О происхождении носителей охотской культуры Северного и Восточного Хоккайдо (по краниологическим данным)
")
Автор: Моисеев В.г
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология
Статья в выпуске: 1 (33), 2008 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14522632
IDR: 14522632 | УДК: 572
Текст обзорной статьи О происхождении носителей охотской культуры Северного и Восточного Хоккайдо (по краниологическим данным)
Проблема происхождения носителей охотской культуры I тыс. н.э. с момента ее выделения в 1930-х гг. остается одной из наиболее актуальных в популяционной истории Дальнего Востока России и Японского архипелага. Изолированность данной культуры от цивилизаций того времени ограничивает роль письменных источников при выяснении ее генезиса, вследствие чего ключевую роль приобретают археологические и палеоантропологические данные.
Согласно археологическим источникам, охотская культура сложилась в V в. н.э. на основе сусуйской в южной части Сахалина. Последняя, по мнению А.А. Василевского [2002], родственна северо-сахалинской культуре, но восприняла некоторые черты культуры эпидзёмон, носители которой неоднократно проникали на Сахалин с Хоккайдо. Исследователь подчеркивает культурную неоднородность охотцев, полагая, что охотская культура является секвенцией родственных локальных археологических культур, сосуществовавших во времени и пространстве. Несомненным представляется культурное воздействие на охотское население материковых культур, связанных с тунгусо-маньчжурскими племенами (культура мохэ).
Сразу после формирования охотской культуры, а именно с V в. н.э., начиналась ее экспансия на территорию Хоккайдо, где она довольно быстро распространилась по узкой береговой линии Охотского моря. Большинство охотских памятников расположено на восточном и северном побережье острова и на Курильских островах (рис. 1); лишь немногие зафиксированы
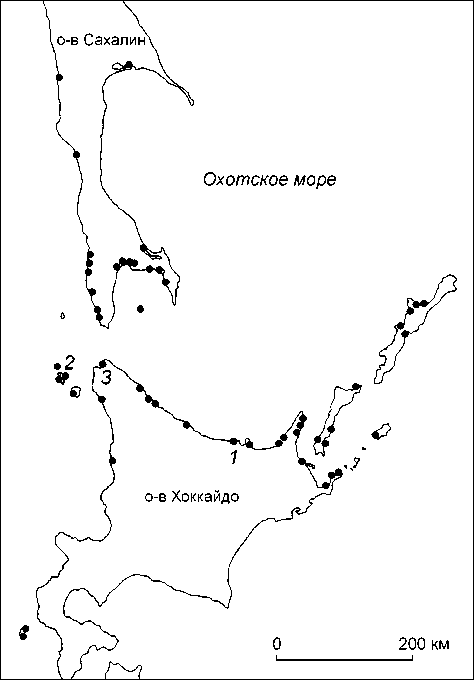
Рис. 1. Расположение главных памятников охотской культуры. Памятники, краниологические серии которых использованы в анализе:
1 – Мойоро; 2 – Хаманака; 3 – Омисаки.
Археология, этнография и антропология Евразии 1 (33) 2008 © В.Г. Моисеев, 2008
в более южных районах Хоккайдо [Matsumura et al., 2006] Такая локализация охотских памятников объясняется высоким уровнем специализации экономики ее носителей, практиковавших охоту на крупных морских млекопитающих. На ведущую роль такого направления специализации охотничьей базы носителей охотской культуры, кроме археологических данных, указывают и результаты изотопного анализа [Yoneda, 2006].
Предполагается, что судьбы локальных групп охот-цев Хоккайдо были различны. Среди японских археологов распространено мнение о большей аккультурации восточных групп охотцев, по сравнению с северными местным протоайнским населением, представляющим культуру сацумон. Результатом аккультурации стала культура тобинатай, бытовавшая на востоке Хоккайдо. В северных же районах острова охотцев вытесняли на Сахалин протоайны [Ohyi, 1975].
