Новейшие сведения о пектиновых полисахаридах

Автор: Оводова Раиса Григорьевна, Головченко В.В., Попов С.В., Оводов Юрий Семенович
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 3, 2010 года.
Бесплатный доступ
В данном обзоре представлена информация о пектиновых полисахаридах (пек- тинах), которая получена в последнее время сотрудниками Отдела молекуляр- ной иммунологии и биотехнологии Института физиологии Коми НЦ УрО РАН и рядом других авторов, работающих по тому же направлению. Приводятся сведения о химическом строении пектиновых макромолекул, о выделении пек- тинов из пищевых растений в условиях, имитирующих гастральные, о физио- логической активности пектинов и их фрагментов и о биотехнологическом по- лучении пектинов с помощью каллусных (клеточных) культур.
Пектины, пектиновые полисахариды, пектиновые вещества, растения, иммуномодуляторы, противовоспалительная активность, каллусные культуры, биотехнология пектинов
Короткий адрес: https://sciup.org/14992400
IDR: 14992400 | УДК: 577.114:581.192
The modern information concerning pectic polysaccharides
The present review is devoted to the newest information about pectic polysaccharides (pectins) obtained by scientists of the Department of Molecular Immunology and Biotechnology of the Institute of Physiology, Komi Science Centre, Ural Branch, Russian Acad. of Sci. and some other authors carrying out research in this direction. The data concerning patterns of the pectin macromolecules, isolation of pectins from nutritive plants using conditions related to gastral ones, physiological activity of pectins and their fragments, and biotechnology of pectins by aid of the callus (cellular) cultures are given.
Текст обзорной статьи Новейшие сведения о пектиновых полисахаридах
C момента выхода монографии [1], в которой даны ссылки на статьи, опубликованные до 2007 г., прошло совсем немного времени, но уже за прошедший промежуток о пектиновых полисахаридах появилось столь большое число статей, что нет возможности их процитировать полностью, в связи с чем авторы анализируют в основном результаты собственных исследований последних лет и ряд других данных, полученных в близких областях за тот же период.
Определение и общая схема строения
Пектиновые вещества входят в состав практически всех растений, являются их основными функциональными компонентами, выполняют в растениях множество различных жизненно важных функций и характеризуются широким спектром физиологической активности. Пектины, выделяемые из непищевых растений, представляют основу ряда лекарственных средств и биологически активных добавок, а из пищевых растений являются важным компонентом рациона человека. Последние входят в группу так называемых «пищевых волокон» с большим сродством к организму человека, выработанным за многовековой период их потребления. Они практически не токсичны, недостаток их в pа- ционе питания человека приводит к серьезным нарушениям в обмене веществ.
Пектиновые вещества включают нерастворимый протопектин, растворимые пектиновые полисахариды и сопутствующие им галактаны, арабина-ны и арабиногалактаны [1, 2]. Пектиновые полисахариды (пектины) входят в большую группу глика-ногалактуронанов – кислых растительных полисахаридов, главную углеводную цепь которых составляют 1,4-связанные остатки α-D-галактопиранози-луроновой кислоты [2]:
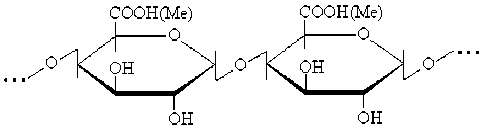
где Ме – метил или ион металла.
Пектины включают в себя как составляющие компоненты нерастворимого протопектина, так и растворимые полисахариды соков растений [2].
Они входят в состав растительных клеток в виде частично метилэтерифицированных по карбоксилу биополимеров часто в виде солей натрия, кальция и магния. Вместе с другими компонентами клеточных стенок растений они обеспечивают их прочность и растяжимость, предохраняют растения от высыхания, обеспечивая засухоустойчивость и морозостойкость, выполняют защитную роль во взаимоотношениях растений с фитопатогенами, способствуют ликвидации повреждений и выходу растения из состояния стресса. Их макромолекулы являются определяющими в прорастании семян и роста растений, в созревании и хранении овощей и фруктов, а структура может существенно меняться в процессе роста и развития растения. Они характеризуются нерегулярным типом строения и рассматриваются как один из самых сложных и динамичных по структуре класс биополимеров [1].
Общая схема строения пектинов приведена во многих работах (в частности, см. [2]). Пектиновые макромолекулы включают в себя следующие основные структурные области:
Линейная область гомогалакту-ронана
Линейная область рам-ногалактуро-нана
Разветвленная область рамногалакту-ронана I (RG-I): боковые цепи: ара-бинан, галактан, арабиногалактан; области ксилога-лактуронана, апио-галактуронана
Линейная область гомогалактуронана состоит из 1,4-связанных остатков α-D-галактопиранози-луроновой кислоты. Эти участки соединяются между собой одним или двумя 1,2-связанными остатками α-L-рамнопиранозы, образуя линейную область рамногалактуронана как главной углеводной цепи (кора) макромолекулы.
Разветвленная область состоит из рамнога-лактуронана I (RG-I), содержащего линейную цепь галактуронана и боковые углеводные цепи из остатков галактозы, арабинозы. Кроме того, возможно наличие в разветвленной области фрагментов кси-логалактуронана, в котором к цепи галактуронана 1,3-гликозидной связью присоединены одиночные остатки β-D-ксилопиранозы, а также апиогалакту-ронана. В последнем одиночные, 1,2- или 1,5-связанные остатки D-апиозы присоединяются 1,2-и/или 1,3-связями к остаткам D-галактуроновой кислоты главной углеводной цепи [2].
Методы установления строения пектинов
Для определения строения пектинов используют классические методы: полный и частичный гидролиз, ферментативное расщепление и периодатное окисление (распад по Смиту), метод метилирования Хеуорзса, а также физико-химические методы: все виды спектроскопии ядерного магнитного резонанса (ЯМР), хромато-масс-спектрометрию [2]. Для определения расположения углеводных цепей пектинов в пространстве в последние годы применяют атомносиловую микроскопию (АСМ), впервые предложенную для этой цели в 1997 г. [3].
