование диаметра венчика трубчатого цветка от 1,4 у № 424924 до 3,6 мм у КГ19, т.е. с различием в 2,6 раза (рис. 6). Среднее значение при этом составило 2,4 мм, размах варьирования – 2,2 мм со средним коэффициентом вариации CV = 13 %.
Трехлетнее наблюдение в 2017–2019 гг. за четырьмя отобранными линиями с контрастными значениями диаметра венчика КГ19, ВИР130-3 (max) и ВИР391, №
424924 (min) показало, как и в случае с длиной венчика, сильное влияние генотипа на варьирование признака без смены ежегодных фенотипических рангов линий.
-
з,б- з,7 е
3,4-3,5
3,2-3,3
з,о-з,і ее
-
2,s- 2,9 ееееще
в 2,6-2,? ■■■■■■■В в 2,4-2,5 ■■■■■■■■■■■■■■
-
2,2- 2,з eeeeeeeemee
2,о-2,і евееееевев
-
1,8- 1,9 ■■■■
-
1,6- 1,7 •
-
іл-1,5 е
0 5 10 15 20 25 30
Рисунок 6 – Изменчивость диаметра венчика трубчатых цветков у линий генетической коллекции (n = 90)
В среднем за три года диаметр венчика по двум группам линий (max и min) изменялся от 2,9 до 3,4 мм и от 1,4 до 1,7 мм соответственно (табл. 6). Доля влияния фактора генотипа по результатам дисперсионного анализа оценивается в 94 % (табл. 7). При этом влияние условий года выращивания было недостоверным.
Таблица 6
Диаметр венчика трубчатого цветка у линий генетической коллекции
ЦЭБ ВНИИМК, г. Краснодар, 2017–2019 гг.
|
Линия |
Год |
Среднее по годам |
НСР 05 |
|||
|
2017 |
2018 |
2019 |
||||
|
Мax |
КГ19 |
3,6 |
3,3 |
3,3 |
3,4 |
0,1 |
|
ВИР130-3 |
3,0 |
2,9 |
2,8 |
2,9 |
0,1 |
|
|
Мin |
ВИР391 |
1,7 |
1,7 |
1,7 |
1,7 |
0,1 |
|
№ 424924 |
1,4 |
1,7 |
1,6 |
1,6 |
0,1 |
|
|
НСР 05 |
0,2 |
0,1 |
0,1 |
|||
Таблица 7
Результаты двухфакторного дисперсионного анализа диаметра венчика трубчатого цветка у линий генетической коллекции подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2017–2019 гг.
|
Источник вариации |
SS |
df |
MS |
F |
F кри-тиче-ское |
Доля влияния, % |
|
Год |
0,06 |
2 |
0,03 |
1,01 |
3,03 |
- |
|
Генотип |
174,39 |
3 |
58,13 |
2071,69 |
2,64 |
93,8 |
|
Взаимодействие |
3,51 |
6 |
0,59 |
20,84 |
2,13 |
- |
|
Внутри |
8,08 |
288 |
0,03 |
4,3 |
||
|
Итого |
186,04 |
299 |
100 |
Примечание: SS - сумма квадратов;
df – степени свободы; MS – средний квадрат;
F – критерий Фишера
При реципрокном скрещивании линий с максимальными значениями диаметра венчика в 2018 г. max × max (табл. 8) наблюдали наследование в F 1 по полностью промежуточному типу (h/d = 0). В 2019 г. (табл. 9) также установлено промежуточное наследование (h/d = 0,20).
Таблица 8
Наследование в F 1 диаметра венчика при скрещивании линий подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2018 г.
|
Комбинация скрещивания |
Генотип |
Диаметр венчика, мм |
НСР 05 |
h/d |
|||
|
Р 1 |
Р 2 |
F 1 |
F 1 по группе |
||||
|
Max × max |
КГ19 × ВИР130-3 |
3,3 |
2,9 |
3,1 |
3,1 |
0,10 |
0 |
|
ВИР130-3 × КГ19 |
2,9 |
3,3 |
3,1 |
0,14 |
0 |
||
|
ВИР130-3 × № 424924 |
2,9 |
1,7 |
2,6 |
0,07 |
0,50 |
||
|
Max × min |
ВИР130-3 × ВИР391 |
2,9 |
1,7 |
2,4 |
2,4 |
0,07 |
0,16 |
|
КГ19 × ВИР391 |
3,3 |
1,7 |
2,4 |
0,09 |
-0,13 |
||
|
КГ19 × № 424924 |
3,3 |
1,7 |
2,3 |
0,06 |
-0,25 |
||
|
ВИР391 × ВИР130-3 |
1,7 |
2,9 |
2,4 |
0,06 |
0,16 |
||
|
Min × max |
ВИР391 × КГ19 |
1,7 |
3,3 |
2,4 |
2,4 |
0,08 |
-0,13 |
|
№ 424924 × ВИР130-3 |
1,7 |
2,9 |
2,6 |
0,09 |
0,50 |
||
|
№ 424924 × КГ19 |
1,7 |
3,3 |
2,2 |
0,07 |
-0,37 |
||
|
Min × min |
№ 424924 × ВИР391 |
1,7 |
1,7 |
1,8 |
1,9 |
0,04 |
- |
|
ВИР391 × № 424924 |
1,7 |
1,7 |
1,9 |
0,03 |
- |
||
В восьми комбинациях реципрокных скрещиваний линий с максимальным и минимальным значениями диаметра венчика в 2018 г. (табл. 8) наблюдали в шести случаях наследование в F 1 по промежуточному типу (h/d от -0,37 до 0,16), а в двух случаях (ВИР130-3 × № 424924 и № 424924 × ВИР130-3) отмечено неполное доминирование (0,50). В 2019 г. (табл. 9) в четырех комбинациях установлено промежуточное наследование (h/d от -0,25 до -0,06) и в четырех – неполное доминирование (h/d от 0,50 до 0,81).
