О распространении и биологии трясогузок в Туве

Автор: Цветков А.В., Редькин Я.А., Коблик Е.А.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 229 т.12, 2003 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149673
IDR: 140149673
On distribution and biology of wagtails in Tuva
Текст статьи О распространении и биологии трясогузок в Туве
Орнитологические наблюдения в Туве, в пределах современной Республики Тыва, проводили многие натуралисты (Дорогостайский 1908; Нестеров 1909; Сушкин 1914; Тугаринов 1915, 1916, 1927; Иоганзен 1929; Янушевич 1948; Янушевич, Юрлов 1950). Во второй половине XX в. сведения о птицах Тувы были существенно дополнены и уточнены (Спасский, Сонин 1959; Флинт 1962; Берман, Забелин 1963; Берман, Колонии 1967; Головуш-кин 1970; Берман, Злотин 1972; Забелин 1976; Емельянов 1986). Ряд работ представляет эколого-фаунистические очерки по отдельным, преимущественно редким и малоизученным видам (Берман 1967; Головушкин, Щербак 1976; Баранов 1991 и др.). Однако особенности гнездовой экологии, распределения, численности, пространственного взаимодействия таксономически близких форм целого ряда обычных и широко распространённых в регионе видов оставались практически неизученными.
В частности, это касается представителей рода Motacilla, ставших предметом нашего исследования. В настоящее время значительный интерес вызывает сложный характер таксономических и филогенетических отношений форм, входящих в комплексы “белых” и “жёлтых" трясогузок (Sushkin 1925; Johansen 1946; Grant, Mackworth-Praed 1952; Vaurie 1957; Sammalisto 1958, 1961, 1968; Береговой 1963; Назаренко 1968; Степанян 1983; Бахтадзе Казаков, 1985; Бахтадзе 1987; Гричик 1992 и др.).
В Туве в гнездовой период встречается 5 видов рода Motacilla, представленных 9 подвидами. Поскольку здесь на относительно небольшой территории представлено большое разнообразие ландшафтов, особый интерес приобретает изучение взаимоотношений разных форм в репродуктивный период. В этой работе мы приводим данные по экологии и поведению трясогузок во время гнездования. Поскольку подробный обзор пространственных и репродуктивных взаимоотношений близкородственных форм трясогузок на территории Тувы уже опубликован (Редькин 2003), мы ограничимся лишь общими сведениями о распространении и подвидовой принадлежности этих птиц.
Настоящее сообщение основано на результатах анализа материалов, полученных в ходе экспедиций в мае-июне 1999 и 2000 годов и обработки коллекционных материалов Зоологического музея Московского универстита и Зоологического института РАН. При изучении гнездовой биологии трясогузок проводили картирование территории их поселений с помощью глазомерной съёмки (Грюнберг 1991). Участки обитания, занимаемые парами, определялись методом точеч- ных регистраций (Odum, Kuenzler 1955) с изменениями, позволяющими сделать исследования более оперативными (Рябицев 1993). Статус исследуемых поселений определялся с помощью таблицы для диагностики основных типов групповых поселений птиц отряда Passeriformes (Цветков 2001). Питание гнездовых птенцов изучали методом наложения шейных лигатур (Мальчевский, Кадочников 1953; Титаева, Поливанов 1953). Пищевые объекты определялись, как правило, до семейства по “Определителю насекомых Европейской части СССР” под редакцией Г.Я.Бей-Биенко.
Сроки и комплексный характер экспедиционных работ не позволили нам получить одинаково полный экологический материал по всем обитающим в Туве видам трясогузок. Тем не менее, собранный материал может быть полезен не только при дальнейшем изучении трясогузок данной территории, но и для сравнения с данными из других регионов.
