Новая концепция взаимоотношений заразихи и подсолнечника. Сообщение 1 – гипотеза о естественных прививках заразихи и подсолнечника сближением и каллусное происхождение клубеньков

Автор: Зеленцов С.В., Децына А.А., Мошненко Е.В., Илларионова И.В., Хатнянский В.И.
Рубрика: Селекция, семеноводство и биотехнология сельскохозяйственных растений
Статья в выпуске: 1 (201), 2025 года.
Бесплатный доступ
За весь исторический период изучения заразихи и подсолнечника сформировались обоснованные представления о биологии их взаимоотношений. Одновременно накопился целый ряд вопросов, не объясняемых сложившимися представлениями о механизмах первичного контакта проростка заразихи с корнем подсолнечника, образования гаустория, его прорастания в сосудистую систему подсолнечника, формирования клубенька и побега заразихи. Достоверно не установлены причины непрерывного формирования признакового полиморфизма у заразихи, включая преодоление заразихоустойчивости у подсолнечника. Целью настоящих исследований было изучение взаимоотношений заразихи и подсолнечника на основе естественного механизма спонтанных прививок сближением. Исследования проводили в 2021–2023 гг. в ФГБНУ ФНЦ ВНИИМК, г. Краснодар, на сортах подсолнечника СПК плюс и ВНИИМК 8883 улучшенный и на семенах заразихи рас G и F в соотношении 50: 50, собранных в 2021–2023 гг. в Краснодарском крае, Воронежской и Волгоградской областях. Экспериментально подтверждена возможность срастания взрослых побегов заразихи и подсолнечника в результате реципрокных прививок между ними. Сформулирована гипотеза о срастании первичного проростка семени заразихи с корнем подсолнечника как естественной межвидовой прививке сближением с образованием плотного каллуса из тканей привоя в виде недифференцированного клубенька заразихи. Тогда сформировавшиеся из клубеньков побеги заразихи могут представлять собой каллусные регенеранты с ненулевой вероятностью образования генетических рекомбинаций по различным признакам, в том числе по преодолению заразихоустойчивости подсолнечника. В наших исследованиях на отдельных корнях подсолнечника обнаруживали побеги заразихи с различной окраской. При проращивании семян заразихи на нетипичных средах также обнаруживали отдельные проросшие семена. Эти факты подтверждают гипотезу о непрерывном формировании генетического полиморфизма заразихи, вызванного клеточным рекомбиногенезом, свойственным каллусным тканям, к которым вполне можно отнести и клубеньки заразихи.
Заразиха, подсолнечник, клубеньки заразихи, гаустории, прививка сближением, каллус, сомаклональная изменчивость in vivo, каллусные регенеранты
Короткий адрес: https://sciup.org/142244400
IDR: 142244400 | УДК: 633.854.78:632.531 | DOI: 10.25230/2412-608X-2025-1-201-5-20
New concept of the relationship between broom-rape and sunflower. Report 1 - Hypothesis of natu-ral approach grafting of broomrape and sunflower and callus origin of tubercles
Throughout the history of the study of broomrape and sunflower, wellfounded ideas about the biology of their relationship have been formed. At the same time, a number of questions have accumulated that cannot be explained by the established ideas about the mechanisms of the primary contact of the broomrape seedling with the sunflower root, the formation of haustoria, its germination in the sunflower vascular system, and the formation of tubercles and shoots of the broomrape. The reasons for the continuous formation of trait polymorphism in broomrape, including the overcoming of broomrape resistance in sunflower, have not been reliably determined. The aim of this research was to study the relationship between broomrape and sunflower based on the natural mechanism of spontaneous approach grafting. The researches were conducted in 2021–2023 at V.S. Pustovoit AllRussian Research Institute of Oil Crops, Krasnodar on sunflower varieties SPK plus and VNIIMK 8883 improved, and on broomrape seeds of races G and F in the ratio of 50:50, collected in 2021–2023 in Krasnodar, Voronezh and Volgograd regions. The possibility of merging of adult shoots of broomrape and sunflower as a result of reciprocal grafting between them was experimentally confirmed. The hypothesis of merging of the primary seedling of broomrape seed with the sunflower root as a natural interspecific approach grafting with the formation of a dense callus from the scion tissues in the form of undifferentiated broomrape tubercles was formulated. Then, the broomrape shoots formed from tubercles may represent callus regenerants with nonzero probability of genetic recombination formation for various traits, including overcoming broomrape resistance of sunflower. In our studies, broomrape shoots with different coloration were found on individual sunflower roots. When broomrape seeds were germinated on atypical media, individual germinated seeds were also found. These facts support the hypothesis of continuous formation of genetic polymorphism of broomrape caused by cellular recombinogenesis inherent in callus tissues, to which broomrape tubercles can be attributed.