Антропологами было высказано несколько точек зрения по поводу происхождения и генетических связей носителей охотской культуры. Среди возможных «родственников» охотцев назывались эскимосы [Suzuki, 1958] и алеуты [Кодама, 1948; Ито, 1951]. В настоящее время, пожалуй, наиболее распространено мнение об их родстве с современными амурскими народами, а также нивхами [Ямагучи, 1975, 1981; Ishida, 1988, 1994; Ishida et al., 1994; Kozintsev, 1990, 1992б]. Отмечалась также возможность метисации охотцев с протоайнским населением Хоккайдо [Ito, 1965, 1971; Kozintsev, 1990, 1992б].
Одним из главных препятствий на пути изучения антропологических особенностей носителей охотской культуры являлся недостаток краниологического материала. Лишь в последние годы после тщательной обработки и регистрации антропологических коллекций Университета Хоккайдо (г. Саппоро) появилась возможность исследовать, пожалуй, наиболее представительную краниологическую серию – из Мойоро (окрестности г. Абасири), а также менее многочисленные серии – из Хаманака (о-в Ребун) и Омисаки (окрестности г. Вакканай). Прежние исследования этих материалов были проведены по довольно короткой краниометрической программе [Кодама, 1948; Ито, 1951; Ito, 1965], что обусловило ограниченность воз- можностей использования полученных результатов. Разработанные в последние годы новые эффективные системы признаков, а также новые методы статистической обработки материала заставляют вновь обратиться к данным коллекциям.
В настоящее время становится все более очевидным, что различные системы антропологических признаков не являются одинаково информативными при изучении разных этапов популяционной истории. Мне пришлось убедиться в этом при анализе антропологических материалов из Западной и Южной Сибири, а также из Восточной Европы [Моисеев, 2001, 2004]. Выяснилось, что каждая система признаков «рассказывает» свою собственную историю. Если признаки в пределах систем информативны, то любая из этих историй в общем достоверна, хотя и не достаточна. Иными словами, нет «идеальной» системы признаков, которая бы в полной мере отражала историю той или иной человеческой популяции. Разнобой в информации, получаемой по разным системам, можно свести к минимуму, если интегрировать данные различных систем. Преимущества такого подхода были продемонстрированы в ряде работ [Kozintsev, Gromov, Moiseyev, 1999; Козинцев, 2004; Моисеев, 2001, 2004].
Цель настоящей статьи – выявить родственные связи локальных групп носителей охотской культуры Хоккайдо на основе интеграции данных по двум системам антропологических признаков, наиболее часто используемым при исследовании палеоантропологического материала, – краниометрии и краниоскопии.
Программа, материал и методика
Черепа представителей охотской культуры, а также айнов и носителей культуры дзёмон о-ва Хоккайдо, хранящиеся в Университете Хоккайдо и Медицинском колледже Саппоро, изучались мной по краниометрической программе, а также по краниоскопической программе А.Г. Козинцева [Козинцев, 1988; Kozintsev, 1992а]. Для интеграционного межгруппового анализа были отобраны 14 метрических и пять неметрических показателей (табл. 1, 2). Из краниоскопической
Таблица 1. Частоты краниоскопических признаков в охотских группах
|
Группа |
ЗИ |
КВШ |
ЗСШ |
ПГУ II |
ИПНШ |
НО |
|
Мойоро |
64,1 (39) * |
14,3 (96) |
42,4 (66) |
46,2 (39) |
63,7 (110) |
32,7 (107) |
|
Хаманака |
86,2 (31) * |
22,8 (47) |
54,8 (31) |
73,3 (15) |
83,3 (30) |
38,5 (39) |
|
Омисаки |
82,8 (26) * |
22,0 (45) |
28,6 (35) |
73,1 (26) |
57,9 (38) |
26,5 (49) |
Примечание . ЗИ – затылочный индекс, КВШ – клиновидно-верхнечелюстной шов, ЗСШ – заднескуловой шов, ПГУ II – подглазничный узор типа II, ИПНШ – индекс поперечного небного шва, НО – надглазничные отверстия. В скобках указано число наблюдений.
*Частота, высчитанная по уравнению регрессии.