Неоднозначность понимания разными авторами ряда терминов, принятых при исследованиях территориального поведения птиц, требует конкретизации нашей позиции, без которой определение статуса исследуемых поселений может оказаться неточным. Участком обитания мы называем всё пространство, используемое парой во время репродуктивного периода для любых целей, а территорией — те части индивидуального участка, которые защищаются от особей своего вида (Рябицев 1993; Шилов 2001). В зависимости от характера использования пространства выделяют гнездовые и кормовые территории. Понятия гнездовой и охотничий (кормовой) участки используются в тех случаях, если эти функциональные части участка обитания не защищаются или же придание им статуса территории специально оговаривается в тексте. Термином поселение обозначаются группы граничащих или перекрывающихся друг с другом участков обитания птиц одного вида при отсутствии их в окрестных аналогичных местообитаниях (Рябицев 1993), т.е. между соседними поселениями должно существовать пространство, не входящее в их участки обитания и превышающее расстояние от геометрического центра поселения до его границ.
Под интерференцией мы понимаем такой тип конкурентных отношений и такой способ эксплуатации ресурсов, когда пространство, из которого черпаются последние, охраняется. В этом случае ресурсы или определённый ресурс становятся недоступными для соседних особей. Эксплуатационной конкуренцией называют совместное использование ресурсов на неохраняемом пространстве (Бигон и др. 1989).
При картировании поселений нами были приняты следующие допущения. Во-первых, гнездовые территории у разных видов трясогузок и в поселениях с разным уровнем социальных отношений существуют разное время. У трясогузок охрана гнездового участка осуществляется, как минимум, до выбора самкой места для будущего гнезда и, как максимум, до начала выкармливания птенцов. В дальнейшем птицы охраняют лишь место расположения гнезда. На карте поселения гнездовые участки показаны на стадии, когда они выполняли функции территорий. Во-вторых, в гнездовой период птицы довольно жёстко придерживаются границ своих участков обитания, однако периодически совершают дальние вылеты за их пределы (Nice 1941). Дальность таких перемещений может достигать 1 км и более. Исходя из значения понятия “участок обитания”, мы должны включать конечные пункты таких ориентировочных вылетов в его границы. Однако в рамках избранной методики исследований такой подход сильно осложнил бы проведение наблюдений. В связи с этим ориентировочными вылетами мы считаем единичные перемещения птиц за границы участка обитания, которые отвечают следующим критериям. Их дальность превышает среднее расстояние от геометрического центра участка обитания до его границ, а частота не превышает 1% от числа всех перемещений птиц на участке. Направления таких вылетов не являются постоянными и не связаны с поведением, характерным для репродуктивного периода (токование, сбор материала для гнезда, сбор корма для птенцов и т.д.). Ориентировочные вылеты, соответствующие этим критериям, при определении границ участка обитания не учитывались.
Характеристика района исследований
Исследования проводились в центральной, южной и юго-западной частях Тувы: в Тувинской котловине, в российской части Убсунурской, Саглинской и Урэгнурской котловин, а так же на южных макросклонах хребтов Западный и Восточный Танну-Ола, Цаган-Шибету, на хребте Сенгилен и в горном массиве Монгун-Тайга. Часть обследованных территорий входят в состав кластерных участков Убсунурского биосферного заповедника.
Котловины представляют собой плоские или холмистые равнины на высоте 770-1150 м н.у.м., занятые мелкодернистыми злаково-полынными степями, в большинстве мест с кустарниковой караганой Caragana pygmaea, С. bungei. В некоторых местах карагана покрывает до 40% площади степей. Местами в Убсунурской котловине и на большей части Саглинской и Урэгнурской котловин развиты глинистые и щебнистые пустыни, переходящие в каменистые шлейфы предгорий. В восточной части Убсунурской котловины небольшой участок занимает песчаная полупустыня с крупными барханами. Типичным элементом ландшафта являются останцы и небольшие останцовые хребты (примерно 400 м от подошвы до вершины). Для пойм крупных рек (Тес-Хем, Каргы, Саглы, Моген-Бурен) характерны галерейные редколесья из ив Salix, берёз Betula pendula, В. microphylla, тополей Populus nigra, Р. laurifolia, ольхи Alnus incana, черёмухи Padus avium, лиственницы Larix sibirica, часто с хорошо развитым кустарниковым ярусом. В долине р. Бора-Шай есть участки влажных лугов. В пределах котловин обследовано несколько пресных, солёных и горько-солёных озер (Торе-Холь, Чедер, Хадын, Чагатай, Шара-Нур, Убсу-Нур, Амдайгын-Холь). Общая площадь обследованных равнинных и низкогорных территорий составляет 2800 км2.
