Наследование признака "число зёрен в колосе" у яровой мягкой пшеницы в зависимости от условий среды экологических пунктов программы "Экада"

Автор: Менибаев Асхат Исмаилович, Мальчиков Птр Николаевич, Зуева Анастасия Александровна, Захаров Владимир Григорьевич, Кривобочек Виталий Григорьевич, Василова Нурания Зуфаровна, Багавиева Эльмира Зинуровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-3 т.20, 2018 года.
Бесплатный доступ
Число зерен в колосе один из самых важных компонентов урожайности. В многочисленных исследованиях результатов селекционного улучшения пшеницы, установлена высокая значимость вклада в этот процесс озернённости колоса и колоска. В связи с этим целенаправленная селекция по этому признаку может быть эффективным методом ускорения селекции на высокую урожайность. Научное обоснование этого процесса включает поиск, изучение, подбор исходного материала, определение наследования и выработку стратегии отбора. Многолетнее функционирование кооперативной программы по селекции яровой мягкой пшеницы «Экада», позволило отобрать генотипы с высокой и стабильной урожайностью. Семь сортов (Архат, Омская 36, Экада 113, Экада 148, Эстивум 1079, Эстивум 1311ае72, Экада 204) в том числе созданные в «Экаде», были включены в программу изучения наследования элементов урожайности. Изучение проведено по методике B.I.Hayman с применением полудиаллельной схемы и изучения в 4-х пунктах: Казань, Безенчук, Пенза, Ульяновск. В результате было установлено: 1) в Ульяновске все генетические параметры были недостоверны, что осложняет работу по отбору в этом пункте; 2) в Казани значимыми были только аддитивные эффекты, что создает благоприятные условия для отбора; 3) в Безенчуке значимыми были аддитивные эффекты и один параметр доминирования; 4)в Пензе все генетические параметры были достоверны; 5) отмечены изменения конфигурации графиков регрессии Wr на Vr, смена доминантного статуса сортов и переопределение генетической формулы признака. В связи с этим предложено проводить отборы по числу зерен в колосе в старших поколениях (F4 - F5 ). Для уточнения понимания зависимости генетических параметров от лимитирующих факторов среды, формируемых экологическими пунктами необходимо продолжить изучение признака в различных средах.
Наследование, признак, зерно, колос, яровая мягкая пшеница
Короткий адрес: https://sciup.org/148312397
IDR: 148312397 | УДК: 633.111.1
Inheritance of the trait “number of kernels in the spike” at the spring soft wheat depend on the conditions of environmental ecological points of the “Ecada” program
The number of grains in the spike one of the most important components of yield. In numerous studies of the results of the breeding improvement of wheat, the high importance of the contribution to the process of the lachrymation of the spike and spikelet has been established. In this regard, targeted selection for this feature can be an effective method of accelerating selection for high yields. The scientific rationale for this process includes the search, study, selection of source material, determination of inheritance and development of a selection strategy. The long-term functioning of the cooperative program for the selection of spring soft wheat “Ekada” made it possible to select genotypes with high and stable yields. Seven cultivars (Arhat, Omskaya 36, Ekada 113, Ekada 148, Estivum 1079, Estivum 1311ae72, Ekada 204), including those created in the “Ekada program”, were included in the program for studying the inheritance of the elements of yield. The study was carried out according to the method of B.I.Hayman with the use of a semi-dialect scheme and study in 4 points: Kazan, Bezenchuk, Penza, Ulyanovsk. As a result, it was established: 1) in Ulyanovsk, all genetic parameters were unreliable, which complicates the selection work at this point; 2) in Kazan only additive effects were significant, which creates favorable conditions for selection; 3) in Bezenchuk, additive effects and one dominance parameter were significant; 4) in Penza all genetic parameters were reliable; 5) changes in the configuration of regression graphs Wr on Vr, change of the dominant status of cultivars and redefinition of the genetic formula of the trait are noted. In connection with this, it has been proposed to conduct selections for the number of grains in the spike in the older generations (F4-F5). To clarify the understanding of the dependence of genetic parameters on the limiting factors of the environment formed by environmental points, it is necessary to continue studying the trait in various environments.
