Необычный порядок расположения хромосом в поздней профазе - ранней прометафазе митоза у гаплоидного растения кукурузы, полученного с использованием мутации IG

Автор: Юдина Римма Сергеевна, Левитес Евгений Владимирович
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 1 т.7, 2021 года.
Бесплатный доступ
Несмотря на огромное количество работ, посвященных изучению митотического деления, все еще остается много неясного в его механизмах. Например, недостаточно внимания уделено процессам деления клеток у форм растений разного уровня плоидности. В литературе имеются лишь фрагментарные данные о гаплоидах у низших растений. Это не позволяет делать какие-либо обобщения относительно митоза у гаплоидов высших растений. В данной статье приведены результаты цитологического исследования митотически делящихся клеток гаплоидных растений кукурузы. В статье продемонстрирована эффективность известного метода Чейза, основанного на применении генетических маркерных признаков и мутации ig (indeterminate gametophyte) для получения и выявления гаплоидов. Описана эффективная модификация простого метода ацетокарминового окрашивания цитологических препаратов. Существенным полученным результатом является, на наш взгляд, обнаружение очень краткого момента в состоянии хромосом в делящейся клетке гаплоидного растения кукурузы. Этот момент характеризуется тем, что хроматиды уже разделились и превратились в независимые хромосомы, но еще не начали своего движения под действием кинетохорных микротрубочек. Именно эта особенность позволила обозначить данное состояние как поздняя профаза - ранняя прометафаза митоза. Не менее важной особенностью обнаруженного момента является необычное упорядоченное расположение хромосом, которые лежат параллельно друг другу вплотную по всей своей длине с расположением центромер на одной линии, которую можно рассматривать как экватор веретена деления. Выявленный факт позволяет предположить, что существенную роль в формировании такого расположения хромосом играет связь хромосом с экватором ядерной мембраны и их последующая связь с экватором веретена деления.
Митоз, гаплоиды кукурузы, упорядоченное расположение хромосом
Короткий адрес: https://sciup.org/14117915
IDR: 14117915 | УДК: 576.351 | DOI: 10.33619/2414-2948/62/01
Unusual order of chromosome arrangement location in late prophase - early prometaphase of mitosis in haploid maize plant obtained with the use of mutation IG
Despite the huge amount of works devoted to the study of mitotic division, there is still a lot of unclear in its mechanisms. For example, insufficient attention has been paid to the processes of cell division in plant forms of different ploidy levels. The literature contains only fragmentary data on haploids in lower plants. This does not allow making any generalizations regarding mitosis in haploids of higher plants. This article presents the results of a cytological study of mitotically dividing cells of haploid maize plants. The article demonstrates the effectiveness of the well-known Chase method, based on the use of genetic markers and the ig mutation (indeterminate gametophyte) for obtaining and detecting haploids. An effective modification of a simple method of acetocarmine staining of cytological preparations is described. An essential result obtained is, in our opinion, the detection of a very brief moment in the state of chromosomes in a dividing cell of a haploid maize plant. This moment is characterized by the fact that the chromatids have already separated and turned into independent chromosomes, but have not yet begun their movement under the action of kinetochore microtubules. It is this feature that made it possible to designate this state as late prophase - early prometaphase of mitosis. An equally important feature of the detected moment is the unusual ordered arrangement of chromosomes, which lie parallel to each other close to each other along their entire length with the centromeres located on one line, which can be considered the equator of the fission spindle. The revealed fact allows us to assume that an essential role in the formation of such an arrangement of chromosomes is played by the bond of chromosomes with the equator of the nuclear membrane and their subsequent connection with the equator of the fission spindle.
Текст научной статьи Необычный порядок расположения хромосом в поздней профазе - ранней прометафазе митоза у гаплоидного растения кукурузы, полученного с использованием мутации IG
Бюллетень науки и практики / Bulletin of Science and Practice
УДК 576.351 + 631.523 + 633.15
В настоящее время накоплено достаточно много данных о механизмах, обеспечивающих прохождение различных фаз митоза [1]. Продолжительность той или иной стадии митоза варьирует в зависимости от вида и физиологического состояния организма, от типа ткани и влияния внешних факторов. Открытие новых данных об особенностях перехода митоза из одной стадии в другую приводит к необходимости применения дополнительных названий («поздний» или «ранний») для обозначения промежуточных этапов митотического деления [1]. Значительные успехи в исследовании митоза обусловлены комплексным подходом, сочетающим в себе молекулярный и микроскопический методы анализа. Эффективным путем исследования митоза является использование мутантов, нарушающих ход отдельных этапов этого сложного процесса [2–3].