Нами АСМ применена, наряду с другими методами, для структурного изучения комарумана, пектина сабельника болотного Comarum palustre L. [4]. При этом экспериментально доказано, что главная углеводная цепь этого пектина является разветвленной и содержит боковые цепи, построенные из 1,4-связанных остатков α-D-галактопиранозилу- роновой кислоты. С помощью ЯМР-спектроскопии показано, что боковые цепи галактуронана присоединены к кору макромолекулы 1,2- и 1,3-гликозид-ными связями. Возможно, что небольшое число боковых цепей присоединено одновременно в положения 2 и 3 кора макромолекулы, о чем свидетельствует АСМ-изображение главной углеводной цепи комарумана (рис. 1), из которого видно, что галактуронан присутствует в виде отдельных линейных молекул, а также разветвленных молекул, составляющих не менее 50% общего числа макромолекул. Галактуронан имеет наряду с главной цепью боковые углеводные цепи различного размера, но довольно длинные (см. рис.1). Молекулы содержат по одной и по несколько боковых цепей, присое-

Рис. 1. АСМ – изображение галактуронана комару-мана.
диненных к одному или нескольким участкам главной цепи галактуронана и состоящих из остатков D-галактуроновой кислоты [4].
В работе [5], в которой представлены результаты продолжающегося исследования пектинов, в частности, пектина незрелых томатов Lycopersicon esculentum var. Rutgers, с помощью АСМ показано, что ранее [6] высказанные соображения о необычной морфологии пектиновых полисахаридов подтверждены нашими исследованиями [4] и работой 2008 г. [7]. В этой связи для пектинов (рис. 2), наряду с классической моделью структуры (рис. 2а), вполне реальной представляется более сложная разветвленная модель (рис. 2 b), которая была предложена ранее [6], где разветвленная область рамногалакту-ронана RG-I представляет собой основной скелет макромолекулы, с присоединенными к нему линейными углеводными цепями галактуронана [5].
На основании полученных результатов изучения с помощью АСМ пектина из незрелых томатов и продуктов его кислотного гидролиза сделан вывод [5], что данный пектин состоит из линейных и разветвленных гомогалактуронанов, а также комплексов, содержащих рамногалактуронан RG-I, разветвленные и линейные фрагменты галактуро-нана. Последние или не связаны с RG-I, или присоединены связями, которые разрушаются в процессе экстракции и выделения пектина. При кислот-
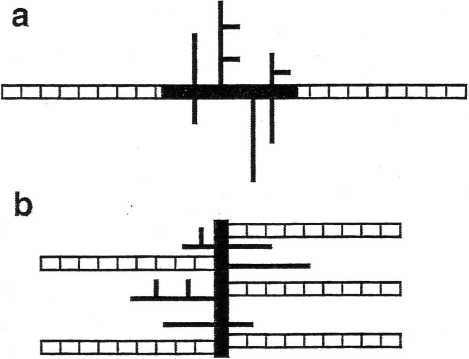
□ HG regions ■ RG-I regions
Рис. 2. Две модели структуры пектина: а) линейная модель, содержащая область: гомогалактуронана (HG) с включением разветвленной области рамнога-лактуронана RG-I; b) модель, изображающая области HG в виде боковых углеводных цепей, присоединенных к RG-I, который также содержит нейтральные боковые цепи.
ном гидролизе остатки нейтральных моносахаридов отщепляются, гидролизуются также RG-I полимеры, что приводит к распаду исходного пектина и получению отдельных его фрагментов. В конечном результате образуется либо линейный галак-туронан, либо его разветвленная часть [5].
Строение пектинов из непищевых растений
Полученные нами ранее данные о структуре и свойствах пектинов из непищевых растений приведены в обзоре [8].
Для выбора наиболее интересных пектиновых полисахаридов проведен скрининг большого числа растений европейского Севера России, при этом принималось во внимание наличие достаточных сырьевых запасов, возможность возведения в культуру, каллусообразование, своеобразие структуры и выраженная физиологическая активность, в частности, иммуномодуляторная и противовоспалительная, после чего проводилось детальное изучение пектинов выбранных растений. В результате были изучены следующие пектины: силенан из смолевки обыкновенной Silene vulgaris Moench (Garke), танацетан из пижмы обыкновенной Tanacetum vulgare L., относящиеся к группе RG-I пектиновых полисахаридов; апиогалактуронановый пектин лемнан из ряски малой Lemna minor L., в сравнении подобным пектином морских трав зосте-раном; своеобразный пектин комаруман из сабельника болотного Comarum palustre L. и бергенан, пектин бадана толстолистного Bergenia crassifolia L. Структура зостерана нами уточнялась несколько раз, однако она еще требует дальнейшего уточнения элементов тонкой структуры [9].
Совсем недавно вышла работа французских исследователей [10], в которой проводится анализ апиогалактуронановых пектинов: зостерана и лем-нана. Авторы, подтверждая наши данные, пытаются уточнить некоторые детали тонкого строения этих пектинов. С этой целью они используют автогидролиз полисахаридов за счет собственных кар- боксильных групп с последующим спектроскопическим анализом образующихся в результате автогидролиза фрагментов макромолекул. Особое внимание они уделяют апиогалактуронану (AGU), полученному из зостерана, и сравнивают его с подобным апиогалактуронаном лемнана, структура которого была установлена нами ранее [11].