Текст научной статьи Наследование признака "число зёрен в колосе" у яровой мягкой пшеницы в зависимости от условий среды экологических пунктов программы "Экада"
ствующих генов (генетических систем), условий выращивания растений и взаимодействием «генотип-среда». Успех селекции (отбора) по этим признакам зависит от соотношения эффектов воздействия на них каждой компоненты из этой триады. Идеальной считается ситуация, где основной вклад в дисперсию признака вносит генетическая система, контролирующая признак. В этом случае отбор по фенотипу оказывается результативным, его эффект сохраняется в последующих поколениях независимо от условий среды. Превалирование в дисперсии признака генотип-средовых взаимодействий, ограничивает эффективность использования результатов отбора параметрами среды, в которой он был проведён. Определяющая роль среды при формировании количественного признака, создаёт неблагоприятный фон для отбора и делает его бессмысленным. Применение двухфакторного дисперсионного анализа (ANOVA) позволят общую сумму квадратов варьирования урожайности в целом по эксперименту аддитивно распределить на эффекты среды, генотипов и их взаимодействие. При этом структура генотипи- ческой и генотип-средовой вариансы не анализируется. Доля генотипической вариансы в общем варьировании признака (фенотипическая варианса), представляет собой коэффициент наследуемости в широком смысле слова H2=σ2g/ σ2ph, который включает в себя аддитивные, доминантные, сверхдоминантные и эпистатиче-ские эффекты. Доминирование и эпистаз создают неблагоприятный фон для отбора, особенно в ранних поколениях. Репродукция (пересев), отобранных фенотипов, «снимает» эффекты доминирования и эпистаза, что может свести к нулю величину селекционного дифференциала, зафиксированного в момент отбора. Наиболее результативен отбор в популяциях с превалированием аддитивной вариансы или с высокими значениями коэффициента наследуемости в узком смысле слова- h2=σ2a/σ2ph. Диаллельный анализ даёт возможность по генетическим параметрам и графику определить тип действия генов [1] и спрогнозировать эффективность отбора в конкретной среде.
Признак «число зерен в колосе» наряду с плотностью продуктивного стеблестоя и массой зерновки, определяет величину урожайности зерна с единицы посевной площади. Сильная изменчивость признака обусловлена его чувствительностью к воздействию условий среды в периоды его формирования в онтогенезе растений. Тем не менее, это один из основных селекционных признаков, роль которого в процессе улучшения пшеницы, на отдельных этапах селекции была определяющей. Актуальность исследования связана с поиском наиболее благоприятных условий среды для максимального проявления генетических систем озернённости колоса, функционирующих в исследуемой сортовой популяции.
В связи с этим цель исследований, результаты которых представлены в настоящей публикации, заключалась в оценке значимости генетических параметров и вклада эффектов генов в формирование признака «число зерен в колосе» в зависимости от условий среды, формируемых экологическими пунктами кооперативной программы по селекции яровой мягкой пшеницы «Экада».
МАТЕРИАЛЫИ МЕТОДЫ ИССЛЕДОВАНИЯ
Для реализации поставленной задачи были подобраны следующие сорта: Архат, Эстивум 1079, Эстивум 1311ае72, Омская 36, Экада148, Экада113, Экада 204. Ранги сортов по исследуемому признаку меняются в зависимости от условий среды в пунктах изучения. Это обстоятельство позволяет получить необходимую информацию по генетике признака «число зерен в колосе» с применением диаллельных скрещиваний. Для полного представления о сортах, включённых в диаллельный анализ приводим их краткую характеристику по основным хозяйственно-ценным признакам.
Архат . Сорт получен путём скрещивания Ишеевская/Л-503. Зерно крупное, - масса 1000 зёрен 25,6-45,2 г. Содержание белка в зерне 13,8–17,0%, клейковины в зерне 30,0–35,6% первой и второй группы качества. Архат имеет муку жёлтой окраски, которая определяется геном Y, сцепленным с геном устойчивости к бурой ржавчине Lr 19, что в сочетании с геном Lr 34, унаследованным от сорта Ишеевская, обеспечивает устойчивость к поражению бурой ржавчиной.