Текст научной статьи Новая концепция взаимоотношений заразихи и подсолнечника. Сообщение 1 – гипотеза о естественных прививках заразихи и подсолнечника сближением и каллусное происхождение клубеньков
Введение. За практически столетний период изучения заразихи в России и в мире сформировался набор постулатов, определяющий общепринятые представления о биологии взаимоотношений заразихи и подсолнечника. Многочисленные анатомические исследования показывают, что зародыш заразихи редуцирован до стадии позднего проэмбрио, окружённого одноклеточным слоем эндосперма, и не имеет дифференцированные корешок и почечку [1; 2; 3]. Тем не менее в присутствии корневых выделений подсолнечника апикальная часть редуцированного зародыша заразихи прорастает и формирует подобие первичного проростка, который отрастает в направлении близлежащего корня подсолнечника [2; 4]. Поскольку настоящий зародышевый корешок в семени заразихи отсутствует, но его подобие всё же отрастает из семени, причём при контакте с корнем подсолнечника формирует утолщение, он получил название «гаусторий» – присоска [1; 2; 4] (рис. 1). Общепринятой функцией гаустория заразихи является прикрепление к корню подсолнечника, проникновение внутрь его тканей и поглощение питательных веществ растения-хозяина [1; 3].
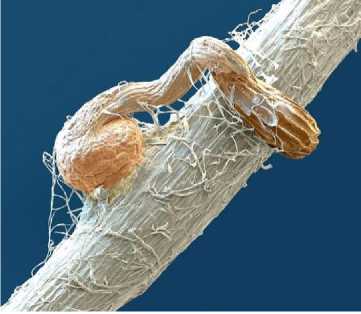
Рисунок 1 – Формирование гаустория при контакте проростка заразихи с корнем подсолнечника под электронным микроскопом (цит. по: [5])
После проникновения гаустория заразихи в корень подсолнечника и установления связи с его сосудистой системой, гаусторий увеличивается в размерах, преобразуясь в клубенёк (а), причём формирование клубенька начинается в зоне прикрепления гаустория к корню. Затем в клубеньке закладывается точка роста, из которого формируется побег заразихи (б) [2; 3] (рис. 2).
Такое представление о внедрении заразихи в корень подсолнечника предельно наглядно и многократно было подтверждено экспериментально. Но при анализе предполагаемого процесса прикрепления и врастания гаустория заразихи в корень подсолнечника возникает ряд вопросов, не объясняемых сложившимися представлениями.

а б
Рисунок 2 – Формирование клубенька (а) и начальный рост побега заразихи (б) на корне подсолнечника (ориг.)
То, что семя заразихи не в состоянии самостоятельно прорасти без наличия неких компонентов корневых выделений подсолнечника и некоторых других видов растений, давно и обстоятельно доказано [1; 2; 3]. В эндосперме семян заразихи мало запасных питательных веществ для инициации самостоятельного построения клеточных стенок первичного проростка и гаустория [3]. Соответственно, главным источником ростостимулирующих метаболитов и глюкозы для постройки целлюлозных клеточных стенок при отрастании первичного проростка заразихи являются корневые выделения подсолнечника [2; 3].
Но с позиций цитоморфогенеза для формирования целлюлозных стенок клеток проростка заразихи, помимо наличия питательных веществ, поступивших из корня подсолнечника, обязательна гормональная и ферментативная инициация синтеза целлюлозы и деления клеток апекса. Следовательно, необходимые для инициации роста первичного проростка заразихи метаболиты также должны входить в состав корневых выделений подсолнечника. И в целом можно констатировать, что на этапе прорастания семени заразиха выступает только в качестве пассивного потребителя целого комплекса биохимических компонентов корневых выделений подсолнечника, который становится для заразихи хеморегулятором [2].
Многочисленные исследования показывают, что практически весь комплекс веществ, синтезируемых гаусториями заразихи, прежде всего, ориентирован на разрушение межклеточного пектина и клеточных стенок подсолнечника [3; 6; 7; 8]. А откуда у проростка заразихи материальные ресурсы и энергия на формирование и на активную (предположительно) работу такого органа, как гаусторий? Микроскопическое семя заразихи размерами 0,36–0,50 × 0,16–0,25 мм [9], состоящее всего из нескольких десятков клеток [1; 3], почти никаких запасных метаболитов не содержит. Есть сведения, что некоторые биохимические компоненты в семенах и апексе проростка заразихи всё же были выделены. Среди них липиды, белки, экстрацеллюлярные ферменты, пероксидаза, аспарагиназа и ряд других соединений [6; 7; 8; 10; 11; 12]. Но все эти метаболиты фиксировались по факту, без определения их массовой доли и достаточности для материального обеспечения процесса формирования гаустория. Но в целом проросток заразихи для своего роста в направлении корня подсолнечника и преобразования в гаусторий пользуется метаболическими ресурсами корневых выделений будущего растения-хозяина [1].