Проводя детальное исследование олигоапио-зы, полученной при автогидролизе AGU, с помощью ЯМР-спектроскопии, авторы приходят к выводу, что этот олигосахарид имеет 1,2-гликозидные связи между остатками β-D-апиозы [10] в отличие от 1,5-связей между теми же моносахаридными остатками в апиодисахариде лемнана, структура которого была установлена нами [11]. В этом определенное своеобразие апиогалактуронана зостерана, тем более, что нам в процессе исследования этого пектина не удалось обнаружить олигоапиозные фрагменты апиогалактуронана, вероятно, из-за более жестких условий автогидролиза, но в то же время было четко показано наличие 1,2-, 1,3- и 1,2,3- гликозидных связей терминальных (концевых, одиночных) остатков D-апиозы с остатками D-галактуроновой кислоты главной углеводной цепи зостерана. Таким образом, можно считать еще более уточненной структуру зостерана, апиогалактуронановый фрагмент которого содержит боковые цепи из терминальных остатков апиозы и олигоапиозных фрагментов, состоящих из двух или трех остатков D-апиозы и присоединенных к кору макромолекулы 1,2- и 1,3-гликозидными связями.
При изучении противовоспалительного действия пектинов и их галактуронанов показано, что противовоспалительное действие обусловливается снижением адгезии нейтрофилов человека к фибронектину [17]. Кроме зостерана и лемнана выявлено снижение адгезии галактуронанами следующих пектинов: комарумана, бергенана и коммерческого цитрусового пектина, полученного из кожуры лимона. Структура бергенана была установлена ранее [18] и отмечено, что он представляет собой пектин, относящийся к группе рамногалактуронана RG-I, но при этом главным компонентом углеводной цепи его макромолекулы являются остатки D-галак-туроновой кислоты, содержание их достигает 80%, кроме того, в его состав входят остатки галактозы, арабинозы и рамнозы, общее содержание которых составляет около 15%. Большая часть остатков 1,4-связанной α-D-галактопиранозилуроновой кислоты, составляющей главную линейную цепь галактуро-нана, присутствует в виде метиловых эфиров, чем, вероятно, обусловлена высокая вязкость водных растворов бергенана. Боковые цепи, как во всех пектинах типа RG-I, присоединены 1,4-связью к остаткам α-L-рамнопиранозы кора макромолекулы и построены из терминальных остатков арабинофуранозы и галактопиранозы, а также из небольшого числа остатков 1,5-связанной α-L-арабинофу-ранозы и 1,4,6-связанной β-D-галактопиранозы. Точками разветвления боковых цепей являются 3,4- и 3,6-ди-О-замещенные остатки галактозы. Оказалось, что исходные пектины не влияют на адгезию нейтрофилов. Исключение составляет комаруман, который обладает противовоспалительной активностью [19] и ингибирует адгезию нейтрофилов к фибронектину в концентрации 50 мкг/мл. Это, вероятно, связано с необычной структурой комарума-на, в макромолекуле которого разветвленная главная углеводная цепь галактуронана играет определяющую роль в физиологической активности данного пектина. В других пектинах боковые углеводные цепи, входящие в разветвленную область макромолекулы, препятствуют взаимодействию главной цепи галактуронана с нейтрофилами [17]. Известно, что прикрепление нейтрофилов к фибронектину осуществляется с помощью полипептидов: β1 и β2-интегринов. Предполагается, что галактуро-наны связываются с функционально активными β-интегринами на поверхности нейтрофилов и препятствуют взаимодействию с фибронектином, чем и обусловливают их противовоспалительное действие [17]. Галактуронаны, выделенные из любого пектина, проявляют выраженный противовоспалительный эффект [17, 20]. Дальнейшее изучение исходных пектиновых полисахаридов [20] показало, что они в зависимости от структуры либо способны защищать систему кишечника от повреждений и ингибировать воспаление, либо обладают противовоспалительным действием. Как уже отмечалось выше, комаруман ингибирует развитие кишечного воспаления [19], в то время как зостеран, бергенан, цитрусовый и яблочный пектины не оказывают су- щественного действия на повреждение кишечной стенки при воспалении, а лемнан и силенан, напротив, обладают провоспалительным действием и способствуют увеличению площади поражения при индукции воспаления кишечника [20].
Почти одновременно [21] из стеблей сельдерея Apium graveolens L. нами выделен пектин, который был назван апиуманом. Особенностью его главной углеводной цепи является наличие большого числа метилэтерифицированных остатков D-галактуроновой кислоты. Строение его было установлено и показано, что он является типичным представителем пектинов группы рамногалактуро-нана RG-I. Тем не менее отмечено, что он обладает выраженным противовоспалительным действием. Способность апиумана предотвращать воспаление была изучена на модели эндотоксинового шока, вызванного инъекцией липополисахарида (ЛПС) из кишечной палочки E. coli 0111:В4 (Sigma). Определено, что апиуман заметно увеличивает выживаемость мышей, которым была введена летальная доза ЛПС (рис. 3). В отсутствии апиумана летальная доза инъецированного ЛПС вызывает в течение 1,5 ч образование провоспалительного цитокина интерлейкина IL-1β и противовоспалительного интерлейкина IL-10. Введение апиумана увеличивает концентрацию IL-10 и предохраняет мышей от гибели, обусловленной наличием IL-1β.
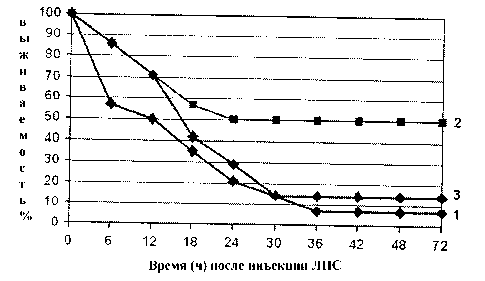
Рис. 3. Влияние апиумана на выживаемость мышей, подвергнутых действию летальной дозы ЛПС (50 мг/кг внутрибрюшинное введение) [16].
Мышам перорально введены за 24 ч до инъекции ЛПС: 1 – PBS (контроль), 2 – апиуман (50 мг/кг) и 3 – яблочный пектин (50 мг/кг)
Кроме того, апиуман снижает число нейтрофилов, мигрирующих в перитонеальную область после инъекции ЛПС. В результате цитокиновый профиль смещается в сторону IL-10 и обусловливает увеличение выживаемости мышей. Эти данные о предотвращении введением апиумана воспаления, индуцированного ЛПС, указывают на высокий противовоспалительный потенциал данного пектина [21].