Омская 36 Происхождение - Лютесценс 150/86–10/ Runar (Норвегия). Масса 1000 зёрен 39–46г. Среднеранний, вегетационный период 87 суток. Устойчивость к засухе высокая, на инфекционном фоне практически устойчив к пыльной головне, устойчивость к бурой ржавчине проявляется по типу медленного «ржавления».
Экада 113 . Получен от скрещивания сорта Скала БР (аналог сорта Скала - аббревиатура БР означает устойчивость к бурой ржавчине) с сортом Юлия. Сорт устойчив к бурой ржавчине (устойчивость к бурой ржавчине детерминируется геном LrTR). Масса 1000 зерен 32-39 г. Сорт степного экотипа, среднеспелый. Вегетационный период 87-90дней. Среднестебельный (66–95 см), среднеустойчивый к полеганию. Обладает комплексной устойчивостью к бурой, стеблевой, жёлтой ржавчине и мучнистой росе. Высокозасухоустойчив.
Экада 148 . Получен в результате скрещивания Экада 6 / Прохоровка. Несёт в себе пирамиду генов Lr19 и Lr26. Обладает свойствами высокой общей сортообразующей способностью, унаследованной от Саратовской 46 и Прохоровки, имеет компенсирующий комплекс генов по качеству, который сформировался на фоне отрицательных эффектов ржаной транслокации (1B/1R) в сегменте хромосомы, включающей Lr 26.
Эстивум 1079. Сорт выведен методом индивидуального отбора из гибрида F3 1438ae45 Дуэт / Юго-Восточная-2. Родительские формы были подобраны, исходя из их высокой продуктивности и пластичности, высокой засухо-жаростойкости, а также устойчивости к листовой бурой ржавчине и полеганию. Несёт ген Lr9. Предполагается что у сорта общая сортообразующая способность и компенсаторный комплекс генов, влияющий на качество, унаследованы от сорта Чайка.
Эстивум 1311ае72. Сорт получен в результате скрещивания Эритроспермум 11/Тулайков-ская 100. Сорт несёт пырейную хромосому 6Agi полностью заменившую 6E пшеничную хромосому. Имеет крупное зерно. Поскольку 6Agi хромосома включает генетическую систему с отрицательным эффектом на массу 1000 зёрен, правомерно предположить наличие у этого генотипа компенсаторного комплекса генов увеличивающих массу зерновки.
Экада 204 получена в результате скрещивания СП188-14 / Любава 5 Предполагается что у сорта общая сортообразующая способность и компенсаторный комплекс генов, действующий на качество, унаследованы от сорта Любавы5.
Исследования проведены в 4 точках программы «Экада»: Безенчуке, Пензе, Ульяновске и Казани. Во всех точках была единая методика исследований. Гибридные зерна получали в каждом учреждении по полудиаллельной схеме. Посев выполнен рендомизированными блоками в трехкратной повторности по 20 зёрен, однорядковыми делянками по 1 метру с междурядьми 20 сантиметров. Фенологические наблюдения отмечались по фазам развития в кущение, колошение и созревание.
Уборка проводилась в момент полной спелости путём выдёргивания растения с корнями. Снопы этикировали и хранили в снопохрани-лище 2 недели. В анализ брали 10 растений с каждой повторности без отбора, исключались больные растения. Полученные значения признака были изучены при помощи дисперсионного анализа и методом диаллельного анализа по B.I.Hayman [2] с применением персонального компьютера и специализированной программы «Аgros», разработанной доктором биологических наук С.П.Мартыновым.
Варьирование условий среды в экопунктах повлияло на формирование и наследование признака, в связи с этим ниже приведена краткая информация о состоянии гидротермического и фитопатогенного режимов в экопунктах эксперимента в период вегетации растений.
В Казани условия среды сильно отличались от среднемноголетних характеристик повышенными температурами и низким уровнем влаго-обеспеченности. Засуха негативно повлияла на фертильность колосков и общую озернённость колоса. Несмотря на засуху отмечено значительное развитие листовых и головневых болезней. На посевах пшеницы в первой половине вегетации наблюдалось развитие мучнистой росы, в период формирования и налива зерна имела место эпифитотия бурой и стеблевой ржавчины. Комплексное влияние этих факторов привело к формированию значительных сортовых различий по признаку «число зёрен в колосе». Признак варьировал от 38 зерен у сорта Омская 36 до 47-46 зерен у Экады 204 и Эстивум 1311ае72 соответственно.