При реципрокном скрещивании двух линий с минимальными значениями диаметра венчика в 2018 г. min × min (табл. 8) наблюдали наследование в F 1 по типу сверхдоминирования (значение h/d из-за отсутствия различий между линиями не рассчитывали). В 2019 г. (табл. 9) в двух комбинациях скрещиваний также установлено сверхдоминирование (h/d = 1,3).
Средние значения диаметра венчика в F 1 за два года по группам скрещиваний составили 3,1 (max × max), 2,5
(max × min), 2,5 (min × max) и 1,9 мм (min × min) при НСР 05 = 0,2 мм, что дополнительно подтверждает существенный генетический контроль признака.
Таблица 9
Наследование в F 1 диаметра венчика при скрещивании линий подсолнечника
ЦЭБ ВНИИМК, г. Краснодар, 2019 г.
|
Комбинация скрещивания |
Генотип |
Диаметр венчика, мм |
НСР 05 |
h/d |
|||
|
Р 1 |
Р 2 |
F 1 |
F 1 по группе |
||||
|
Max × max |
КГ19 × ВИР130-3 |
3,3 |
2,8 |
3,1 |
3,1 |
0,11 |
0,20 |
|
ВИР130-3 × КГ19 |
2,8 |
3,3 |
3,1 |
0,16 |
0,20 |
||
|
ВИР130-3 × № 424924 |
2,8 |
1,7 |
2,7 |
0,13 |
0,81 |
||
|
Max × min |
ВИР130-3 × ВИР391 |
2,8 |
1,6 |
2,5 |
2,5 |
0,15 |
0,50 |
|
КГ19× ВИР391 |
3,3 |
1,6 |
2,4 |
0,10 |
- 0,13 |
||
|
КГ19× № 424924 |
3,3 |
1,7 |
2,3 |
0,10 |
- 0,25 |
||
|
ВИР391 × ВИР130-3 |
1,6 |
2,8 |
2,5 |
0,15 |
0,50 |
||
|
Min × max |
ВИР391 × КГ19 |
1,6 |
3,3 |
2,4 |
2,5 |
0,10 |
- 0,06 |
|
№ 424924 × ВИР130-3 |
1,7 |
2,8 |
2,7 |
0,12 |
0,81 |
||
|
№ 424924 × КГ19 |
1,7 |
3,3 |
2,4 |
0,10 |
- 0,13 |
||
|
min × min |
№ 424924 × ВИР391 ВИР391 × № 424924 |
1,7 1,6 |
1,6 1,7 |
1,9 1,9 |
1,9 |
0,13 0,14 |
1,30 1,30 |
Наследование диаметра венчика в F 2 при скрещивании линий в пределах одной контрастной группы между собой, т.е. max × max (ВИР 130-3 × КГ19) и min × min (№ 424924 × ВИР391), носило континуальный характер с незначительными двусторонними трансгрессиями. Это говорит, как и в случае с длиной венчика, об отсутствии факта потенциальной рекомбинации генов с сильным фенотипическим эффектом, контролирующих контрастные проявления признака у линий одной группы.
С другой стороны, наследование диаметра венчика в F2 при скрещивании линий с контрастными проявлениями признака между собой ВИР130-3 × ВИР391, т.е. max × min, также характеризовалось континуальной изменчивостью, однако без значительных трансгрессий (рис. 7). При этом среднее значение F1 (2,5 мм), F2 (2,4 мм) и среднеродительская величина (2,2 мм) были близки, что предполагает наличие аддитивного действия полигенов. Лимиты распределения значений в F2 – 1,6 и 3,2 мм – соответствовали родителям с минимальной и максималь- ной величиной признака при размахе варьирования равном 1,6 мм. Эта изменчивость классифицируется как средней степени при CV = 13 %.