В горах исследованиями охвачены территории площадью 1200 км2. На высотах 1600-1850 м н.у.м. в Восточном Танну-Ола, восточных частях Западного Тан-ну-Ола, а также на хребте Сенгилен развит пояс лиственничной (иногда с примесью ели Picea obovata и единичными кедрами Pinus sibirica) тайги. Леса перемежаются участками горных степей с выходами скал и курумниками. Лиственничники пятнами встречаются на остепнённых склонах и выше. В центральной части хребта Сенгилен, в верховьях рек Нарын и Балыктыг-Хем, на высотах 2500-2715 м н.у.м. развита сухая горная тундра с элементами альпийских лугов.
На юго-западе Тувы горно-таёжный пояс редуцирован до галерейных лиственничников в верховьях рек и небольших изолированных участков на склонах Цаган-Шибету (1700-2600 м н.у.м.), а в западной части Западного Танну-Ола и массива Монгун-Тайга исчезает совсем (2250-3100 м н.у.м.). Ландшафты этих районов представляют собой горные степи с осыпями, курумниками, скалистыми обрывами, на некоторых выположенных участках и в долинах ручьёв развита мохово-осоковая горная тундра. В восточной части котловины крупного высокогорного озера Хиндиктик-Холь (2366 м н.у.м.) значительные площади заняты чрезвычайно увлажнёнными ерниковыми тундрами и горными болотами.
Основной материал, включающий в себя подробное описание поселений трясогузок, был собран на берегах озёр Тувинской и Убсунурской котловин. Озеро Чедер (2000 га) представляет собой мелководный горько-солёный водоем с топкими берегами и грязевыми отмелями, которые летом превращаются в солончаки и такыры. Площадь оз. Хадын составляет 3700 га. Это глубокое солёное озеро со слабохолмистыми твёрдыми берегами и хорошо выраженными тростниковыми зарослями. Узкая (10-30 м) полоса прибрежного разреженного разнотравья резко граничит с приподнятой полынно-злаковой степью. Озеро Торе-Холь (8150 га) — глубокий холодный пресный водоём с крутыми песчаными берегами. На редких пологих участках берега встречаются мелководные заливчики с солоноватой водой и тростниковыми зарослями. По берегу есть небольшие тополёв-ники, а основная его часть представляет песчаную полупустыню с кустами ка-раганы. Солёное озеро Шара-Нур (350 га) имеет плоские глинистые топкие берега. Тростниковые заросли занимают почти половину его береговой линии. Выше, сменяя друг друга, располагаются хорошо различающиеся травянистые сообщества, отражающие степень увлажнения и засоления почвы, переходящие в типичные полынно-злаковые степи.
Подвидовая систематика и распространение трясогузок
Жёлтая трясогузка Motacilla flava. На гнездовании представлена двумя хорошо различающимися окраской подвидами: М. flava beema (Sykes, 1832) и М. flava leucocephala (Przevalski, 1887). Сероголовая форма — белоухая жёлтая трясогузка М. f. beema распространена на большей части Западной и юге Центральной Сибири к востоку до западного склона Восточного Саяна (Портенко 1960; Степанян 1990). В Туве она встречается, главным образом, в пределах Туранской и Тувинской котловин (Сушкин 1914, 1932, 1938). Кроме того, этот подвид был обнаружен нами в пределах Уб-сунурской котловины. Белоголовая жёлтая трясогузка М. f leucocephala обитает по берегам водоёмов котловины Больших озёр на северо-западе Монголии, а также котловины оз. Убсу-Нур, в пределах которой проникает на юг Тувы (Vaurie 1959, 1960; Степанян 1990). Обе формы гнездятся только в равнинных частях республики, не проникая в горные районы. Судя по нашим сборам и полевым наблюдениям, а также по результатам изучения коллекционных материалов ЗММГУ и ЗИН, в Туранской и Тувинской котловинах гнездятся только фенотипически чистые популяции beema. На юге Тувы (южнее хребта Танну-Ола) предыдущими исследователями и нами гнездование жёлтых трясогузок было установлено по берегам озёр Убсу-Нур, Шара-Нур и Торе-Холь. В период наших исследований в 1999-2000 годах во всех трёх пунктах были обнаружены смешанные поселения, представленные как экземплярами чистых фенотипов обеих форм, так и значительным количеством особей, имеющих переходные признаки.