Таблица 2. Средние величины краниометрических признаков в охотских группах
|
Признак |
Мойоро |
Хаманака |
Омисаки |
|||
|
N |
X |
N |
X |
N |
X |
|
|
1. Продольный диаметр |
29 |
183,8 |
11 |
183,1 |
14 |
187,3 |
|
8. Поперечный диаметр |
21 |
143,3 |
10 |
143,2 |
13 |
144,3 |
|
17. Высотный диаметр |
17 |
138,2 |
7 |
136,3 |
10 |
141,5 |
|
45. Скуловой диаметр |
9 |
143,9 |
6 |
142,6 |
6 |
144,5 |
|
9. Наименьшая ширина лба |
31 |
92,5 |
11 |
94,6 |
12 |
93,8 |
|
77. Назомалярный угол |
17 |
148,1 |
10 |
149,7 |
10 |
146,7 |
|
Zm. Зиго-максиллярный угол |
9 |
134,9 |
8 |
138,6 |
9 |
135,0 |
|
SS:SC. Симотический указатель |
22 |
34,5 |
9 |
38,3 |
7 |
40,6 |
|
48. Верхняя высота лица |
19 |
77,7 |
8 |
78,0 |
9 |
79,3 |
|
55. Высота носа |
21 |
54,0 |
10 |
54,8 |
9 |
56,3 |
|
54. Ширина носа |
23 |
25,1 |
9 |
26,3 |
7 |
25,2 |
|
51. Ширина орбиты от mf |
18 |
43,3 |
9 |
42,4 |
9 |
44,1 |
|
52. Высота орбиты |
15 |
36,2 |
8 |
36,8 |
9 |
36,3 |
|
75(1). Угол выступания носа |
9 |
18,6 |
6 |
20,3 |
6 |
25,0 |
Примечание . N – число наблюдений, X – средняя величина.
программы на этом этапе был исключен затылочный индекс, т.к. данный признак, эффективно разграничивая европейские и азиатские популяции, не имеет четко выраженного градиента на территории Азии. В случае с краниометрическими признаками
Таблица 3 . Материалы, послужившие основой исследования
Носители охотской культуры представлены тремя сериями. Две из них (Омисаки и Хаманака) относятся к северной группе охотских памятников, одна (Мойо-ро) – к восточной (см. рис. 1). Для сравнительного статистического анализа использовались 12 близких к современности серий с территории Дальнего Востока, а также сборная серия представителей культуры дзё-мон с Японского архипелага (табл. 3).
Статистический анализ состоял из двух этапов. На первом этапе краниометрические и краниоско-пические (после преобразования в радианы) данные были обработаны порознь при помощи канонического анализа и метода главных компонент соответственно. Полученные в результате этих двух анализов векторы имеют принципиально сходные свойства и могут быть использованы как новые признаки, основанные на межгрупповой корреляции исходных показателей.
На втором этапе из полученных канонических векторов (КВ) и главных компонент (ГК) было взято по нескольку первых векторов, в сумме отражающих не менее 70 % общей изменчиво сти по каждой из систем. В данном случае этому критерию отвечают три первых КВ и две первые ГК. Эти вектора были снова обработаны методом главных компонент. Поскольку и ГК, и КВ ортогональны, полученные новые интегральные ГК (ИГК) основаны только на межсистемных корреляциях.
Результаты и обсуждение
Два первых вектора в пределах каждой из систем – I КВ и I ГК (46 и 50 % общей дисперсии соответственно) – описывают принципиально сходное направление межпопуляционной дифференциации. Коэффициент корреляции между ними до статочно высок и со ставляет 0,77 при p < 0,05 (табл. 4). Оба вектора отделяют дзё-монцев и обе группы айнов от остальных групп. Следует отметить, что в случае с I ГК наблюдается небольшое перекрывание значений у айнов и охотцев (рис. 2). Отчетливое антропологическое своеобразие дзёмонцев и сходство их с айнами, выявленное данным анализом, является вполне предсказуемым результатом, который еще раз подтверждает установленный ранее факт генетической преемственности между этими группами (см., напр.: [Kozintsev, 1990]). Комбинация измерительных признаков, характерная для дзёмонцев и айнов, включает большие значения наименьшей ширины лба и угла выступания носа, относительно высокое переносье, а также небольшие величины зигомаксиллярного угла, высоты лица и носа. Краниоскопичес-ки эти группы характеризуются высокой частотой ЗСШ, ИПНШ, а также низкой частотой НО и ПГУ II.