Таким образом, различия в характере действия на воспаление связаны с тонкими элементами строения пектинов, такими как структура боковых углеводных цепей и взаимное расположение разветвленных областей пектиновой макромолекулы [17-21].
Пектины пищевых растений
В организм человека пектины, играя роль «пищевых волокон», попадают при включении в рацион питания пищевых растений. Большой интерес представляет идея выявления характера изменений, которые происходят с пектинами пищевых растений в условиях желудочно-кишечного тракта и, в первую очередь, в условиях гастральной среды. С этой целью пектиновый полисахарид, названный капсикуманом СА, был получен из сладкого перца Capsicum annuum экстракцией с помощью раствора соляной кислоты (рН 1.5) и пепсина при 37°С в течение 4 ч. Этот экстрагент в определенной степени имитирует условия гастральной среды [22]. Строение капсикумана было установлено и показано, что он является также представителем группы рамногалактуронана RG-I. Его главная углеводная цепь галактуронана содержит метилэтерифициро-ванные остатки D-галактуроновой кислоты, кроме того, в состав макромолекулы входят сложноэфир-ные О-ацетильные группы, локализация которых не установлена. В образце также содержится 5,8% прочно связанного белка и, по существу, капсику-ман представляет собой пектин-белковый комплекс, невысокое содержание протеина обусловлено ферментативным действием пепсина, присутствующего в экстракционной среде, имитирующей гастральные условия. С помощью АСМ показано, что протеин-белковые комплексы представляют собой макромолекулу пектина с белковым компонентом, присоединенным к одному из концов пектиновой углеводной цепи, что имеет место для пектина из сахарной свеклы [23]. Интересно отметить, что белковая составляющая присоединяется только к одному концу пектина, а не к разным участкам макромолекулы. При этом белок не отделяется от пектина хроматографией даже в присутствии 4М NaCl, что может указывать на наличие скорее ковалентной связи, а не механического комплекса пектина с белком. Конечно, необходимо дополнительное исследование, чтобы строго определить тип связи между пектином и белком в этом агрегате и охарактеризовать углеводные цепи в пектине сахарной свеклы. Выделение и характеристика белкового компонента необходимы для выяснения его роли в клеточной стенке растения. По аналогии, можно сказать, что подобный факт имеет место и в случае пектин-белкового комплекса капсикумана [22].
При частичном гидролизе капсикумана (СА) был получен галактуронан СА-Н, который представляет главную углеводную цепь пектина [22].
При определении противовоспалительной активности СА и СА-Н изучено изменение в продуцировании провоспалительного цитокина – фактора некроза опухоли TNF α и противовоспалительного цитокина IL-10 после стимуляции введением ЛПС. Показано, что капсикуман СА снижает уровень TNF α спустя 24 ч. после перорального введения мышам в дозах 40-100 мг/кг. Продуцирование IL-10 увеличивается у мышей, которые получили СА за 36 и 48 ч до стимуляции ЛПС. При помощи модели эндотоксинового шока показано [22], что СА увеличивает выживаемость мышей, обработанных летальной дозой ЛПС. СА-Н также снижает продукцию TNF-α и увеличивает выживаемость мышей в ответ на введение ЛПС (рис. 4).
Таким образом, капсикуман СА и его галакту-ронан СА-Н обладают противовоспалительным действием.
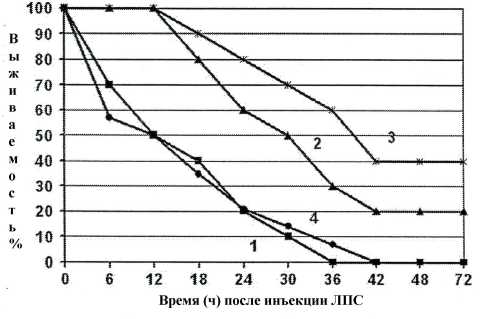
Рис. 4. Влияние капсикумана СА и его галактуро-нана СА-Н на выживаемость мышей, подвергнутых действию летальной дозы ЛПС (50 мг/кг i.p .) [17]. Мышам перорально введены за 24 ч до инъекции ЛПС: 1 – PBS (контроль), 2 – СА (50 мг/кг), 3 – СА-Н (50 мг/кг) и 4 – яблочный пектин (50 мг/кг).
Большой интерес представляют работы группы норвежских ученых [24-29], их работы в конце прошлого столетия (1995-2000 гг.) посвящены детальному изучению пектиновых веществ подорожника Plantago major L., были исследованы пектиновые полисахариды и сопутствующие им биологически активные арабиногалактаны [24-26].
В последней серии их работ [27-29] изучен пектин из пищевого растения, в частности, белокочанной капусты Brassica oleracea , которая является важным овощным продуктом в рационе скандинавов и широко культивируется. Кроме пищевого аспекта, капуста широко применяется для лечения пролежней и заживления ран. Механизм этого не совсем ясен, но вполне возможно, что это действие обусловлено пектиновыми веществами как стимуляторами иммунной системы.