Примечательно, что в исследованиях факторов, влияющих на ход митоза, не развит такой аспект как изучение влияния уровня плоидности. На сегодняшний день крайне мало сведений об особенностях прохождения митоза у гаплоидных растений. Есть, например, первые работы по характеристике митоза у гаплоидных и диплоидных водорослей Rhizoclonium hieroglyphicum [4–5]. В них представлено общее описание стадий митоза у этого вида, дано число и размеры хромосом, и сопоставлены характеристики кариотипов у представителей рода Rhizoclonium . Эти знания полезны, однако для большей полноты картины необходимы сведения о ходе митоза у гаплоидов высших растений.
На возможное влияние дозы генома на расположение хромосом в делящихся клетках указывают результаты, полученные на арабидопсисе: в ядрах диплоидных клеток центромеры хромосом занимают периферийное положение, а в клетках с эндоредуплицированными хромосомами расположение центромер иное; оно зависит от типа ткани: в эпидермальных клетках корня сестринские центромеры кластеризуются, а в эпидермальных клетках листа они диспергируют [6].
В связи с вышеизложенным цель данной статьи — привести данные о митозе у гаплоидных растений кукурузы.
Материал и методы исследования
В исследование были взяты гаплоидные растения кукурузы, полученные по методу Чейза [7–8] путем скрещивания растений тестерной линии кукурузы Т с инбредной линией кукурузы W155. Тестерная линия Т была гомозиготна по доминантным генам A, C, R-nj, обуславливающим пурпурную окраску алейронового слоя эндосперма, щитка и зародыша [7– 8], а инбредная линия W155 не имела окраски. При нормальном процессе оплодотворения в гибридных зерновках проявлялась окраска эндосперма, щитка и зародыша. Однако особенность данной тестерной линии заключалась также в том, что она несла в гомозиготном состоянии рецессивный ген ig (indeterminate gametophyte), приводящий к различным нарушениям в развитии зерновки. К таким нарушениям относится гибель либо яйцеклетки, либо привнесенного спермия. Именно это позволяло выявлять гаплоиды. Гаплоиды выявляли в тех зерновках, у которых был окрашен эндосперм, но отсутствовала окраска зародыша. Окрашенный эндосперм указывал на то, что опыление произошло нормально, а отсутствие окраски зародыша указывало на то, что зародыш несет в себе геном только одного родителя. Скрещивание ♀Т х ♂ W155 позволяло выявлять патроклинные (андрогенные) гаплоиды, развившиеся из спермия, полученного от линии W155.
Отобранные зерновки с неокрашенным зародышем высевали в маленькие стаканчики, а затем на стадии четырех листьев пересаживали в грунт. Цитологический контроль проводили на стадии проростков, анализируя точки роста корешков, а также на стадии 3-месячных растений, анализируя точки роста воздушных корней. Взятые образцы тканей фиксировали в смеси спирт: ледяная уксусная кислота (3:1) [9]. Окрашивание цитологических препаратов проводили, помещая образец анализируемой ткани на предметное стекло в каплю раствора Кармина [9]. Все накрывали покровным стеклом почти без надавливания, чтобы не нарушить структуру делящихся клеток, и просматривали под микроскопом.
Результаты и обсуждение
Использование по методу Чейза окраски алейрона и зародыша в качестве маркерных признаков очень эффективно. На Рисунке1 показаны зерновки кукурузы исходных форм, а также зерновка с предполагаемым гаплоидом. Частота выявляемых гаплоидов составляла порядка 1:300, что соответствует частоте, выявленной в работе Камайкла с использованием тестерной формы такого же генотипа [8]. В нашей работе обнаружены зерновки с близнецовыми зародышами (Рисунок 1). Это также совпадает с результатами, полученными ранее в работе Камайкла [8].
Цитологический анализ показал, что в клетках растений, полученных от скрещивания ♀Т х ♂ W155 и отобранных по фенотипическим признакам зерновки, содержится по 10 хромосом (Рисунки 2–3). Эти гаплоиды являются патроклинными, поскольку их хромосомы получены от отцовского растения линии W155.
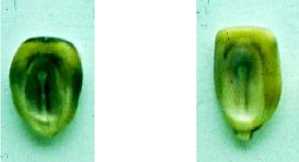



Рисунок 1. Получение патроклинных гаплоидов кукурузы. 1— линия тестера Т; 2 — гибрид ♀Т × ♂ W155; 3 — линия W155; 4 — предполагаемый патроклинный гаплоид; 5 — гибридное зерно с двумя зародышами.