Предположим, что в геноме заразихи имеется некий комплекс генов, кодирующий формирование гаустория за счёт выделяемых корнями подсолнечника метаболитов. Но по аналогии с гаусториями других видов растительных организмов предполагаемая биологическая функция присоски – прикрепление (присасывание, приклеивание) к поверхности корня с последующим активным забором питательных веществ из тканей и сосудистой системы растения-хозяина [13; 14]. Но чтобы эти вещества изъять у подсолнечника, гаусторий должен иметь некий рабочий механизм для активного забора водных растворов питательных веществ из тканей корня подсолнечника. И для этого гаусторию заразихи необходимо иметь дополнительные специализированные органы или физиологические механизмы, а их никто пока не обнаруживал.
Далее в соответствующей литературе отмечается, что гаусторий заразихи раздвигает клетки корня растения-хозяина за счёт разрушения межклеточного пектина и прорастает к сосудам ксилемы корня [3; 10]. Но для разрушения межклеточного пектина необходимы специализированные ферменты. И такие ферменты у гаусториев заразихи действительно описаны [3; 6; 7; 8].
Но на этой стадии опять возникают вопросы. А откуда в гаустории появились экстрацеллюлярные ферменты? Если они были синтезированы в самом гаустории, то откуда взялись материальные ресурсы для их синтеза? Возможно, апикальная часть гаустория заразихи использует некие уже готовые ферменты и фитогормоны, содержащиеся в корневых выделениях подсолнечника. Но если эти соединения были изначально синтезированы в тканях корня подсолнечника, то почему они не приводили к разрушению или растворению межклеточного пектина до момента контакта с гаусторием заразихи?
Может гаусторий эти ферменты подсолнечника как-то преобразует в химически более активные формы? Возможно. Но для их преобразования необходимо иметь достаточные ресурсы для запуска соответствующих химических реакций. А семя заразихи, его первичный проросток и гаусторий, по умолчанию, своих ресурсов практически не имеют и используют только внешние ресурсы из корневых выделений подсолнечника.
Допустим, что неизвестно откуда взявшиеся у заразихи экстрацеллюлярные ферменты так или иначе, но растворили межклеточный пектин в тканях корня подсолнечника, и гаусторий пророс до сосудов ксилемы. А дальше для врастания в сосуды ксилемы опять нужны специализированные активные метаболиты для растворения или раздвигания целлюлозных волокон стенок сосудов в месте контакта. Затем гаусторию нужно срастись с ксилемой с образованием единой сосудистой системы. И снова возникают вопросы – как и за счёт чего? Да, можно ответить, что за счёт ресурсов растения-хозяина, но эти ресурсы должны поступить внутрь клеток гаустория на основе некоего механизма поглощения, затем гаусторий должен их использовать для синтеза соответствующих активных соединений, а это опять специфические химические реакции, специализированные ферменты для их регуляции и энергия для запуска этих реакций.
После врастания гаустория заразихи в ксилему корня подсолнечника наружные ткани проростка заразихи начинают диффузно разрастаться, формирую клубенёк с последующей эндогенной закладкой в нём точки роста и отрастанием из неё побега заразихи. Вот на этом этапе у заразихи уже появляется неиссякаемый источник водно-минерального питания и пластических веществ, который позволяет ей паразитически развиваться [1; 3].
Но и на этапе формирования клубенька заразихи на корне подсолнечника возникают вопросы. Так, по Э.С. Терёхину (1977) образование гаустория и клубенька – это промежуточные этапы подготовки к развитию воспроизводящей половой системы заразихи [1]. Но почему вросший в корень подсолнечника гаусторий не сразу формирует побег, а сначала образует клубенёк? Зачем заразихе такое усложнение стадий развития? Вместо типичного для цветковых растений двухэтапного развития « семя – растение » у неё формируются дополнительные промежуточные этапы и органы: « семя – гаусторий – клубенёк – растение ». Можно, конечно, сослаться на то, что это характерно для паразитического образа жизни заразихи, но это формальный аргумент, который ничего не объясняет.
Какова же функция промежуточного клубенькового этапа у заразихи? Зачем на эту стадию тратятся драгоценные, с такими усилиями изъятые у подсолнечника ресурсы? Вот у папоротниковидных между прорастанием споры и формированием растения имеется промежуточная стадия в виде развития из споры листовидного заростка – гаметофита, на котором происходит оплодотворение яйцеклетки, и из зиготы отрастает побег [15]. Может в клубеньках, как у папоротниковидных, происходит оплодотворение? Нет, все виды рода Orobanche , включая заразиху кумскую Or-obanche cumana Wallr., – цветковые растения, соответственно, у всех видов заразихи оплодотворение и формирование семян происходит в цветках [1; 2; 3]. И в целом при общем анализе литературы так и остались не прояснёнными биологический смысл образования и функциональность клубенька заразихи до этапа формирования из него цветоносного побега.