В Безенчуке в целом за вегетационный период сложились очень засушливые условия (ГТК=0,52). Также, как и в Казани имела место эпифитотия бурой и стеблевой ржавчине. На- блюдалась дифференциация по признаку «число зёрен в колосе» от 19 у сорта Экада 113 до 43 зёрен у сорта Экада 204.
В Пензе на фоне повышенных температур выпало достаточное количество осадков. Гидротермический коэффициент варьировал в период вегетации от 0,81 до 1,1. В этих условиях дифференциация сортов по признаку «число зёрен в колосе» была вполне приемлемой. Варьирование признака отмечено в пределах от 34 зёрна у Эстивум 1079 до 46 у Эстивум 1311ае72 .
В Ульяновске наблюдалось сильное отрицательное воздействие негативных условий среды на элементы продуктивности. Кроме недостаточного увлажнения, посевы были поражены мучнистой росой в начале вегетации, пиренофорозом, бурой и стеблевой ржавчинами в колошение и в период налива зерна. Число зерен в колосе по сортам варьировало от 38 до 52 штук соответственно у Омской 36 и Экады 204.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
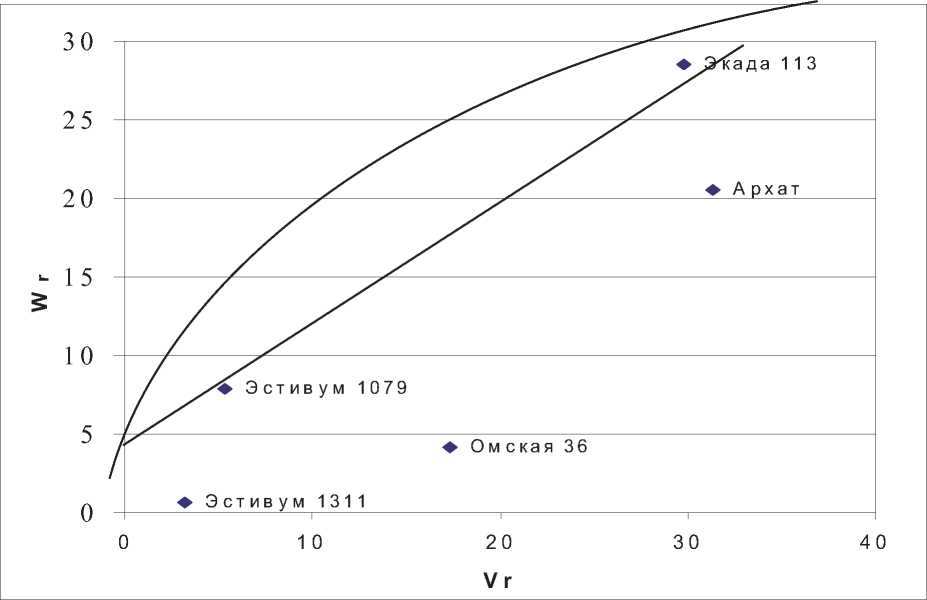
В среднем по всем пунктам наибольшее число зерен в колосе отмечено у Экады 204 (45,9 шт.), Экады 148 (42,2шт.) и Архата (41,4шт.). Наименьшая величина признака была у сортов Омская 36 (33,7 шт.), Экада 113 (33,8шт.) и Эсти-вум 1079 (35,0 шт.). При этом у сорта Архат варьирование признака по средам было одним из самых незначительных – второе место среди исследуемых сортов в ряду, при ранжировании по возрастанию коэффициента вариации (CV e = 10,7%). Эти обстоятельства позволили вполне обоснованно применить к массиву данных методику диаллельного анализа и получить генетико-статистические параметры исследуемого признака (табл. 2; рис.1-3).