В корешках проростков у гаплоидов встречается миксоплоидия — наличие в ткани наряду с гаплоидными клетками примеси диплоидных. Фотография этих клеток на стадии метафазных пластинок представлена на Рисунке 2.
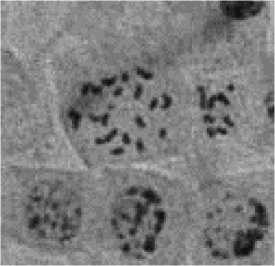
Рисунок 2. Миксоплоидия в корешке гаплоидного растения кукурузы — наличие диплоидной и гаплоидной клеток.
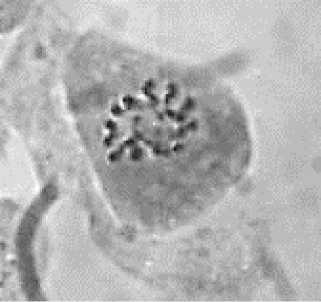
Обращает на себя внимание расположение хромосом в митотических клетках. Можно видеть, что конденсирующиеся хромосомы, выявляемые в корешках проростков, располагаются преимущественно по периферии ядра (Рисунок 3). Это хорошо согласуется с известными данными о расположении хроматина на периферии ядра [10–12]. Гаплоидные клетки, изображенные на этом рисунке, находятся в стадии профазы и различаются по степени конденсации хроматина.
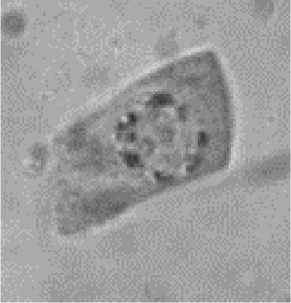
а
б
в
Рисунок 3. Конденсирующиеся хромосомы в профазе митоза в корешках гаплоидных проростков кукурузы: начальная (а), средняя (б) и поздняя (в) стадии конденсации.
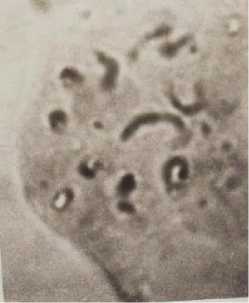
Интересное расположение хромосом было обнаружено при анализе митотического деления в делящихся клетках воздушных корней гаплоидного растения кукурузы. Препарат был получен без применения надавливания на покровное стекло, что позволило в значительной степени сохранить нативное расположение хромосом в делящейся клетке (Рисунок 4). Хромосомы располагаются тесно бок о бок параллельно друг другу, колинеарно, образуя как бы ленту, на средней линии которой располагаются центромеры (Рисунок 4). Несмотря на очень щадящие условия приготовления препарата, лента из хромосом распалась на отдельные фрагменты, однако в каждом фрагменте наблюдается одна и та же закономерность, которую можно интерпретировать следующим образом: центромеры хромосом располагаются по экватору ядра (возможно веретена деления), а сами хромосомы ориентированы по меридианам ядра. Анализируя представленную фотографию препарата можно отметить также следующее.
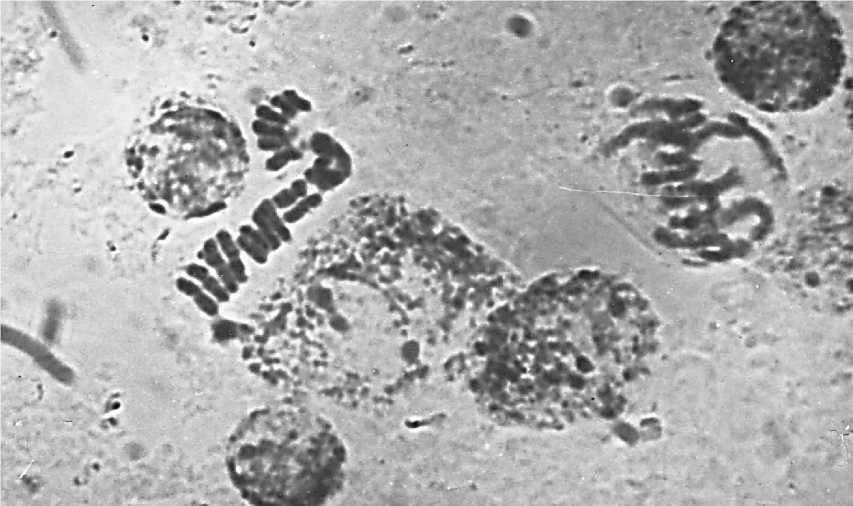
Рисунок 4. Расположение хромосом в поздней профазе — ранней прометафазе митоза в точке роста воздушного корня гаплоидного растения кукурузы.