Используя фрагментацию с помощью мета-нолиза с последующей спектроскопией ЯМР полученных соединений, авторы показали [28], что пектин белокочанной капусты, как обычно, имеет кор галактуронана, состоящего из 1,4-связанных остатков α-D-галактопиранозилуроновой кислоты с высокой степенью метилэтерифицирования (СМ) и небольшим числом 1,3-связанных боковых цепей. Кроме того, пектин содержит большое число остатков α-L-арабинофуранозы, D-галактопиранозы и незначительное количество остатков L-рамнозы и D-ксилозы в пиранозной форме. Своеобразием данного пектина является наличие фрагмента кси-логалактуронана, в котором присутствуют терминальные и 1,4-связанные остатки D-ксилозы. Остальные боковые цепи макромолекулы пектина состоят из терминальных и 1,5-связанных остатков α-L-арабинофуранозы, а также терминальных; 1,3-; 1,6 и 1,3,6-связанных остатков β-D-галактопирано-зы. Боковые цепи присоединены 1,4-связями к остаткам рамнозы основной цепи линейного рамнога-лактуронана, в которую остатки α-L-рамнозы включены 1,2-гликозидными связями [28]. Полученные результаты свидетельствуют о том, что пектин капусты имеет большое структурное сходство с другими пектинами типа RG-I, его своеобразием является наличие фрагмента ксилогалактуронана [28]. Он так же, как и другие пектины, обладает иммуно- логическими свойствами. В частности, данный пектин способен связывать комплемент, эта способность усиливается при гидролитическом отщеплении остатков арабинофуранозы из боковых цепей макромолекулы. Молекулярная масса пектина варьирует в широких пределах от 10 до 150 кДа и зависит от способа выделения [29]. Были использованы два различных метода выделения пектина из белокочанной капусты, которые привели к получению образцов с различными характеристиками. Экстракция водой создает слабокислые условия, в которых экстрагируются только водорастворимые пектиновые фракции. В этих условиях частично отщепляются боковые цепи, что снижает физиологическую активность полученного пектинового образца [29], и это следует принимать во внимание при выборе метода экстракции. Второй метод представляет собой последовательную экстракцию водными ионными растворами, которые приводят к извлечению не только водорастворимых фракций, но также и пектиновых фрагментов протопектина. Этот метод включает экстракцию водой, подкисленной соляной кислотой до рН 4.5, затем последовательно 0.05 М оксалатом аммония, двукратно 0.05 М раствором соды Na2CO3 и, наконец, 0.05 М раствором соляной кислоты при 85°С и 100°С. В этом случае получаются образцы пектина с большей молекулярной массой и с более высокой СМ остатков D-галактуроновой кислоты в углеводной цепи галактуронана. Эти остатки легко подвергаются β-элиминации горячими, нейтральными водными растворами. Полученные пектиновые фракции обладают способностью связывать комплемент, особенно после предварительной обработки полигалактуроназой [29].
Синтез и свойства сульфатированных полисахаридов
Хорошо известно, что сульфатированные полисахариды обладают рядом новых видов физиологической активности, в частности, обнаруживают высокую антикоагулянтную активность [30] и другие виды физиологической активности, такие как противовирусная, антипролиферативная, антисклеротическая, иммуномодулирующая и ряд других [3034]. Так, было найдено, что сульфаты лентинана и полисахаридов Astragalus sp. существенно повышают иммунный ответ на вакцину к болезни Ньюка-стла и могут быть рекомендованы как новый тип иммуноадъювантов [31].
Хронологически первым антикоагулянтом стал открытый в начале XX в. гепарин, представляющий собой природный линейный сульфатированный глюкозаминогликан, который вырабатывается тучными клетками ряда животных [30]. Макромолекула гепарина состоит из чередующихся 1.4-связанных остатков α-D-глюкозамина и 2-О-сульфо-α-L-идуроновой или β-D-глюкуроновой кислоты. Остатки глюкозамина содержат сульфатные группы по 6- и реже по 3-гидроксильным группам, аминогруппа также сульфатирована (6-О- или 3-О-суль-фо- и N-сульфо-гепарин), антикоагулянтная и анти-тромбическая активности гепарина связаны с ингибированием сериновых протеиназ, включенных в каскад реакций свертывания крови, и проявляются только в присутствии антитромбина, с которым гепарин образует комплекс [30]. Благодаря высокой эффективности гепарин широко используется в медицине для предотвращения тромбообразования и предупреждения тромбозов, являющихся важнейшей причиной инфарктов и инсультов, а также феномена внезапной смерти.
К сожалению, применение гепарина сопровождается и рядом нежелательных побочных эффектов, таких как остеопороз, тромбоцитопения, геморрагии и др. В качестве замены были предложены препараты с подобной активностью, в частности, хондроитин сульфат, дерматан сульфат, сульфаты 1,3-L-галактанов морских беспозвоночных, сульфатированные фуканы водорослей и т.д. [30].
В качестве альтернативы нами сделана попытка синтеза сульфатированных пектинов и изучена их антикоагулянтная активность [35]. До настоящего времени не было данных о влиянии введения сульфатных групп в макромолекулу пектинов на их физиологическую активность и антикоагулянтные свойства. Нами проведено сульфатирование следующих пектиновых полисахаридов: бергенана, лемнана, а также галактуронана, полученного в результате частичного кислотного гидролиза бергенана.
Получение сульфатов полисахаридов достигается нуклеофильным замещением гидроксильных групп [30]. Сульфатирование вышеназванных пектинов проводили с использованием в качестве сульфатирующих агентов монометилсульфата пиридина, пиридинсульфотриоксида и хлорсульфоновой кислоты [35]. В результате было найдено, что оптимальным реагентом является хлорсульфоновая кислота. Увеличение количества сульфатирующего реагента до трехкратного избытка приводит к увеличению степени сульфатирования (в случае галактуронана до 15 моль –SO 3 ¯ на 10 моль остатков галактуроновой кислоты), сопровождается частичной деструкцией углеводной цепи (до 30%) и образованием фрагментов с мол. массой ниже 50 кДа. При сульфатировании лемнана и бергенана наблюдается частичный гидролиз боковых цепей, но образуются полисахариды, которые содержат сульфатные группы. Полученные сульфатированные производные пектиновых фрагментов удлиняют время свертывания плазмы крови человека по сравнению с нефракционированным гепарином, выбранным в качестве положительного стандарта, и с исходными пектинами. Их антикоагулянтное действие связано с ингибированием фибриногенсвертываю-щей и амидолитической активности тромбина и фактора Ха каскада реакций свертывания крови [35].