По расположению в пределах I ГК и I КВ охотские группы не сколько различаются. Если по краниоскопическим признакам отмечается сильно выраженное отклонение охотцев от всех монголоидных групп в сторону дзёмонской серии, то по краниометрическим признакам этот сдвиг не столь выражен. В данном случае охотцы не противостоят монголоидам, а занимают промежуточное положение между дзёмонцами и амурскими группами, подобно эскимосам, алеутам и чукчам. В обоих случаях амурские народы, будучи максимально удаленными от дзёмонцев, характеризуются наиболее выраженными монголоидными особенностями. Несмотря на указанные различия, результаты анализа обеих систем признаков в целом не противоречат друг другу.
Для проверки последнего вывода был проведен еще один канонический анализ с привлечением доступных мне индивидуальных краниометрических данных по четырем сериям, измеренным мною, – охотским из Мойоро, Хаманака и Омисаки, а также айнской с Хоккайдо. Анализ проводился по 28 признакам: 1, 8, 17, 5, 45, 43, 46, 40, 9, 77, Zm, SC, SS, 29, 30, 31, 48, 55, 54, 51, 52, 26, 27, 28, 32, 72, 74, 75(1).
Таблица 4. Коэффициенты корреляции между ГК (краниоскопия) и КВ (краниометрия)
|
I ГК |
II ГК |
I КВ |
II КВ |
III КВ |
|
|
I ГК |
1,00 |
0,00 |
0,77 |
0,06 |
0,29 |
|
II ГК |
0,00 |
1,00 |
0,30 |
0,30 |
–0,12 |
|
I КВ |
0,77 * |
0,30 |
1,00 |
0,00 |
0,00 |
|
II КВ |
0,06 |
0,30 |
–0,00 |
1,00 |
0,00 |
|
III КВ |
0,29 |
–0,12 |
–0,00 |
0,00 |
1,00 |
* P < 0,05.
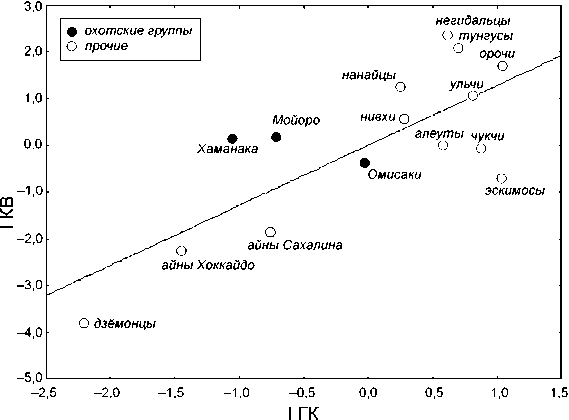
Рис. 2. Расположение групп в пространстве I ГК и I КВ.
При отсутствии того или иного измерения для индивидуума бралась средняя величина по группе.
Области значений I КВ в айнской серии и во всех группах охотцев в значительной степени перекрываются (рис. 3). При этом наибольшее число случаев перекрывания величин I КВ с айнами наблюдается в группе Мойоро (у 17 % индивидуумов данной с ерии). В группах Хаманака и Омисаки такие случаи составляет 16 и 13 % соответственно. Таким образом, наличие протоайнской примеси у охотцев подтверждается. Ее величина по результатам анализа индивидуальных данных в среднем составляла ок. 15 %.