К известным ограничениям метода B.I.Hayman (гомозиготность родителей, отсутствие множественного аллелизма, эпистаза, реципрокных эффектов, независимость распределения генов у родителей, диплоидное расщепление), В.А.Драгавцев и др. (1984) добавили ещё три: равная доля доминантных и рецессивных генов, направленное доминирование, одинаковый вклад генов в признак.
При этом авторы рекомендовали использовать следующие относительно бесспорные параметры: (H1 / D)1/2 – средняя степень доминирования в локусе, H2 / 4 H1 – среднее значение произведения частот доминантных и рецессив-
Таблица 1. Число зёрен в колосе сортов и гибридов F1 по экологическим пунктам, 2016 год
|
Сорт |
Казань |
Безенчук |
Пенза |
Ульяновск |
||||
|
Число зерен в колосе у родител. сортов |
Число зерен в к-се гибридов F1 |
Число зерен в колосе у родител. сортов |
Число зерен в к-се гибридов F1 |
Число зерен в колосе у родител. сортов |
Число зерен в к-се гибри дов F1 |
Число зерен в колосе у родител. сортов |
Число зерен в к-се гибридов F1 |
|
|
Эстивум 1311 |
48,5 |
47,6 |
33,8 |
34,5 |
45,9 |
38,1 |
47,9 |
49,4 |
|
Эстивум 1079 |
41,7 |
43,2 |
23,8 |
23,9 |
34,2 |
38,3 |
40,3 |
42,5 |
|
Экада148 |
45,5 |
- |
35,8 |
25,9 |
42,8 |
37,6 |
44,5 |
44,4 |
|
Омская36 |
38,5 |
40,1 |
23,7 |
22,7 |
34,9 |
37,4 |
37,6 |
40,1 |
|
Архат |
47,5 |
42,1 |
28,2 |
31,8 |
41,3 |
39 |
48,6 |
46,3 |
|
Экада204 |
45,9 |
- |
43,4 |
25,2 |
43,9 |
41,9 |
50,3 |
48,2 |
|
Экада 113 |
35,9 |
43,5 |
19,5 |
29 |
36,3 |
40,2 |
43,5 |
44,6 |
|
НСР 0,05 |
7,3 |
7,9 |
6,1 |
8,9 |
||||
Таблица 2. Компоненты генетической дисперсии в зависимости от условий среды в пунктах изучения, 2016 год
|
Компонента |
Казань |
Безенчук |
Пенза |
Ульяновск |
|
D |
21.8423* |
98.9197* |
21.7737* |
6.8049 |
|
F |
1.3667 |
157.2635 |
33.7432* |
-40.6523 |
|
H1 |
30.7004 |
237.0639* |
48.1010* |
8.7699 |
|
H2 |
21.6225 |
137.1356 |
31.4880* |
7.5639 |
|
h |
-10.3683 |
-21.8637 |
2.2923 |
-14.3200 |
|
E |
16.3217* |
54.2956* |
2.7795* |
24.0151* |
|
Эстивум 1311 ае 72 |
44.3989 |
225.5911 |
12.6019 |
-5.3323 |
|
Эстивум 1079 |
40.2651 |
- |
43.0559* |
-78.5016* |
|
Экада 148 |
- |
372.0055* |
16.6151 |
- |
|
Омская 36 |
29.9059 |
293.2096 |
50.7481* |
-24.7255 |
|
Архат |
-50.6003 |
126.4120 |
36.3037* |
-38.7166 |
|
Экада 204 |
- |
-294.8004* |
- |
-8.6659 |
|
Экада 113 |
-57.1363 |
221,1613 |
43.1347* |
-87.9720 |
|
V (H1/ D) |
1,185558 |
1,5480725 |
1,4863152 |
1,135236 |
|
1/4 H2/ H1 |
0,1 |
0,14 |
0,163655 |
0,215621 |
|
V(D H1)+ F/V(D H1)- F |
1,11 |
-75,18 |
0,02 |
-0,6744 |
|
r |
-0.353 |
0.738 |
-0.528 |
0.902 |
ных генов в локусе, ((4DH1)1/2 + F1) / ((4DH1)1/2 - F1) -отношение общего числа доминантных к общему числу рецессивных генов у всех родителей, R [xi; (Wr +Vr)] – корреляция между средними значениями родителей и суммой коварианс и вари-анс или мера направленности доминирования. Эти параметры были использованы при анализе полученных данных по признаку «число зерен в колосе» по, представленным выше экопунктам.