Биотехнология получения пектиновых веществ
Многие исследования структуры и биосинтеза растительных полисахаридов проводятся на клеточных культурах, которые способны продуцировать полисахариды, входящие в состав клеточных стенок, в частности, пектиновые вещества: пектины и сопутствующие им нейтральные полисахариды, в основном арабиногалактаны, содержащие небольшой процент остатков D-галактуро-новой кислоты, которые придают им кислый характер [1]. Нами были разработаны методы получения каллусных культур следующих растений: смолевки обыкно- венной Silene vulgaris и смолевки татарской Silene tatarica, пижмы обыкновенной Tanacetum vulgare, ряски малой Lemna minor и шиповника морщинистого Rosа rugosa, – с дальнейшим выделением из них пектиновых веществ, продуцируемых данными каллусными культурами [1].
Для выделения пектинов из каллусной ткани использован стандартный метод последовательной экстракции растительного материала водой и 0,7%-ным водным оксалатом аммония при 68°С после предварительного гидролиза протопектина разбавленным раствором соляной кислоты (рН 4.0; 50°С). При этом найдено, что клеточные стенки каллуса смолевки обыкновенной содержат 2-12% пектинов от сухой биомассы, смолевки татарской – 3-4, пижмы обыкновенной – 5-7, ряски малой – 2-4 [1] и шиповника морщинистого – 7,4% [36].
Структурное изучение пектинов, выделенных из каллусных культур, и сравнение их с пектинами нативных растений дали следующие результаты. Силенан SVC из каллуса и силенан SV из интактного растения близки по структуре и свойствам [37], то же самое имеет место для танацетана TV и TVC [38], а также для розолинанов, выделенных из мякоти плодов и из каллуса шиповника морщинистого [36]. Все они относятся к группе рамногалактурона-на RG-I, являются типичными его представителями, имеют много общего в структуре, но различаются по характеру строения боковых цепей [1]. Установлено, что данные пектины являются иммуномодуляторами, а их галактуронановые фрагменты обладают противовоспалительным действием. Розоли-наны из нативного растения и из каллусной культуры обладают, кроме того, гиполипидемической активностью и влияют на адгезивную способность перитонеальных макрофагов мышей [36].
Лемнаны LM и LMC [37, 38] из нативного растения и из каллуса, соответственно, отличаются тем, что LMC содержит значительно меньшее количество остатков D-апиозы. Однако получены линии каллуса, продуцирующие лемнан LMC с высоким содержанием остатков нейтральных моносахаридов, включая D-апиозу [39].
Клеточные культуры представляют собой стабильную гомогенную систему, которая продуцирует в стандартных условиях пектиновые вещества с постоянной, вполне определенной структурой, что позволяет использовать их в качестве модельных соединений для изучения элементов тонкого строения, метаболизма и биосинтеза, для выяснения механизмов регуляции этих процессов для исследования взаимосвязи структуры и физиологической активности [1].
Данные свойства клеточных культур привели к идее получения с их помощью пектиновых полисахаридов с заранее заданной структурой и свойствами. В этой связи было изучено действие ультрафиолетового излучения на рост каллусной культуры, на характер пектиновых веществ, а также на их антиоксидантную активность [40, 41]. Кроме того, исследована модификация пектинов каллусных культур: силенана и лемнана, – при обработке ферментами – карбогидразами [42, 43]. При этом установлено, что при действии ультрафиолетового облучения наблюдается изменение роста каллуса, меняется и полисахаридный состав пектиновых веществ клеточных стенок: силенана и кислого арабиногалактана. Влияние ультрафиолета зависит от дозы облучения, что сопровождается ростом концентрации пектиновых веществ и снижением содержания остатков арабинозы и галактозы в силе-нане и арабиногалактане. Максимальная скорость роста наблюдается при низких дозах облучения, а снижение содержания моносахаридных остатков, напротив, при высоких дозах и продолжительной экспозиции. Следует отметить, что в процессе облучения меняется соотношение остатков арабинозы и галактозы, что позволяет направленно влиять на состав пектиновых веществ, продуцируемых каллусом смолевки, регулируя условия облучения. Учитывая, что пектиновые вещества каллуса, как и нативного растения, обладают выраженной иммуномодулирующей активностью, а также принимая во внимание высокую продуктивность каллусных культур по биомассе и синтезируемым пектиновым веществам, можно с уверенностью сказать, что каллусные культуры смолевки могут использоваться для биотехнологического получения физиологически активных пектиновых веществ [40, 41]. Интересно отметить, что полученный таким путем силе-нан обладает антиоксидантной активностью, которая усиливается при ультрафиолетовом облучении более чем в два раза [41].
При действии на каллусную культуру смолевки различными карбогидразами, в частности, пектиназой (полигалактуроназой) наблюдается активация внутриклеточных ферментов: α-L-арабино-фуранозидазы и β-D-галактопиранозидазы, – что приводит к уменьшению числа остатков нейтральных моносахаридов в силенане и к снижению его мол. массы. В результате культивирования каллуса на средах с высокой концентрацией пектиназы получают пектиновые вещества с низким содержанием арабинозы [42]. При действии на каллусные культуры смолевки и ряски β-D-галактозидазы [42, 43] образуются пектиновые вещества с низким содержанием остатков арабинозы и галактозы, при этом наибольшие изменения наблюдаются в высокомолекулярных полисахаридных фракциях. Таким образом, культивирование клеточных культур растений в присутствии карбогидраз или при облучении ультрафиолетом может быть использовано как инструмент модификации структуры и физиологической активности пектиновых веществ. Как следствие, клеточные культуры могут рассматриваться как альтернативные источники получения стандартизированных физиологически активных пектиновых полисахаридов с регуляцией их строения и свойств.
Кроме того, установлено, что пектины как нативных растений, так и каллусных культур оказывают стимулирующее действие на рост и развитие овощных и зерновых культур и способствуют заметному увеличению их урожайности [44].
Заключение
Пектиновые вещества привлекают все большее внимание исследователей, благодаря своеобразному строению, уникальным биологическим функциям в растениях и широкому спектру физиологической активности [1, 2]. Это полипотентные биополимеры, биологические функции и все виды физиологической активности которых тесно связаны с изменяющимся строением при переходе от растения к растению, от периода вегетации и от ряда других факторов, обусловленных средой обитания растения или условиями получения клеточных культур.