Желтоголовая трясогузка Motacilla citreola. На территории Тувы нами обнаружены три формы желтоголовой трясогузки, различающиеся общими размерами, пропорциями отдельных частей тела и особенностями окраски (Редькин 2003). Специфика распространения этих форм в южных районах Сибири заключается в их частичной разобщённости по занимаемым высотным поясам.
Малая желтоголовая трясогузка Motacilla citreola werae (Buturlin, 1907) встречается исключительно по берегам равнинных водоёмов, избегая горных районов. Вследствие этого её гнездовой ареал образует изолированные участки, охватывающие Туранскую, Тувинскую и, по крайней мере, север- ную часть Убсунурской котловины. По-видимому, к югу от Саян эта форма распространилась только во второй половине XX века, поскольку никем из исследователей, работавших там ранее (Сушкин 1914, 1925, 1932, 1938; Тугаринов 1915, 1916; Янушевич 1952), для территории современной Республики Тыва этот подвид не упоминался. Примечательно, что именно на этот период приходится активное расселение малой желтоголовой трясогузки в Европе в западном направлении.
Северная желтоголовая трясогузка Motacilla citreola citreola (Pallas, 1776) на юге Сибири гнездится преимущественно в высокогорьях, тогда как на равнинных участках встречается довольно редко. В Туве она была обнаружена нами гнездящейся на хребте Сенгилен у истоков р. Балыктыг-Хем. Кроме того, эта форма обитает в Восточно-Тувинском нагорье. Летом одна птица была добыта в Туранской котловине. Возможно, этот же подвид гнездится в Восточном Танну-Ола и на некоторых хребтах севера Монголии (Редькин 2003). Пролётные стайки желтоголовых трясогузок этого подвида наблюдались нами 22 мая 2002 на полях в окрестностях пос. Каа-Хем близ Кызыла.
Монгольская желтоголовая трясогузка Motacilla citreola quassatrix (Por-tenko, 1960), насколько мы можем судить по собственным наблюдениям на северной границе её ареала в Туве, а также по местам гнездовых находок в Монголии и на Алтае, одинаково часто встречается как по берегам степных или пустынных водоёмов на равнине, так и в высокогорьях. Наиболее северные гнездовые находки птиц этого подвида в Туве известны из следующих локалитетов: оз. Хиндиктиг-Холь, горный массив Мангун-Тайга, верховья р. Саглы в Западном Танну-Ола, водоёмы Убсунурской котловины. Пространственный контакт подвидов quassatrix и citreola, предположительно, имеет место в центральных частях хребта Танну-Ола, а также в пределах Алашского нагорья и Шапшальского хребта, примыкающего к Алтаю (Редькин 2003).
В местах совместного гнездования quassatrix и werae, обнаруженных нами по берегам озёр Торе-Холь, Шара-Нур и Убсу-Нур, количество фенотипических гибридов этих двух форм оказалось относительно невелико, что не укладывается в представления о зоне интерградации, характерной для географических рас одного вида. Учитывая наши наблюдения совместного гнездования пар, в составе которых оба партнера являлись представителями того или другого подвида, можно предполагать существование, по крайней мере, частичной репродуктивной изоляции между ними. Важно подчеркнуть, что определение всех птиц, собранных нами, было подтверждено анализом их митохондриальной ДНК (Pavlova et al, in press). Причём все чистые фенотипы quassatrix и werae всегда соответствовали хорошо отличимым генотипам первой и второй форм, что само по себе свидетельствует об отсутствии между ними широкой гибридизации. Возможно, дальнейшее накопление фактических материалов позволит рассматривать их отношения в свете концепций superspecies или ex-conspecies, а М. citreola werae — в качестве самостоятельного вида. Однако в настоящий момент имеющиеся данные не могут считаться достаточными для подобных заключений.