Материалы и методы. Для экспериментальной проверки и обоснования разрабатываемой концепции взаимоотношений заразихи и подсолнечника, на сортах подсолнечника СПК плюс и ВНИИМК 8883 улучшенный в 2021–2023 гг. на базе ФГБНУ ФНЦ ВНИИМК, г. Краснодар, в полевых условиях на инфицированном заразихой участке и в условиях искусственного климата в осенне-зимний период закладывали модельные эксперименты. Эти же сорта подсолнечника использовали для получения водных растворов корневых выделений в модельных экспериментах по отдельным аспектам прорастания семян заразихи. Для исследований брали смесь семян заразихи, собранной в 2021–2023 гг. в Каневском, Кореновском и Тихорецком районах Краснодарского края, в Богучарском районе Воронежской области и в Михайловском районе Волгоградской области. Лабораторией иммунитета ВНИИМК была установлена принадлежность семян к расам G (50 %) и F (50 %).
Результаты и обсуждение. Рассмотрим взаимоотношения заразихи и подсолнечника с ботанической точки зрения. Подсолнечник однолетний Helianthus an-nuus L. относится к семейству Астровые порядка Астроцветные, а заразиха кумская Orobanche cumana Wallr. – к семейству За-разиховые, ранее порядка Норичниковые [2], а с 2016 г. отнесена к порядку Ясноткоцветные [16]. Но таксономически семейства Астровые и Заразиховые очень далеки друг от друга [2; 17]. Поэтому заразиха и подсолнечник должны быть физиологически несовместимы. Однако в реальности заразиха кумская вполне способна не просто врасти в корни подсолнечника, но активно и успешно использовать для своего роста и развития продукты метаболизма таксономически очень отдалённого растения-хозяина. Следовательно, в природе должны быть некие естественные механизмы, обеспечивающие совместное сосуществование неродственных видов растений.
И действительно, подобные факты совместного сосуществования разных видов высших растений с частично объединёнными сосудистыми системами, хотя и довольно редкие, но известны. В естественной природе такие случаи осуществляются за счёт самого древнего, естественного механизма спонтанной прививки сближением побегов или корней, она же растительная иноскуляция, между рядом стоящими хвойными или цветковыми растениями. Способствует развитию иноску-ляции тесное произрастание соседних растений, которые при этом плотно касаются друг друга центральными или боковыми побегами и за счёт взаимного трения нарушают целостность своей коры, и начинают контактировать тканями оголённого камбия [18; 19; 20; 21]. А у корней и трения не требуется, достаточно плотного касания молодых корешков до образования на них опробковевшей коры [22; 23]. Такие явления, преимущественно, отмечаются между растениями одного вида, но изредка встречаются и между представителями различных таксонов [24]. В частности, в горных лесах Западного Кавказа нами был обнаружен случай межвидовой естественной прививки сближением плюща обыкновенного Hedera helix L. (сем. Аралиевые, порядок Зонтикоцветные) с ясенем высоким Fraxinus excelsior L., (сем. Маслиновые, порядок Ясноткоцветные) с объединением сосудистых систем в зоне срастания (рис. 3).
плющ ясень
Рисунок 3 – Срастание побега плюща обыкновенного со стволом ясеня высокого. Горный лес в Туапсинском районе Краснодарского края (ориг.)
Явление спонтанной прививки соседних растений сближением их побегов некогда подметил И.В. Мичурин, на основе чего разработал технологию межродовых и межсемейственных прививок за счёт использования промежуточных «менторов», представляющих собой промежуточные прививки сближением между таксономически неродственными растениями, с частичным сохранением собственных сосудистых систем, с целью постепенного «привыкания» побегов к использованию чужих метаболитов [25]. В результате И.В. Мичурину через предварительные прививки методом сближения побегов разных видов удалось привить даже лимон на грушу (рис. 4).

Рисунок 4 - Шестимесячный прививок лимона на груше (цит. по: [25])
Заразиха и подсолнечник также таксономически отдалены друг от друга, как мичуринские лимон и груша. Но если провести определённые подготовительные «менторные» процедуры постепенного сближения, то, теоретически, заразиха может прививаться на подсолнечник.