Изучение связи между структурой и активностью выявило уникальное свойство пектинов – поли-потентность их структуры в отношении действия на иммунную систему. Пектиновая макромолекула содержит фрагменты, способные снижать или увеличивать иммуномодулирующее действие. Использование с этой целью фрагментов главной углеводной цепи и разветвленных областей пектина представляется перспективной задачей будущих исследований.
Насущной проблемой является выявление новых структурных типов пектинов из новых природных источников, установление их строения, в том числе выяснение элементов тонкой структуры, включая характер связи с белком, присутствующим в составе пектиновых веществ. Чрезвычайно важны – разработка новых методов исследования структуры и свойств пектиновых веществ, изучение путей их метаболизма в организме человека и животных, а также путей биосинтеза в растениях [45].
Работа выполнена при финансовой поддержке грантов РФФИ, Программы РАН «Молекулярная и клеточная биология», Программы «Ведущие научные школы», интеграционных проектов совместных исследований УрО РАН с ДВО РАН и СО РАН, гранта Минобрнауки РФ 02.512.11.2190.
Авторы выражают глубокую признательность ст. инженеру Института физиологии Коми НЦ УрО РАН Ю.А.Овчинниковой за большую помощь в подготовке и оформлении данного обзора.
Список литературы Новейшие сведения о пектиновых полисахаридах
- Оводов Ю.С., Головченко В.В., Гюнтер Е.А., Попов С.В. Пектиновые вещества растений европейского Севера России. Екатеринбург, 2009. 111 с.
- Оводов Ю.С. Современные представления о пектиновых веществах//Биоорган. химия, 2009. Т. 35, №3. С. 293-310.
- Round A.N., MacDougal A.J., Ring S.G., Morris V.J. Unexpected branching in pectin observed by atomic force microscopy//Carbohydr. Res., 1997. Vol. 303. P. 251-253.
- Оводова Р.Г., Попов С.В., Бушнева О.А. и др. Разветвление галактуронанового кора макромолекулы комарумана, пектина сабельника болотного Comarum palustre L.//Биохимия, 2006. Т. 71. №5. С. 666-671.
- Round A.N., Rigby N.M., MacDougal A.J., Morris V.J. A new view of pectin structure revealed by acid hydrolysis and atomic force microscopy//Carbohydr. Res., 2010. Vol. 345. P. 487-497.
- Vincken J.-P., Schols H.A., Oomen R.J.F.J. et al. If Homogalacturonan were a side chain of rhamnogalacturonan I. Implication for cell wall architecture//Plant Physiol., 2003. Vol. 132. P. 1781-1789.
- Kirby A.R., MacDougal A.J., Morris V.J. Atomic force microscopy of tomato and sugar beet pectin molecules//Carbohydr. Polym., 2008. Vol. 71. P. 640-647.
- Оводов Ю.С. Биологически активные пектиновые полисахариды растений Республики Коми//Север: наука и перспективы инновационного развития/Отв. ред. чл.-корр. РАН В.Н.Лаженцев. Сыктывкар, 2006. С. 236-255; 374-395.
- Оводов Ю.С., Оводова Р.Г., Попов С.В. Биогликаны-имммуномодуляторы. Строение и свойства//Фитотерапия, биологически активные вещества естественного происхождения. ЗАО «Биопрогресс». Черноголовка, 2004. С. 348-363.
- Gloaguen V., Brudieux V., Closs B. et al. Structural characterization and cytotoxic properties of an apiose-rich pectic polysaccharide obtained from the cell wall of the marine phanerogam Zostera marina//J. Nat. Prod., 2010. Vol. 73. P. 1087-1092.
- Golovchenko V.V., Ovodova R.G., Shashkov A.S., Ovodov Yu.S. Structural studies of the pectic polysaccharide from duckweed Lemna minor L.//Phytochemistry, 2002. Vol. 60. P. 89-97.
- Хасина Э.И., Сгребнева М.Н., Оводова Р.Г. и др. Гастропротективное действие лемнана -пектинового полисахарида, выделенного из ряски малой Lemna minor L.//Доклады АН, 2003. Т. 390. №3. С. 413-415.
- Khasina E.I., Sgrebneva M.N., Ovodova R.G. et al. Gastroprotective effect of lemnan, a pectic polysaccharide from Lemna minor L.//Phytopharmacology and therapeutic values, Eds. J.N.Govil, V.K.Singh, S.K.Mishra, Studium Press LLС, Houston, USA. Recent Progress in Medical Plants, 2008. Vol. 20. P. 181-188.
- Попов С.В., Оводова Р.Г., Попова Г.Ю. и др. Ингибирующее действие пектиновых галактуронанов на адгезию нейтрофилов//Биоорган. химия, 2007. Т. 33. №1. С. 187-192.
- Popov S.V., Ovodova R.G., Ovodov Yu.S. Effect of lemnan, pectin from Lemna minor L. and its fragments on inflammatory reaction//Phytother. Res., 2006. Vol. 20. №5. P. 403-407.
- Попов С.В. Взаимодействие фагоцитов млекопитающих с полисахаридами растений. Сыктывкар, 2002. 97 с.
- Попов С.В., Оводова Р.Г., Попова Г.Ю. и др. Ингибирующее действие пектиновых галактуронанов на адгезию нейтрофилов//Биоорган. химия, 2007. Т.33. №1. С. 187-192.
- Головченко В.В., Бушнева О.А., Оводова Р.Г. и др. Структурное исследование бергенана, пектина из бадана толстолистного Bergenia crassifolia//Биоорган. химия, 2007. Т. 31. №1. С. 54-63.
- Popov S.V., Ovodova R.G., Markov P.A. et al. Protective effect of comaruman, pectin from cinquefoil Comarum palustre L. on acetic acidinduced colitis in mice//Digest. Dis. Sci., 2006. Vol. 51. №9. P. 1532-1537.