Белая трясогузка Motacilla alba, Маскированная трясогузка Motacilla personata. На территории Тувы гнездятся три формы группировки белых трясогузок: М. alba baicalensis (Swinhoe, 1871), М. alba dukhunensis (Sykes, 1832), а также M. personata (Gould, 1861), рассматриваемая в последние десятилетия в качестве самостоятельного вида, а не подвида М. alba (Степанян 1983, 1990; Ильяшенко 2001). Для маскированной трясогузки, широко распроспространённой в Средней Азии и на юге Казахстана, через Туву проводится восточная граница ареала (Степанян 1990). Эта форма обычна в Туранской и по всей Тувинской котловине, а также в Монгун-Тайгинском районе, где встречается преимущественно вблизи человеческого жилья и в лесистых долинах рек. Маскированная трясогузка отмечалась и по всему хребту Танну-Ола к востоку до долины р. Шуурмак, гнездящиеся пары были встречены в населённых пунктах Убсунурской котловины (Дус-Даг, Чаа-Суур, Ак-Чыраа, Самагалтай, Эрзин), на станах скотоводов и рыбаков в долине р. Тес-Хем и по берегам озёр Убсу-Нур и Шара-Нур. Кроме того, прежде эта форма добывалась на р. Ий (Ий-Хем) в Тод-жинском районе (Янушевич 1952).
Западносибирский подвид М. a. dukhunensis спорадично встречается на большей части территории Тувы, где, вероятно, вследствие недостатка брачных партнёров своей формы, ограниченно гибридизирует с personata (Редькин 2003). В гнездовое время dukhunensis добывалась в Тоджинском районе на оз. Азас (экз. в колл. ЗММГУ). Одиночная, возможно пролётная птица, встречена 22 мая 2000 в окрестностях пос. Каа-Хем близ Кызыла. В посёлке Эрзин взрослая птица, кормившая слётка, наблюдалась 20 июня 1999. Одиночная птица, вероятно неразмножавшаяся, встречена на оз. Торе-Холь 2 июня 1999. Два экземпляра с явными следами гибридизации с personata были добыты 31 мая в Монгун-Тайгинском районе в урочище Семигорки в долине р. Карты, а также во второй половине июня 2000 в окрестностях пос. Мугур-Аксы.
Байкальская форма М. a. baicalensis гнездится, главным образом, в юговосточных районах республики. В гнездовое время добывалась в верховьях р. Шивей (Улут-Шивей) на южном макросклоне хребта Хорумнуг-Тайга в Каа-Хемском районе (Янушевич 1952). Нами baicalensis отмечена только в Эрзинском районе. Отдельными парами она гнездится по берегам оз. Торе-Холь и в целом редка. В среднем и верхнем течении р. Нарын, в предгорьях и по южному макросклону хребта Сенгилен она полностью замещает маскированную трясогузку. В этих местах она становится немногочисленной. 19 июня 2000 на юго-восточном берегу оз. Торе-Холь из гнездовой пары был добыт самец со строго промежуточными признаками personata и baicalensis. Такой экземпляр оказался единственным среди наших сборов и изученных коллекционных материалов с территории Тувы.
Горная трясогузка Motacilla cinerea в Туве, как и на большей части Восточной Палеарктики, представлена подвидом М. с. melanope (Pallas, 1776). Немногочисленна, местами обычна (до 4-5 пар на 1 км маршрута) на горных реках всех обследованных хребтов. В равнинных ландшафтах котловин гоная трясогузка не гнездится. В Убсунурской котловине нами отмечались лишь одиночные пролётные и бродячие особи.
Характер поселений
Наиболее полный материал, касающийся структуры и статуса поселений, был получен при наблюдениях за жёлтыми, желтоголовыми и, отчасти, маскированными трясогузками. Первые два вида населяют одни и те же биотопы, иногда формируя смешанные поселения. Это обстоятельство заставляет нас отказаться от подачи материала в виде строгих повидовых очерков, что обусловлено необходимостью сравнительного анализа и позволит избежать нежелательных повторений.