Обращают на себя внимание различия в характере распределения значений I КВ в разных группах охотцев. Так, если индивидуумы из Хаманака, которые попадают в зону трансгрессии с айнами, достаточно сильно отличаются от прочих членов своей группы, то в Мойоро такого разрыва не наблюдается, ареал распределения индивидуумов здесь более компактен. Возможно, это указывает на длительно сть периода метисации охотцев с
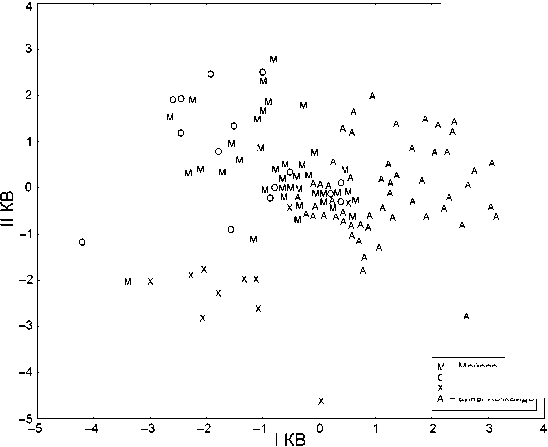
м - мойоро
О - Ом исаки
X -Хаманака
А - айны Хоккайдо
Рис. 3. Расположение индивидуумов в локальных охотских группах и айнской группе с Хоккайдо в пространстве I и II КВ.
Таблица 5. Коэффициенты корреляции между ГК (краниоскопия) и КВ (краниометрия) с первыми тремя ИГК
|
I ИГК |
II ИГК |
III ИГК |
|
|
I ГК |
0,90 |
0,29 |
0,00 |
|
II ГК |
0,34 |
–0,77 |
0,00 |
|
I КВ |
0,91 |
–0,02 |
0,34 |
|
II КВ |
0,18 |
–0,64 |
–0,60 |
|
III КВ |
0,26 |
0,51 |
–0,73 |
|
Собственное число |
1,87 |
1,34 |
1,00 |
|
Процент вариации |
37,4 |
26,8 |
20,0 |
3,0
алеуты
2,5
2,0
1,0
1,5
§ 0,5
0,0
-0,5
-1,0
-1,5
-2,0 L
-3,0
тунгусы
дзёмонцы
-2,5
айны Сахалина
айны Хоккайдо
-2,о
-1,5
-1,0
I ИГК
чукчи эскимосы Q О
нивхи о орочи
О° ,
О негидальцы ульчи нанайцы
Мойоро
Омисаки
Хаманака
0,0
0,5
1,0
1,5
Рис. 4. Расположение групп в пространстве I и II ИГК.
Усл. обозн. см. на рис. 2.
протоайнами в районе Мойоро. Серия из Омисаки в этом отношении более сходна с группой из Мойоро.
Вернемся к результатам основного анализа. Рассмотренные выше I ГК и I КВ – единственные вектора, между которыми наблюдается высокая корреляция (табл. 4). Следовательно, остальные вектора представляют собой относительно независимые направления межпопуляционной дифференциации, что еще раз подтверждает тезис о своеобразии различных систем признаков в плане исторической информации. Для того, чтобы решить, какие их этих направлений наиболее значимы, обратимся к результатам интеграционного анализа.
Как и можно было ожидать, I ИГК (37 % общей изменчивости) в сущности является линейной комбинацией I КВ и I ГК (табл. 5). Она противопоставляет дзёмонскую и айнскую серии прочим (рис. 4). Среди охотских групп наиболее выраженной дзёмонской краниологической спецификой характеризуется группа из Мойоро. Из всех включенных в анализ серий она наиболее близка к айнам. Данный результат соответствует популярной в японской археологии идеи о том, что восточные группы охотцев имели наиболее тесные контакты с протоайнскими представителями культуры сацумон [Ohyi, 1975; Амано, 1979]. Однако айнская специфика в серии из Мойоро все же достаточно умеренна, и в целом группа из Мойоро ближе к остальным охотским группам, чем к айнам. Это свидетельствует о том, что по удельному весу айнский компонент и в данной группе значительно уступает собственно охотскому.