Исходя из факта, что материнское растение заразихи, произрастая на подсолнечнике, фактически полностью развивается на метаболитах его корней, то процедура биохимического сближения между этими видами уже проведена естественным путём и не один раз. По данным В.К. Морозова (1947), первые сообщения о заразихе на подсолнечнике в России (Саратовская губерния) зафиксированы в 1890-е годы [цит. по: 26]. По данным Т.С. Антоновой (2018), первые сообщения о массовом поражении подсолнечника заразихой в Воронежской губернии относятся к 1866 г. [3]. Это указывает на то, что история совместного сосуществования заразихи и подсолнечника насчитывает очень большое количество поколений, даже с учётом длинных севооборотов [27; 28]. Тогда подсолнечник является типичным мичуринским ментором для заразихи. Следовательно, и биохимический состав цитоплазмы тканей проростка заразихи тоже будет близок или даже аналогичен цитоплазме подсолнечника. Поэтому современные расы заразихи полностью адаптированы к использованию для своего роста и развития биохимических компонентов клеточного сока этого растения-хозяина. Развивая эту мысль, можно предположить, что в таком случае побег заразихи можно непосредственно привить на подсолнечник без промежуточных этапов сближения по И.В. Мичурину, и, теоретически, привой заразихи может срастись с подвоем подсолнечника.
Для экспериментальной проверки этого предположения в 2022–2023 гг. нами были проведены прививки побегов заразихи на подсолнечник. Прививки осуществляли разными методами - в расщеп, в боковой зарез, под кору, на пенёк. Максимальная приживаемость была зафиксирована при прививке в расщеп, который обеспечивал самую большую контактную поверхность привоя и подвоя. И даже в жарких полевых условиях с низкой влажностью воздуха такой способ прививки обеспечивал положительный водный баланс между поступающей из подвоя и испаряющейся с поверхности привоя воды. Через 2 недели после прививки в контактной зоне отмечена начальная стадия формирования раневого каллуса. Успешно привившиеся побеги заразихи увеличивались в размерах и зацветали (рис. 5).

а б в
Рисунок 5 – Динамика развития привитого побега заразихи от бутонизации до цветения на боковом побеге подсолнечника, сорт СПК плюс, полевой инфицированный фон
ВНИИМК, 2022 г. (ориг.): а) 3-е сутки после прививки; б) 10-е сутки после прививки;
в) 13-е сутки после прививки
Часть привоев заразихи через 1–2 суток после прививки теряла тургор и засыхала, однако практически во всех случаях это было вызвано неполным совпадением сосудистых тканей при совмещении привоя и подвоя в процессе прививки или подсыханием тканей подвоя ниже зоны прививки. Но в целом проведённый эксперимент показал, что заразиха может быть успешно привита на подсолнечник.
Но если у вегетирующей заразихи метаболические продукты близки или идентичны подсолнечнику, то, теоретически, и подсолнечник можно привить на заразиху. С этой целью в 2022–2023 гг. осуществляли экспериментальные прививки побегов подсолнечника в расщеп на побеги заразихи. Через 2 недели после прививки привои подсолнечника сохраняли жизнеспособность и тургор. Прирост биомассы привоев подсолнечника на заразихе визуально не зафиксирован вследствие ещё не сформировавшейся единой сосудистой системы. Тем не менее у привоя подсолнечника на 11 сутки после прививки было отмечено небольшое увеличение ширины верхнего листочка (рис. 6).
В целом, проведённые исследования не выявили каких-либо физиологических барьеров для реципрокных прививок заразихи и подсолнечника. Привитые побеги заразихи на подсолнечник и подсолнечника на заразиху успешно приживались. Часть привитых побегов заразихи на подсолнечных подвоях развивалась до стадии бутонизации и цветения. Таким образом, возможность успешного использования привоями заразихи и подсолнечника воды и метаболитов из подвоев, соответственно, подсолнечника и заразихи можно считать доказанной.

Рисунок 6 – Жизнеспособность побега подсолнечника после прививки на заразихе, сорт ВНИИМК 8883 улучшенный, теплица ВНИИМК, 2023 г. (ориг.): а) 2-е сутки после прививки;
б) 11-е сутки после прививки
На основании результатов проведённых исследований была выдвинута очень необычная гипотеза, что срастание первичного проростка заразихи с корнем подсолнечника тоже может быть процессом естественной прививки сближением, а не актом паразитического проникновения гаустория в ткани корня. В случае естественной прививки все метаболические и энергетические ресурсы к месту контакта будут поставляться подвоем, в данном случае подсолнечником. А проросток заразихи в качестве привоя на начальном этапе срастания будет лишь потребителем и транслятором каллусогенных метаболитов подвоя.
Первым характерным признаком успешной прививки, помимо сохранения жизнеспособности привоя, является образование каллусной ткани в месте контакта [29]. Каллус представляет собой группу недифференцированных клеток на раневом или контактном участке, образовавшихся за счёт дополнительного притока к этим участкам фитогормонов, прежде всего, цитокининов. Существует несколько типов каллуса, в том числе рыхлый и плотный [30; 31; 32].
Рыхлый каллус формируется при избытке цитокининов, плотный – при их недостатке. Рыхлый каллус представляет собой группу слабо связанных между собой паренхимных клеток без выраженных пектиновых межклеточных пластинок. Такой тип каллуса легко может быть разделён на отдельные клетки в суспензии. Плотный каллус, наоборот, отличается формированием прочных пектиновых межклеточных пластинок, обеспечивающих тесную связь клеток каллуса. Поэтому форма плотного каллуса более компактная и почковидная (рис. 7) [30; 31; 32; 33; 34; 35].