-
1. Данная стадия может быть обозначена как поздняя профаза — ранняя прометафаза.
-
2. На этой стадии в каждой хромосоме гаплоида хроматиды не только сформировались, но и отделились друг от друга, превратившись в индивидуальные хромосомы, лежащие бок о бок и не начавшие свое движение под действием микротрубочек веретена деления.
-
3. Расположение хромосом на данной стадии можно рассматривать как следствие их расположения по экватору ядра.
Известно, что расположение хромосом в различные периоды клеточного цикла определяется положением центромер и многочисленными связями хроматина с ядерной мембраной [1–2]. Выявленное в корешках гаплоидов расположение хромосом по периферии ядра достаточно хорошо согласуется с известными литературными данными о том, что в интерфазе центромеры хромосом не только располагаются вблизи ядерной мембраны [6], но и непосредственно связаны с ней [2]. Этот контакт очень важен для нормального функционирования генома. Так, например, показано, что мутации, нарушающие связь центромер с ядерной мембраной, вызывают потерю хромосом, нарушения и задержку митоза, нарушение взаимодействия между микротрубочками и кинетохорами [2]. Существуют многочисленные данные о существовании связи между специфическими участками хромосом и ядерной мембраной [10–12]. Это указывает на то, что выявленное в данной работе четкое расположение хромосом в митотически делящихся клетках гаплоидного растения кукурузы — неслучайно.
Учитывая эти факты рассмотрим изображенную на Рисунке 4 фазу митоза более детально. Отнесение момента данного расположения хромосом к поздней профазе — ранней прометафазе выбрано методом исключения. На данном препарате зафиксировано то состояние, когда полностью завершена конденсация хромосом. Следовательно, можно говорить о том, что данное расположение хромосом может соответствовать концу профазы [1]. Отсутствие ядерной мембраны указывает на то, что это прометафаза. С другой стороны, здесь нет пока еще характерного для прометафазы беспорядочного расположения хромосом, которое возникает вследствие неодновременного присоединения кинетохорных микротрубочек к кинетохорам сестринских хроматид и, следовательно, неодновременного движения хромосом [1]. Это означает, что в данном случае получена фотография очень краткого момента, когда завершившие конденсацию хромосомы еще не начали своего движения под действием кинетохорных трубочек.
В стадии, представленной на Рисунке 4, сестринские хроматиды уже полностью изолированы друг от друга, т.е. они превратились в самостоятельные хромосомы. Они располагаются в виде распавшейся ленты, в которой центромеры хромосом (хроматид) находятся на одной линии, положение которой можно с высокой вероятностью рассматривать как следствие формирования данного расположения на экваторе ядра. Принимая это допущение, расположение разделившихся сестринских хроматид, характеризуемое как колинеарное, следует рассматривать как ориентированное по меридианам ядра (Рисунок 4). Это расположение необычно, поскольку в известных нам публикациях отмечается лишь расположение центромер или просто хромосом по экватору веретена деления. В наиболее впечатляющей статье, посвященной пространственному положению хромосом в прометафазе митотически делящейся клетки человека, показано расположение центромер по экватору веретена деления, но при абсолютно свободном расположении плеч хромосом [13]. В митотически делящейся клетке гаплоидного растения кукурузы выявлено еще более упорядоченное расположение хромосом (Рисунок 4). Этот феномен требует дальнейшего изучения.
Необходимо отметить, что выявленный момент характеризуется совмещением черт, присущих самым разным фазам митоза. Например, по ряду признаков мы определили, что выявленное расположение хромосом присуще поздней профазе — ранней прометафазе митоза, хотя четкое обособление сестринских хроматид и даже их центромер характерно для более поздней стадии — метафазы.
Выявленное расположение хромосом не обнаружено в корешках проростков кукурузы, что позволяет предположить его обусловленность спецификой состояния генома клеток воздушного корня. Оценить возможную связь такого расположения хромосом с гаплоидией можно будет при сравнении митоза в воздушных корнях диплоидных и гаплоидных растений кукурузы. Кроме того, не исключено, что выявленное расположение хромосом является результатом сочетания сразу двух факторов: и гаплоидным состоянием генома, и тканевыми особенностями организации генома в клетках воздушных корней. Еще одним возможным фактором, определившим данное расположение хромосом, может быть влияние материнской цитоплазмы тестерной линии кукурузы, использованной для получения гаплоидов и несущей ген ig, вызывающий различные нарушения в функционировании генома. Рассмотрение возможного влияния гена ig на выявленное расположение хромосом ставит вопрос о том, является ли такое расположение нормой или является отражением каких-либо нарушений в ходе митоза? В любом случае анализ этого явления позволит выяснить дополнительные характеристики столь сложного явления, каким является митотическое деление клетки.