Итак, промежуточный результат интегративного анализа подтверждает мнение ряда исследователей о наличии в составе охот-цев небольшой протоайнской примеси [Ito, 1965, 1971; Kozintsev, 1990, 1992б]. Важно отметить, что единственный смысл I ИГК состоит в противопоставлении дзёмонцев и айнов другим группам. Серии, равноудаленные от названных двух, вовсе не обязательно должны быть родственны между собой. Чтобы выяснить их генетические связи, требуется рассмотреть иные направления дифференциации.
Вторая ИГК (27 % общей изменчивости) определяется в основном II КВ и II ГК (табл. 5). Около четверти ее вариации приходится на одну группу – алеутов. Поскольку проблема происхождения алеутов не входит
в число задач данного исследования, я не буду обсуждать здесь возможные причины их краниологической специфичности. Замечу лишь, что в свете имеющихся генетических данных [Rubicz et al., 2003] полученный результат выглядит неслучайным. Для настоящей работы более важно, что все охотские группы находятся на противоположном от алеутов конце II ИГК. Этот результат не согласуется с мнением о родстве охот-цев с алеутами [Кодама, 1948; Ито, 1951]. Если все же допустить возможность такого родства, то придется предположить, что различия между алеутами и охот-цами были следствием популяционных процессов, имевших место уже после миграции охотцев на Хоккайдо, а это маловероятно. Скорее всего, средневековые предки алеутов не были родственны предкам охотцев.
2,0
1,5
1,0
0,5
^
S 0,0
-0,5
-1,0
-1,5
-2,0
нивхи
нанайцы п тунгусы
айны Хоккайдо
дзёмонцы
айны Сахалина Хаманака ф ульчи негидальцЫ
Мойоро
алеуты
Омисаки
__ __ чукчи эскимосы
О орочи
-3.0 -2,5 -2.0 -1,5 -1,0 -0,5 0,0 0,5 1.0 1,5
I игк
Рис. 5. Расположение групп в пространстве I и III ИГК. Усл. обозн. см. на рис. 2.
Весьма существенной для охотской проблемы оказывается III ИГК (20 % общей дисперсии). Эта ИГК почти полностью определяется краниометрическими данными, причем наибольший вклад вносит III КВ (табл. 5). Третья ИГК противопоставляет арктические группы (эскимосов, чукчей и алеутов) амурским, включая нивхов (рис. 5). Первые характеризуются отрицательными значениями, вторые – положительными. Дзёмонцы и обе айнские серии занимают нейтральное положение (напомню, что их своеобразие проявляется по первым векторам).
Охотцам также соответствуют промежуточные значения III ИГК, однако, в отличие от дзёмонцев и айнов, они не обнаруживают специфичности по иным значимым векторам, что свидетельствует об их метисном происхождении. По III ИГК охотцы неоднородны. Серия из Хаманака характеризуется довольно отчетливым сдвигом в сторону амурских групп, а серия из Омисаки – в сторону арктических. Серия из Мойоро занимает промежуточное положение. «Арктическая» тенденция, обнаруживаемая в группе из Омисаки, подтверждается тем, что данная серия, находясь ниже линии регрессии, проявила сходное с арктическими группами направление межсистемных различий при сравнении I КВ и I ГК (см. рис. 2). Такая же тенденция наблюдается у дзёмонцев и обеих групп айнов. Если это не случайно, то наиболее правдоподобной представляется гипотеза о консервации протоморфных черт у аборигенов Японии, а также у эскимосов и чукчей.
Итак, можно предположить, что локальные группы охотцев Хоккайдо различаются по соотношению амурского и арктического компонентов, чем и может объясняться антропологическая изменчивость в пределах охотского населения. Данное обстоятельство приобретает особое значение в связи с мнением А.А. Василевского о неправомерности использования понятия «охотская культура», поскольку памятники, относимые к ней, принадлежат к нескольким родственным, но вполне самостоятельным культурам [1999]. Комментировать данную точку зрения с позиций физической антропологии преждевременно, поскольку в моем распоряжении практически нет охотского материала с Сахалина. Пока что можно лишь отметить, что краниологические различия между тремя охотскими сериями с Хоккайдо довольно велики и превышают различия между эскимосами и чукчами или между большинством амурских народов (табл. 6).