Отчетливо проявилось влияние условий пунктов на структуру генетической вариансы. В Ульяновске все генетические параметры оказались недостоверными. Значимые различия между генотипами в Ульяновске оказываются бесполезными при прогнозировании эф- фективности отбора, поскольку селекционный дифференциал (разница в величине признака между родительскими сортами) в этих условиях будет плохо наследоваться и эффективность отбора может оказаться низкой. В Казани, Безен-чуке и Пензе отмечены значимые аддитивные эффекты, что предполагает положительный отклик на отбор в потомстве. Средняя степень доминирования по признаку в Безенчуке и Пензе составила 1,55 и 1,49 соответственно. В Казани параметр доминирования Н1 был недостоверен, что при значимых аддитивных эффектах создает благоприятные условия для отбора в ранних расщепляющихся поколениях. В Безенчуке у родительских сортов превалируют рецессив-
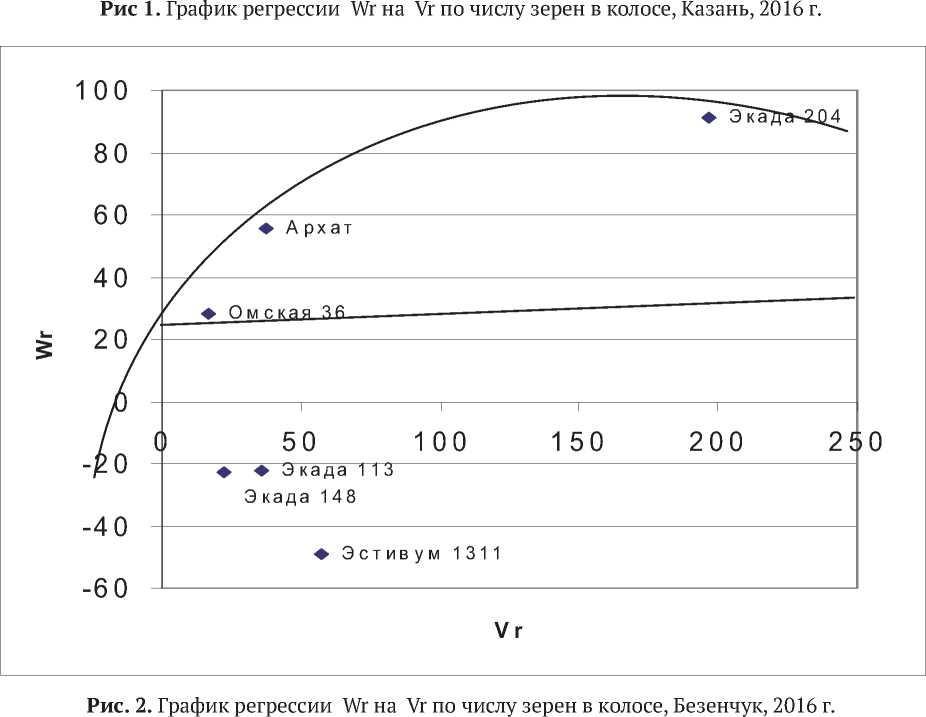
ные гены ((4DH1)1/2 + F1) / ((4DH1)1/2 - F1) = -75.2. Доминирование носит направленный характер (R= 0.74), то есть рецессивные гены увеличивают признак. Линия регрессии отклоняется от линии единичного наклона в сторону оси Vr, пере- секая ось Wr выше начала координат, указывая на взаимодействие генов по типу комплементарного эпистаза, что предполагает очень благоприятную ситуацию для отбора трансгрессий на основе рецессивных генов в ранних расщепляю-
Vr
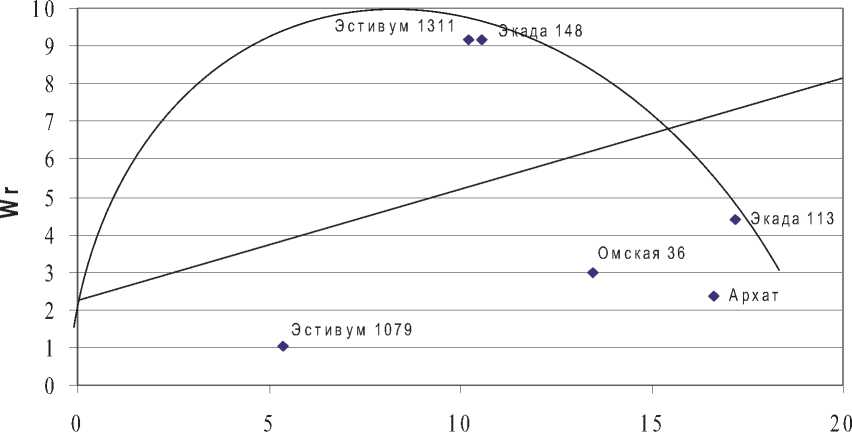
Рис. 3. График регрессии Wr на Vr по числу зерен в колосе, Пенза, 2016 г.