- Марков П.А., Попов С.В., Никитина И.Р., Оводова Р.Г. и др. Противовоспалительная активность пектинов и их галактуронанового кора//Химия раст. сырья, 2010. №1. С. 21-26.
- Ovodova R.G., Golovchenko V.V., Popov S.V. et al. Chemical composition and anti-inflammatory activity of pectic polysaccharide isolated from celery stalks//Food. Chem., 2009. Vol. 114. P. 610-615.
- Popov S.V., Ovodova R.G., Golovchenko V.V. et al. Chemical composition and anti-inflammatory activity of pectic polysaccharide isolated from sweet pepper using a simulated gastric medium//Food. Chem., 2011. Vol. 124. P. 309-315.
- Kirby A.R., MacDougal A.J. Sugar beet pectinprotein complexes//Food. Biophysics (FOBI), 2006. Vol. 1. P. 51-56.
- Samuelsen A.B., Paulsen B.S., Wold J.K. et. al. Isolation and partial characterization of biologically active pectin from Plantago major L.//Carbohydr. Polym., 1996. Vol. 30. P. 37-44.
- Samuelsen A.B., Paulsen B.S., Wold J.K. et al. Characterization of a biologically active arabinogalactan from the leaves Plantago major L.//Carbohydr. Polym., 1998. Vol. 35. P. 145-153.
- Samuelsen A.B. Polysaccharides in Plantago major L. Studies of structural and biological activity. Ph.D.Thesis.Univ. Oslo, 1998.
- Westereng B., Yousif O., Michaelsen T.E., Samuelsen A.B. et al. Pectin isolated from white cabbage -structure and complementfixing activity//Mol. Nutr. Food. Res., 2006. Vol. 50. P. 746-756.
- Samuelsen A.B., Westereng B., Yousif O. et al. Structural features and complement-fixing activity of pectin from three Brassica oleracea varieties: white cabbage, kale, and red kale//Biomacromolecules, 2007. Vol. 8. P. 644-649.
- Westereng B., Michaelsen T.E., Samuelsen A.B.,Knutsen S.H. // Effect of extraction conditions on the chemical structure and biological activity of white cabbage pectin // Carbohydr. Polym., 2008. Vol. 72. P. 32-42.
- Местечкина Н.М., Щербухин В.Д. Сульфаты полисахаридов и их антикоагулянтная активность//Прикл. биохим. микробиол., 2010. Т. 46. №3. С. 291-298.
- Ma X., Guo Z., Wang D. et al. Effect of sulfated polysaccharides and their prescriptions on immune response of ND vaccine in chicken//Carbohydr. Polym., 2010. Vol. 82. P. 9-13.
- Huang X., Hu Y., Zhao X. et al. Sulfated modification can enhance the adjuvanicity activity of astragalus polysaccharide for Newcastle disease vaccine//Carbohydr. Polym., 2008. Vol. 73. P. 303-308.
- Huang X., Wang D., Hu Y. et al. Effect of sulfated astragalus polysaccharide on cellular infectivity of infections bursal disease virus//Intern. J.Biol. Macromol., 2008. Vol.3. P.166-171.
- Guo Z., Hu Y., Wang G. et al. Sulfated modification can enhance the adjuvanicity of lentinan and improve the immune effect of ND Vaccine//Vaccine, 2009. Vol. 27. P. 660-665.
- Витязев Ф.В., Головченко В.В., Патова О.А. и др. Синтез сульфатированных пектинов и их антикоагулянтная активность//Биохимия, 2010. Т. 75. Вып. 6. С. 857-867.
- Злобин А.А., Жуков Н.А., Оводова Р.Г. Химическая характеристика водорастворимых полисахаридов каллусной ткани шиповника морщинистого Rosa rugosa Thunb//Химия раст. сырья, 2008. №1. С. 51-55.
- Бушнева О.А., Оводова Р.Г., Шашков А.С. и др. Структурное исследование арабиногалактана и пектина из каллуса Silene vulgaris (M.) G.//Биохимия, 2006. Т. 71. Вып. 6. С. 798-807.
- Гюнтер Е.А. Культуры клеток нетрадиционных растений как продуценты полисахаридов//Аграрная Россия, 2001. №6. С. 73-74.
- Гюнтер Е.А., Попейко О.В., Оводов Ю.С. Продуцирование полисахаридов каллусными культурами ряски малой//Прикл. биохим. микробиол., 2008. Т. 44. №1. С. 117-122.
- Гюнтер Е.А., Оводов Ю.С. Действие ультрафиолетового излучения на рост и полисахаридный состав каллусной культуры Silene vulgaris//Прикл. биохим. микробиол., 2007. Т. 43. №4. С. 518-526.
- Гюнтер Е.А., Борисенков М.Ф., Оводов Ю.С. Действие ультрафиолета на строение и антиоксидантную активность силенана каллусасмолевки обыкновенной//Прикл. биохим. микробиол., 2009. Т. 45. №4. С. 470-475.
- Гюнтер Е.А., Попейко О.В., Оводов Ю.С. Модификация полисахаридов каллусной культуры Silene vulgaris (M.) G. c помощью карбогидраз in vitro//Биохимия, 2007. Т. 72. Вып. 9. С. 1238-1247.
- Günter E.A., Popeyko O.V., Ovodov Yu.S. Action of -galactosidase in medium on the Lemna minor L. callus polysaccharides//Carbohydr. Res., 2009. Vol. 344. P. 2602-2605.
- Елькина Е.А., Оводов Ю.С. Пектиновые полисахариды как регуляторы роста зерновых культур//Тезисы докладов IV Всероссийской конференции «Химия и технология растительных веществ». Сыктывкар, 2006. С. 245.
- Harholt J., Suttangkakul A., Scheller H.V. Biosynthesis of pectin//Plant Physiol., 2010. Vol. 153. P. 384-395.