При этом плотный тип каллуса способен к быстрой дифференциации клеток с формированием на своей поверхности ре-генерантных побегов и образованием сосудистых систем через предварительное формирование протососудов – гидроцитов [33; 35]. Перевод рыхлого каллуса в плотный и обратно регулируется изменением концентрации цитокининов [33; 34].
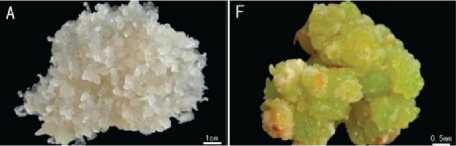
Рисунок 7 – Внешний вид рыхлого (А) и плотного (F) типов каллуса (цит. по: [30])
Рассмотрим с позиций гипотезы каллу-сообразования внешний вид формирующегося клубенька заразихи и прилегающих тканей корня подсолнечника (рис. 8).

Рисунок 8 – Формирующийся клубенек заразихи (а) и сопутствующее разрыхление прилегающей ткани корня подсолнечника (б), сорт ВНИИМК 8883 улучшенный (ориг.)
На вершине клубенька заметны остатки семенной оболочки. Граница тканей клубенька и корня различается довольно отчётливо. Слева, ниже клубенька на корне подсолнечника заметна группа рыхлых клеток. Именно эти рыхлые клетки и послужили визуальным основанием для классического утверждения, что гаусторий заразихи паразитически разрыхляет ткань корня подсолнечника и проникает к сосудистой системе.
Но если рассматривать эту разрыхлённую ткань корня с позиций каллусогенеза, то она очень похожа на типичный рыхлый каллус с избытком цитокинина, поступающего по сосудистой системе корня подсолнечника к месту контакта. Тогда в рамках нашей гипотезы сам клубенёк заразихи можно квалифицировать как комок плотного каллуса.
В качестве модели можно рассмотреть явление, когда в образующемся после прививок плодовых культур плотном каллусе формируются водопроводящие удлинённые клетки – гидроциты, представляющие собой протососуды, через которые устанавливается начальная связь с сосудистыми системами подвоя и привоя. Затем гидроциты преобразуются в полноценные сосуды проводящей системы. Визуально клетки гидроцитов выделяются в плотном каллусе более интенсивной окраской [33; 34; 36].
Окраска поперечного среза клубенька заразихи красителем метиленовый синий также, как и в каллусах плодовых прививок, позволила выделить в его тканях более интенсивно окрашенные, удлинённые клетки гидроцитов, причём уже ориентированные в направлении корня подсолнечника (рис. 9).
корень подсолнечника клубенёк заразихи гидроциты
Рисунок 9 – Продольное сечение клубенька заразихи с темноокрашенными протососудами – гидроцитами (ориг.)
Таким образом, визуальный анализ рисунков 8 и 9 даёт начальные подтверждения концепции прививки сближением первичного проростка заразихи на корень подсолнечника с последующим образованием в области их контакта двух типов каллусов: рыхлого – в тканях корня подсолнечника, и плотного – из ткани первичного проростка заразихи.
В рамках этой же концепции можно объяснить и формирование присосковидного утолщения (гаустория) в апикальной части проростка заразихи при его контакте с корнем подсолнечника. Через контактную поверхность в проросток заразихи из подсолнечника начинают поступать каллусогенные метаболиты. Соответственно, апикальная часть ткани проростка в месте контакта с корнем подсолнечника в первую очередь получает эти метаболиты и переходит в стадию каллусогенеза, которая и визуализируется в виде присосковидного утолщения (см. рис. 1). Затем процесс каллусогенеза активируется по всему проростку заразихи, который визуально проявляется в формировании клубенька (см. рис. 2а). Тогда и упоминаемая в литературе функция приклеивания гаустория к поверхности корня [13] вполне может быть объяснена синтезом пектина при каллусогенезе, обеспечивающем тесный контакт клеток плотного каллуса. При этом все метаболические и энергетические ресурсы к месту контакта будут поставляться подвоем, в данном случае подсолнечником. А проросток заразихи, в качестве привоя, на начальном этапе срастания будет лишь потребителем и транслятором каллусогенных метаболитов подвоя.
Каллусогенез, как биологическое явление, довольно хорошо изучен в мире. Одним из известных и хорошо изученных свойств каллусных культур является их генетическая нестабильность, по разным оценкам, вследствие регулярных мутаций, хромосомных аберраций, генных рекомбинаций (дупликаций, делеций, инверсий) [31; 37; 38]. Причём, чем моложе эксплантат, тем выше будет потенциальная генетическая гетерогенность полученной из него каллусной культуры. При последующем получении растений-регенерантов из каллуса высока вероятность обнаружения среди них форм с генетическими рекомбинациями. Именно на этой потенциально высокой гетерогенности каллусных культур и была основана технология клеточной селекции растений на основе каллусных регенерантов со вновь приобретёнными генотипическими отличиями от исходного организма-донора [31; 38; 39].