Заключение
Необычность выявленного расположения хромосом в поздней профазе –ранней прометафазе митоза хорошо дополняет известные данные о бесчисленных вариациях всех стадий деления клеток в различных тканях у различных видов растений [1]. Все это еще раз указывает на то, что клеточное деление следует рассматривать не как простое распределение генетического материала между дочерними клетками, а как специфичный для каждого вида этап в функционировании генома, в ходе которого распределение генетического материала между дочерними клетками обеспечивается специфической последовательностью функционирования всех элементов митотического аппарата, призванных обеспечить сохранение специфического набора активных и неактивных генов, характерных для того или иного типа делящихся клеток.
Работа поддерживалась бюджетным проектом «Генетический контроль развития и формирования хозяйственно-ценных признаков у сельскохозяйственных растений», №02592021-0012
Список литературы Необычный порядок расположения хромосом в поздней профазе - ранней прометафазе митоза у гаплоидного растения кукурузы, полученного с использованием мутации IG
- Альбертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки. М.: Мир, 1994. Т. 2. С. 400-467.
- Hou H., Zhou Z., Wang Y., Wang J., Kallgren S.P., Kurchuk T., Miller E.A., Chang F., Jia S. Csi1 links centromeres to the nuclear envelope for centromere clustering // Journal of Cell Biology. 2012. V. 199. №5. P. 735-744. DOI: 10.1083/jcb.201208001
- Buttrick G. J., Meadows J. C., Lancaster T. C., Vanoosthuyse V., Shepperd L. A., Hoe K.-L., Kim D.-U., Park H.-O., Hardwickand K. G., Millar J. B. A. Nsk1 ensures accurate chromosome segregation by promoting association of kinetochores to spindle poles during anaphase B // Mol Biol Cell. 2011 V. 22. №23. P. 4486-4502. DOI: 10.1091/mbc.e11-07-0608
- Parodi E. R., Cáceres E. J. Mitosis and cytokinesis in haploid plants of Rhizoclonium hieroglyphicum ssp. hieroglyphicum (Cladophoraceae, Chlorophyta) // Cytologia. 1992. V. 57. №2. P. 203-208. DOI: 10.1508/cytologia.57.203
- Parodi E. R., Cáceres E. J. Life history of freshwater populations of Rhizoclonium hieroglyphicum (Cladophorales, Chlorophyta) // European Journal of Phycology. 1993. V. 28. №1. P. 69-74. DOI: 10.1080/09670269300650111
- Fang Y., Spector D. L. Centromere positioning and dynamics in living Arabidopsis plants // Molecular biology of the cell. 2005. V. 16. №12. P. 5710-5718.
- Chase S. S. Monoploids and monoploid-derivatives of maize (Zea mays L.) // The Botanical Review. 1969. V. 35. №2. P. 117-168.
- DOI: 10.1091/mbc.e05-08-0706
- Kermicle J. L. Androgenesis conditioned by a mutation in maize // Science. 1969. V. 166. №3911. P. 1422-1424.
- DOI: 10.1126/science.166.3911.1422
- Паушева З. П. Практикум по цитологии растений. М.: Агропромиздат, 1988. 271 с.
- White M.J.D. Animal cytology and evolution. Cambridge: Univ. Press, 1954. P. 1-454.
- DOI: 10.1111/J.1558-5646.1955.TB01521.X
- Mikulski P., Hohenstatt M. L., Farrona S., Smaczniak C., Stahl Y., Kaufmann K.,.. Schubert D. The chromatin-associated protein PWO1 interacts with plant nuclear lamin-like components to regulate nuclear size // The Plant Cell. 2019. V. 31. №5. P. 1141-1154.
- DOI: 10.1105/tpc.18.00663
- Gordon M. R., Pope B. D., Sima J., Gilbert D. M. Many paths lead chromatin to the nuclear periphery // BioEssays. 2015. V. 37. №8. P. 862-866.
- DOI: 10.1002/bies.201500034
- Magidson V., O'Connell C. B., Lončarek J., Paul R., Mogilner A., Khodjakov A. The spatial arrangement of chromosomes during prometaphase facilitates spindle assembly // Cell. 2011. V. 146. №4. P. 555-567.
- DOI: 10.1016/j.cell.2011.07.012