Места гнездования жёлтых и желтоголовых трясогузок приурочены к пресным источникам, поймам рек и берегам озёр, в т.ч. солёных и горько-солёных. В большинстве случаев тростниковые заросли, участки луговой и лугово-степной растительности по берегам водоёмов используются птицами как кормовой биотоп, а гнёзда они устраивают на более высоких полынно-злаковых, часто с кустами караганы, участках берега. Первые пары приступают к гнездованию во второй декаде мая, а массовая откладка яиц начинается в третьей декаде месяца.
В Туве места, подходящие для гнездования жёлтых трясогузок, расположены фрагментарно, что определяет высокую концентрацию птиц на таких участках несмотря на то, что некоторые из них имеют крайне малые размеры. Одним из таких мест является пресный источник Кос-Булак, расположенный в 10 км к югу от г. Кызыл. Вокруг него в небольшом понижении рельефа сформировался участок влажного разнотравного луга размерами примерно 100x40 м. Здесь 28 мая 1999 мы отметили не менее 30 самцов жёлтых трясогузок. Самок же, приступивших к насиживанию, вспугивали на окружающих луговину остепнённых участках.
На восточном берегу солёного озера Хадын поселение жёлтых трясогузок было обнаружено 29 мая 1999. Вдоль уреза воды на 1 км учётного маршрута было отмечено 37 самцов и 19 самок М. f beema. Полоса прибрежного разнотравья шириной от 10 до 30 м и заросли невысокого тростника на мелководье использовались птицами в качестве общего кормового участка. Гнёзда находились в степи, отделённой от охотничьего участка довольно высоким, местами обрывистым краем озёрной котловины. Гнездовые участки располагались компактно на расстоянии 100-150 м от берега озера. Свободные от насиживания самцы и самки, периодически покидающие свои гнёзда, демонстрировали во время кормления выраженные элементы стайного поведения, которые заключались в синхронных перемещениях групп птиц (по 3-5 особей) между кормовым и гнездовым биотопом. Вдоль береговой линии птицы также перемещались группами, поддерживая индивидуальную дистанцию в 1.5-2 м, что в норме характерно для этого вида на пролёте.
Здесь же нам удалось наблюдать необычное для этого вида токовое поведение. В сторону тростниковых зарослей пролетела самка жёлтой трясогузки, преследуемая четырьмя самцами. Птицы летели плотной стайкой с характерным щебетом. В тростниках самка нырнула вниз и опустилась на залом прошлогодних стеблей. Самец, летевший первым, с лёта сделал садку и спарился с ней. Последний самец из “свиты” сделал круг над берегом и улетел в степь. Два других самца с щебетом опустились в 30-40 см от пары.
Первый самец соскочил с самки, но тут же повторил садку. На некоторое время птицы затихли, сидя в нижней трети тростниковых зарослей. Примерно через полминуты самка взлетела, и, преследуемая тремя самцами, улетела вдоль берега. По характеру и динамике происходящее больше всего напоминало групповой ток домовых воробьев Passer domesticus.
Повторное обследование восточного берега оз. Хадын было проведено 1 июля того же года. Учёт жёлтых трясогузок повторить не удалось, т.к. с 3040-метровой полосы берега вдоль тростников вспугивалось одновременно 5-6 десятков птиц, перемещающихся хаотично. Стайки состояли из небольшого числа взрослых птиц, сопровождавших объединённые выводки. Большинство молодых птиц были вполне самостоятельными. Отмечено также 2-3 выводка птенцов с недоросшими рулевыми перьями. Слётков докармливали взрослые птицы. На остепнённых и злаково-осоковых участках берега было отмечено не менее 5-6 пар, вероятно, приступивших ко второму или повторному репродуктивному циклу. Токовое поведение самцов в этих парах было обычным для вида.
Несмотря на кратковременность наших наблюдений на оз. Хадын, мы можем с уверенностью диагностировать наблюдавшееся здесь поселение Ьеета как колониальное. Об этом говорит высокая плотность гнездящихся пар, разделение кормового и гнездового биотопов, эксплуатационная конкуренция за пищевые ресурсы, ярко выраженное стайное поведение в гнездовой период, повлиявшее на характер токового поведения птиц.