Мировая история селекции подсолнечника на заразихоустойчивость свидетельствует о регулярном появлении новых, более агрессивных рас заразихи, легко преодолевающих защиту ранее уже имеющихся у подсолнечника генов заразихоустойчивости [3; 26; 40]. При этом причины появления новых рас заразихи так и не были однозначно установлены. Поэтому современная селекция подсолнечника на заразихоустойчи-вость, фактически, ведётся «вдогонку», только после появления очередной расы и на основе поиска устойчивых к ней форм.
Если принять за гипотетическую основу, что клубенёк заразихи представляет собой плотную каллусную ткань, сформировав- шуюся из молодого первичного проростка при прививке сближением с корнем подсолнечника, то часть клеток клубенька, по аналогии с сомаклональной изменчивостью каллусов in vitro, вполне может быть подвержена аналогичным генетическим рекомбинациям, только in vivo. Тем более что проросток заразихи, как эксплантат, очень молод, соответственно, потенциальная генетическая гетерогенность образующегося из него каллуса должна быть достаточно высокой. И в случае формирования регенерант-ного побега заразихи из одной из рекомбинантных клеток клубенькового каллуса полученное семенное потомство этого побега генетически будет отличаться от генома материнского растения заразихи. Тогда это и есть ежегодно пополняющийся на любом заражённом поле подсолнечника неиссякающий источник генетического полиморфизма заразихи. А условия выращивания и наличие генов устойчивости сорта или гибрида подсолнечника к той или иной расе заразихи для таких рекомбинантных побегов растения-паразита становятся лишь дифференцирующим фоном естественного отбора.
Прямыми экспериментами доказать наличие рекомбиногенеза в каллусной ткани клубенька заразихи непросто. Но сам факт регулярного появления новых рас заразихи как раз и подтверждает, как минимум, периодическое образование генетических рекомбинаций у заразихи.
Ранее было замечено, что корневые выделения некоторых культурных растений вызывают частичное прорастание семян заразихи, но без последующего внедрения в их корневые системы. К таким культурам относили отдельные гибриды кукурузы, сорта гороха, сои, сорго, проса, суданской травы [12; 41; 42; 43]. Но оказалось, что не так и много семян заразихи провокационно прорастало в корневых зонах этих культур. Большая часть оставалась в режиме ожидания до очередного появления на этом поле подсолнечника. Но в целом способность части семян заразихи прорастать в корневых выделениях других видов растений также свидетельствует об их рекомбинантной природе.
При этом причины прорастания семян заразихи в корневых выделениях других культур, но полного отсутствия фактов внедрения гаусториев в их корни, так и не были выяснены. Хотя в рамках концепции прививок сближением проростков заразихи с корнями подсолнечника на фоне их полной физиологической и биохимической совместимости эта дилемма вполне объяснима, например, наличием сомаклональ-ной генетической изменчивости в каллу-сных клубеньках заразихи и последующим формированием из них регенерантных побегов. А вот прививки сближением между таксономически отдалёнными видами, в данном случае заразихи с кукурузой, горохом, соей и другими культурами, возможны только после мичуринской «менторской» процедуры постепенного физиологического и биохимического сближения заразихи с этими видами. А поскольку проростки заразихи ранее не были адаптированы к биохимическим особенностям тканевых метаболитов вышеперечисленных культур, то между ними возникала физиологическая несовместимость, препятствующая прививкам и последующему образованию каллусных клубеньков.
Крупнейший в России специалист по заразихе Т.С. Антонова с коллегами в своих исследованиях выделяла несколько морфологически отличающихся форм заразихи. Исходный, классический фенотип заразихи -это отрастающий из клубенька одиночный штамбовый побег, формирующий соцветие в верхней половине побега. В результате исследований ими была обнаружена и описана фенотипически новая форма заразихи, у которой побег полностью представлял собой цветонос с формированием цветков даже ниже уровня поверхности почвы [3; 44; 45]. Ещё одна выделенная Т.С. Антоновой с коллегами форма заразихи - многостебельная, у которой из одного клубенька отрастают несколько побегов заразихи [3; 44; 46]. В рамках концепции о сомакло-нальной изменчивости in vivo каллусной ткани клубеньков такие морфологически отличающиеся формы заразихи можно отнести к генетическим рекомбинантам.
С целью иллюстрации наличия признакового полиморфизма у заразихи мы визуально оценивали окраску побегов заразихи в сосудах с растениями сорта подсолнечника ВНИИМК 8883 улучшенный и провели модельный эксперимент по улучшению возможности прорастания семян заразихи на разных средах, помимо выделений корней подсолнечника (рис. 10 и 11).