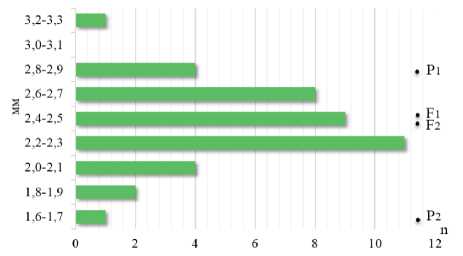
Рисунок 7 – Наследование диаметра венчика трубчатого цветка в скрещивании ВИР130-3 × ВИР391 (max × min)
Выводы . В генетической коллекции подсолнечника ВНИИМК содержатся линии с наследственно контролируемыми различиями по морфологическим признакам трубчатого цветка – длине венчика от 6,3 до 9,2 мм и его диаметру от 1,4 до 3,6 мм. Изменчивость длины венчика оценивается как слабая (CV = 9 %), а диаметра – как средняя (CV = 13 %), что отражает коэволюционное ограничение варьирования, прежде всего, адаптивно ценного признака длины венчика. Это ограничение по максимальной длине венчика, вероятно, необходимо для эффективной работы насекомых-опылителей с фиксированной длиной хоботка, а по минимальной – целесообразностью защиты нектара от нежелательных насекомых или от его чрезмерно быстрого высыхания. При этом между длиной венчика и его диаметром в линиях генетической коллекции не установлено достоверной корреляции (r = 0,12 < r 05 = 0,21).
Наследование длины и диаметра венчика в F1 и F2 соответствовало полиген-ной аддитивной модели. Установлено, что скрещивание между собой линий с максимальными значениями признаков не приводило к увеличению значений фенотипа в потомстве, а скрещивание линий с минимальными параметрами не вызывало дальнейшего снижения величин признаков. Типы наследования в F1 этих энто-мофильно значимых морфологических признаков трубчатого цветка подтверждают тезис о т.н. закрытой изменчивости адаптивно ценных особенностей растений.
Список литературы Наследование длины и диаметра венчика у трубчатых цветков подсолнечника
- Бочковой А.Д., Перетягин Е.А., Хатнянский В.И., Камардин В.А. Роль пчёлоопыления в получении высоких и стабильных урожаев кондитерских сортов подсолнечника (обзор) // Масличные культуры. Науч.-тех. бюл. ВНИИМК. -2017. - Вып. 1 (169). - С. 83-92.
- Mallinger R.E., Bradshaw J., Varenhorst A.J., Prasifka J.R. Native Solitary Bees Provide Economically Significant Pollination Services to Confection Sunflowers (Helianthus annuus L.) (Asterales: Asteraceae) Grown Across the Northern Great Plains // Journal of Economic Entomology. - 2019. -No 1. - P. 40-48.
- Матиенко А.Ф. Влияние цитоплазматической мужской стерильности типа Heliantus petiolaris на некоторые селекционно-ценные признаки растения подсолнечника: дис.. канд. с.-х. наук / Александр Филиппович Матиенко. -Краснодар, 1988. - 162 с.
- Joksimovic J., Atlagic J., Sakac Z., Miklic V., Dusanic N. Mode of inheritance and heritability of disc flower corolla length and nectar content in sunflower // Genetika. -2003. - V. 35. - No 1. - P. 59-65.
- Palmer J.H., Steer V.T. The generative area as the site of floret initiation in the sunflower capitulum and its intergration to predict floret number // Field Crop Research. -1985. - No 11. - P. 1-12.
- Skoric D. Sunflower breeding // Sunflower Genetics and Breeding. International monograph. - Serbian Academy of Science and Arts, Branch in Novi Sad, 2012. - 520 p.
- Mallinger R.E., Prasifka J.R. Bee visitation rates to cultivated sunflowers increase with the amount and accessibility of nectar sugars // Journal of applied entomology. - 2017. - P. 1-13.
- Шаповалова Л.Г. Влияние условий выращивания гибридных семян подсолнечника на их генетическую чистоту, урожайные свойства и посевные качества: дис.. канд. с.-х. наук / Любовь Геннадиевна Шаповалова. - Краснодар, 2003. -113 с.
- Portlas Z.M., Tetlie J.R., Prischmann-Voldseth D., Hulke B.S., Prasifka J.R. Variation in floret size explains differences in wild bee visitation to cultivated sunflowers // Plant Genetic Resources. - 2018. - Vol. 16. -Is. 6. - P. 498-503.
- Joksimovic J., Atlagic J., Sakac Z., Miklic V., Dusanic N. Phenotypic and genotypic variability of disc flower corolla length and nectar content in sunflower // Genetika. - 2003. - V. 35. - No 2. - P. 131-138.