Тем не менее расстояния между охотскими сериями меньше, чем между ними и любой другой группой, участвующей в анализе, вследствие чего при кластеризации они объединяются в единую группу (рис. 6). Видимо, локальные группы охотцев Хоккайдо имели общее происхождение, а впоследствии различия между ними увеличились в силу изоляции или несовпадения направлений этнических связей. Хотя точно датировать каждое охотское погребение в настоящее время невозможно, представляется маловероятным, что наблюдаемые межгрупповые расхождения вызваны разновременностью групп. Имеющиеся даты для памятников в значительной мере перекрываются, а использование суммарных серий должно нивелировать эффект разновременности, если таковой имел место.
Комментируя результаты кластерного анализа, нельзя не отметить, что не следует, видимо, придавать особое значение объединению охотских групп с чукчами и эскимосами. Данный результат в опреде-
Таблица 6. Евклидовы расстояния между группами
|
Группа |
О о о |
го го го го X |
го о о |
х о 11 84 |
Айны |
о |
с; го С[ S ф т |
ZT >s го го т |
т о о |
X X £Q ZE |
о О о СО |
ф |
|||
|
о С[ го о X |
го X с; го X го О |
||||||||||||||
|
Мойоро |
0,00 |
1,83 |
1,70 |
4,73 |
3,23 |
2,83 |
3,15 |
2,85 |
2,09 |
1,92 |
3,21 |
2,02 |
1,85 |
2,07 |
3,72 |
|
Хаманака |
1,83 |
0,00 |
1,82 |
5,20 |
3,81 |
4,09 |
3,86 |
3,57 |
3,42 |
3,07 |
3,36 |
2,41 |
3,12 |
3,27 |
4,98 |
|
Омисаки |
1,70 |
1,82 |
0,00 |
5,20 |
4,07 |
3,67 |
3,99 |
3,83 |
3,29 |
2,79 |
3,78 |
2,79 |
1,86 |
2,02 |
4,42 |
|
Носители культуры дзёмон |
4,73 |
5,20 |
5,20 |
0,00 |
2,21 |
2,67 |
6,66 |
6,80 |
6,02 |
5,93 |
6,62 |
5,18 |
5,29 |
4,91 |
5,16 |
|
Айны Хокайдо |
3,23 |
3,81 |
4,07 |
2,21 |
0,00 |
1,80 |
5,22 |
5,05 |
4,15 |
4,20 |
5,00 |
3,55 |
4,10 |
3,71 |
4,57 |
|
Айны Сахалина |
2,83 |
4,09 |
3,67 |
2,67 |
1,80 |
0,00 |
4,63 |
4,66 |
3,64 |
3,61 |
4,82 |
3,43 |
3,06 |
2,71 |
3,24 |
|
Тунгусы |
3,15 |
3,86 |
3,99 |
6,66 |
5,22 |
4,63 |
0,00 |
1,26 |
2,88 |
2,11 |
1,46 |
2,07 |
3,10 |
3,51 |
3,03 |
|
Негидальцы |
2,85 |
3,57 |
3,83 |
6,80 |
5,05 |
4,66 |
1,26 |
0,00 |
2,01 |
1,51 |
1,49 |
1,98 |
3,10 |
3,46 |
3,83 |
|
Нанайцы |
2,09 |
3,42 |
3,29 |
6,02 |
4,15 |
3,64 |
2,88 |
2,01 |
0,00 |
1,17 |
3,05 |
2,48 |
2,50 |
2,69 |
4,05 |
|
Ульчи |
1,92 |
3,07 |
2,79 |
5,93 |
4,20 |
3,61 |
2,11 |
1,51 |
1,17 |
0,00 |
2,15 |
1,68 |
1,83 |