а
Рисунок 10 – Разнообразие окраски побегов заразихи: от желтой (а) до фиолетовой (б), на корнях подсолнечника сорта
ВНИИМК 8883 улучшенный (ориг.)

б
На рисунке 10 видно, что даже в пределах сосуда в теплице окраска отдельных побегов заразихи, сформировавшихся на корнях подсолнечника ВНИИМК 8883 улучшенный, может заметно отличаться. Подобные факты различий по окраске побегов заразихи давно известны и нередко встречаются при рутинной селекционной оценке генотипов подсолнечника на зара-зихоустойчивость. Однако не ясно, чем подобный полиморфизм определяется: уже имевшейся гетерогенностью семенного материала заразихи, подмешанного в почву сосуда, или это свежие рекомбинации в данном поколении регенерантов из каллу-сных клубеньков. Тем не менее факт полиморфизма заразихи по признаку окраски побегов вполне очевиден.
Для выявления потенциального полиморфизма заразихи по признаку прорастания семян в нетипичных средах был поставлен простой модельный эксперимент. Примерно по 2 тысячи семян заразихи были помещены в вытяжки из клеточного сока стеблей подсолнечника, в водный раствор корневых выделений сафлора и в чистую водопроводную воду. Клеточный сок стеблей подсолнечника был взят, поскольку его биохимический состав, как минимум частично, подобен составу клеточного сока его корней и корневых выделений. Корневые выделения сафлора были взяты потому, что этот вид также относится семейству Астровые, хотя роды Сафлор и Подсолнечник таксономически довольно отдалены друг к друга. А вариант с чистой водой служил условным контролем.
Из примерно 2 тысяч семян заразихи на вытяжке клеточного сока из стеблей подсолнечника сорта ВНИИМК 8883 улучшенный через 11 суток замачивания было обнаружено 20 проростков заразихи. Из примерно такого же количества семян заразихи, замоченных в корневых выделениях сафлора, был обнаружен один проросток. На варианте с помещением 2 тысяч семян заразихи в чистую водопроводную воду через 11 суток было выявлено восемь проростков. Последний результат выглядит фантастично, но тем не менее факт прорастания семян заразихи в чистой воде зафиксирован (рис. 11).

а б

в
Рисунок 11 – Прорастание отдельных семян заразихи (ориг.):
а) в клеточном соке стеблей подсолнечника; б) в корневых выделениях сафлора;
в) в чистой водопроводной воде
Прорастание семян заразихи в чистой воде является крайне редким событием. Уникальность здесь не столько в экзотике, сколько в необычном факте наличия в этих семенах достаточного объёма запасных веществ и фитогормонов для самостоятельного прорастания в воде. Эти восемь проростков заразихи – уже не просто генетические рекомбинанты. Гипотетически их можно квалифицировать как рекомбинат-ные рудименты, откатившиеся на филогенетически предыдущую, допаразитическую ступеньку эволюции заразихи. Но если в результате происходящих в каллусной ткани клубеньков рекомбинационных процессов такие формы и появляются эпизодически в семенных потомствах заразихи, то их вредоносность, теоретически, должна стремиться к нулю, поскольку они прорастут при первом же увлажнении почвы и погибнут, как проростки заразихи на корнях растений-провокантов.
Здесь можно сделать предварительный вывод о том, что вышеописанный признаковый полиморфизм у заразихи кумской: от различной окраски побегов и способности прорастать в нетипичных средах до появления новых агрессивных рас, как минимум, частично может быть вызван сома-клональной изменчивостью in vivo каллу-сной ткани клубеньков заразихи.
Заключение. Экспериментально подтверждена возможность реципрокных прививок между взрослыми побегами заразихи и подсолнечника. На основании этого сформулирована гипотеза о срастании первичного проростка (гаустория) заразихи с корнем подсолнечника, которая может представлять собой естественную межвидовую прививку сближением (иноскуляцию) с образованием плотной каллусной ткани привоя в виде недифференцированного клубенька заразихи.
Сформировавшиеся из клубеньков побеги заразихи могут представлять собой каллусные регенеранты. По аналогии с известной сомаклональной изменчивостью каллусных регенерантов in vitro клубеньковые регенерантные побеги заразихи также могут нести в своих геномах генетические рекомбинации по различным признакам, в том числе по преодолению заразихоустойчивости подсолнечника.
Периодически обнаруживаемые на корнях подсолнечника побеги заразихи с различной окраской, а также способность части семян заразихи прорастать на нетипичных средах подтверждают генетический полиморфизм семян заразихи. Это даёт основания считать, что такие мутации и рекомбинации могут быть вызваны, в том числе, и клеточным соматическим реком-биногенезом, свойственным каллусным тканям, к которым, предположительно, вполне можно отнести и клубеньки заразихи.