Горько-солёное озеро Чедер, в отличие от оз. Хадын, имеет очень низкие, местами заболоченные берега. Здесь на обширном пространстве участки, занятые галофитами, чередуются с осоково-злаковым кочкарником, который начинает преобладать по мере удаления от уреза воды. Желтые трясогузки М. / Ьеета гнездятся на луговых участках берега и выше — на остепнённых участках. В этих условиях жёлтые трясогузки, избегая мест с разреженной растительностью на периодически затопляемой части берега, устраивают свои гнёзда не менее чем в 300 м от уреза воды. Несмотря на небольшое расстояние между озёрами (около 20 км), на оз. Чедер плотность гнездящихся пар была значительно ниже, чем на оз. Хадын. Она составила 8-12 пар на 1 км2. Жёлтые трясогузки гнездились здесь небольшими группами, чаще всего из 3 пар, т.е. их размещение в пространстве носило типичный контагиозный характер (Одум 1975). Гнездовые участки, перекрывающиеся по периферии, были окружены однообразным и обширным кормовым биотопом, что позволяло птицам из разных пар занимать и нди видуальные охотничьи участки, не имеющие чётких границ. При встречах друг с другом собирающие корм птицы демонстрировали реакцию взаимного избегания. Таким образом, взаимоотношения пар в группировках носили обычный в таких ситуациях проколониальный характер (Цветков 1994).
На озере Торе-Холь в Убсунурской котловине нами было обследовано смешанное поселение трясогузок, располагавшееся на косе, вдававшейся в озеро. Поселение занимало площадь 12.5 га. В центре этой территории находилось мелководное озерцо с тростниковыми зарослями, что значительно увеличивало длину береговой линии, где в основном и кормились трясогузки. На этой территории было отмечено 25 гнездящихся пар: 2 пары М. alba, 4 пары М. citreola и 19 пар М. flava, представленной здесь двумя формами: beema и leucocephala. Здесь же держалось два территориальных самца beema, которые, по-видимому, оставались холостыми (см. рисунок). В этом поселении белые трясогузки приступили к откладке яиц в 4-й пятидневке мая, т.е. на 6-8 дней раньше, чем первые пары желтоголовых и жёлтых трясогузок. В большинстве гнёзд откладка яиц началась в последних числах мая.
Гнездовые участки белых трясогузок носили полифункциональный характер, что определило их относительно крупные размеры (0.9-1 га). Они перекрывались между собой и полностью или почти полностью охватывали по 2-3 соседних участка жёлтых трясогузок. Кроме этого, белые трясогузки использовали общий кормовой участок вдоль береговой линии оз. Торе-Холь и внутреннего водоёма. Жёлтые трясогузки, раньше других приступившие к размножению (пары №№ 22 и 23), не использовали белых трясогузок в качестве пар-инициаторов. Таким образом, две пары белых трясогузок образовали минимальное по численности проколониальное поселение, которое не оказало существенного влияния на сроки размножения и распределение пар других видов трясогузок.
Желтоголовые трясогузки приступили к размножению практически одновременно с первыми парами жёлтых трясогузок. Но их роль как пар-инициаторов в смешанном поселении нам кажется неясной и, вероятнее всего, отсутствует. Вокруг пары №21 гнездовая группировка не сложилась. Рядом с парой №7 расположены территории жёлтых трясогузок №8 и № 9, но сроки их гнездования свидетельствуют о том, что скорее всего они заполняли пространство между группировками, сложившимися вокруг пар жёлтых трясогузок № 1 и № 11. В то же время взаимного влияния этих двух видов друг на друга в поселении отрицать нельзя. Во-первых, размножение жёлтых и желтоголовых трясогузок в восточной части поселения было синхронным. Во-вторых, желтоголовые трясогузки, гнездившиеся на описываемой территории, приступили к размножению почти на неделю позже, чем пары этого вида вне поселения, но на ближайшем к нему пространстве. Этот факт получает достойное объяснение только в том случае, если мы допускаем, что при межвидовых контактах роль социального доминанта играют жёлтые трясогузки.