2,08 |
3,45 |
|
Орочи |
3,21 |
3,36 |
3,78 |
6,62 |
5,00 |
4,82 |
1,46 |
1,49 |
3,05 |
2,15 |
0,00 |
1,51 |
3,31 |
3,54 |
3,88 |
|
Нивхи |
2,02 |
2,41 |
2,79 |
5,18 |
3,55 |
3,43 |
2,07 |
1,98 |
2,48 |
1,68 |
1,51 |
0,00 |
2,46 |
2,54 |
3,35 |
|
Чукчи |
1,85 |
3,12 |
1,86 |
5,29 |
4,10 |
3,06 |
3,10 |
3,10 |
2,50 |
1,83 |
3,31 |
2,46 |
0,00 |
0,72 |
3,02 |
|
Эскимосы Чукотки |
2,07 |
3,27 |
2,02 |
4,91 |
3,71 |
2,71 |
3,51 |
3,46 |
2,69 |
2,08 |
3,54 |
2,54 |
0,72 |
0,00 |
3,14 |
|
Алеуты |
3,72 |
4,98 |
4,42 |
5,16 |
4,57 |
3,24 |
3,03 |
3,83 |
4,05 |
3,45 |
3,88 |
3,35 |
3,02 |
3,14 |
0,00 |
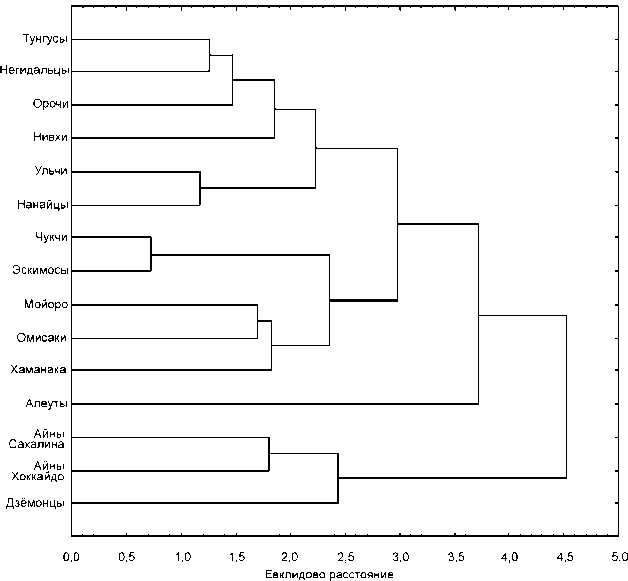
Рис. 6. Дендрограмма, полученная при кластеризации евклидовых расстояний невзвешенным парногрупповым методом.
ленной мере случаен и связан с особенностями примененного метода, который не позволяет отразить промежуточное положение метисных групп. Как было показано выше, охотцы занимают промежуточное положение между амурскими и арктическими группами.
Выводы
Основные итоги проделанного интеграционного анализа кранио-скопических и краниометрических признаков сводятся к следующему:
-
1. Носители охотской культуры (или охотских культур) сформировались в результате метисации, в которой участвовали арктические популяции, сходные с эскимосами и чукчам и, и древние группы тунгусо-маньчжуров, проникших на территорию Южного Сахалина с нижнего Амура в первых веках нашей эры.
-
2. Алеуты демонстрируют выраженную краниометрическую и краниоскопическую специфику, черты которой не наблюдаются в охотских группах. Это позволяет исключить алеутов из числа близких родственников охотцев.
-
3. Антропологическое своеобразие локальных охотских групп Хоккайдо может быть отражением различий в удельных весах амурского и арктического компонентов в их составе.
-
4. Восточные группы охотцев Хоккайдо имеют более выраженный протоайский компонент, чем северные.
Выражаю искреннюю благодарность проф. T. Амано (Университет Хоккайдо), проф. Х. Иcида (Университет Окинавы) и доктору Х. Мацумуре (Медицинский колледж Саппоро) за предоставленную возможность работать с краниологическими коллекциями и научные консультации, а также доктору Х. Оно (Университет Хоккайдо), без чьей практической помощи моя работа в Японии была бы невозможна. Я глубоко признателен А.Г. Козинцеву за критические замечания и помощь в работе.