щихся поколениях. Максимальное количество доминантных генов, если судить по расположению точек на графике – около 90,0%, содержат сорта Омская 36, Экада 113 и Экада 148. Архат и Эстивум 1311 имеют 70,0% доминантных генов. Генетическая система исследуемого признака у сорта Экада 204 включала около 80,0 % рецессивных генов с положительными эффектами на формирование признака.
Поскольку в Казани эффекты доминирования были недостоверны, коэффициент корреляции R [xi; (Wr +Vr)] был отрицательным, но по абсолютной величине незначительным и незначимым, правомерно предположить, что в этих условиях функционирует две системы рецессивных и доминантных генов с положительными эффектами на величину признака.
В Пензе параметры доминирования, также как аддитивные эффекты, оказывали значимое влияние на формирование признака. Соотношение доминантных и рецессивных генов в исследуемой сортовой популяции было одинаковым, коэффициент корреляции R [xi; (Wr +Vr)] достиг средней абсолютной величины с отрицательным знаком, что предполагает положительное действие доминантных генов в направлении отбора. Линия регрессии Wr на Vr пересекает ось коварианс выше начала координат, что указывает на неполное доминирование в локусах. Сорта Экада 113, Омская 36, Архат и Эстивум 1079 имели значимые величины Fr линий. Первые три сорта расположены на графике регрессии Wr на Vr в зоне превалирования рецессивных генов, последний в зоне доминирования. Эстивум 1311 и Экада 148 имели недостоверные значения Fr линий, что предполагает баланс доминантных и рецессивных генов.
Учитывая, что сорта Экада 113 и Омская 36 в условиях Безенчука содержали максимальное количество доминантных аллелей с отрицательными эффектами и их превалирование наблюдалось также в сорте Архат, очевидно, что в наследовании признака в Безенчуке и Пензе имеет место переопределение генетической формулы признака. Этот вывод подтверждается общим изменением конфигурации графиков в зависимости от условий среды в экопунктах. При этом наиболее благоприятные генетические предпосылки для отбора сложились в Безенчуке и Пензе. Учитывая значительную динамику точек на графике, отражающих свойства сортов, изменение генетической формулы признака с включением как доминантных, так и рецессивных аллелей, эпистатические эффекты, отборы в расщепляющихся популяциях по числу зерен в колосе, целесообразнее проводить в старших поколениях (F4 - F5). Необходимо продолжить изучение признака в различных средах для уточнения понимания зависимости генетических параметров от лимитирующих факторов среды, формируемых экологическими пунктами.
Список литературы Наследование признака "число зёрен в колосе" у яровой мягкой пшеницы в зависимости от условий среды экологических пунктов программы "Экада"
- Драгавцев В.А., Цильке Р.А., Рейтер Б.Г. и др. Генетика признаков продуктивности яровых пшениц в Западной Сибири. Новосибирск: Наука, 1984. 230 с.
- Hayman B.I. The theory and analis of diallel crosses. I.//Genetics, 1954. Vol.39. N 4. Рр. 789-809.