На карте поселения (см. рисунок) видно, что оно территориально распадается на две части: восточную и западную. Несмотря на небольшие размеры озерца, птицы ни с той, ни с другой стороны на противоположный его берег не залетали. Южный берег этого водоёма посещался ближайшими к нему парами и из восточной и из западной группировки, но не интенсивно. Характер взаимодействия пар в восточной и западной частях также оказался различным, что даёт возможность рассматривать их как самостоятельные поселения.
В восточном поселении птицы, помимо индивидуальных кормовых участков, использовали и общее кормовое пространство вдоль берега внутреннего озерца. При редких встречах на индивидуальных охотничьих участках между особями преобладали нейтральные контакты, выражавшиеся чаще всего в виде взаимного избегания. Реже возникали конфликты без выраженного лидерства со стороны номинального хозяина. На общем кормовом участке все пары использовали стратегию эксплуатационной конкуренции
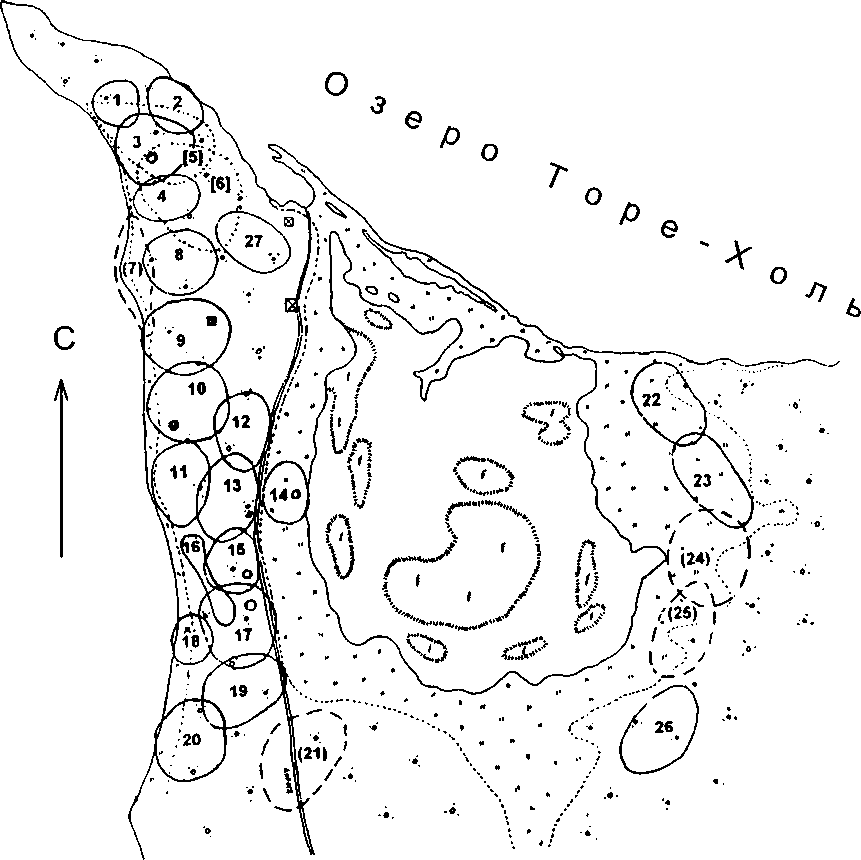
Схема смешанного поселения трясогузок на озере Торе-Холь.
Условные обозначения: сплошная линия — границы участков пар жёлтых трясогузок Motacilla flava ssp.; прерывистая линия — границы участков пар желтоголовых трясогузок М. citreola ssp.; пунктирная линия — границы участков пар белых трясогузок М. alba baicalensis ([5], [6]);
О — места расположения гнёзд; И — строения.
за пищевые ресурсы. Гнездовые участки этих пар занимали площадь порядка 0.5-0.6 га, их границы изначально пересекались, а функцию территории и отчетливые очертания они сохраняли до начала насиживания кладки. Между гнездовыми участками и кормовой территорией отчётливо выделялось пространство, используемое преимущественно для транзитных перемещений. С учётом этого пространства их участки обитания занимали площадь от 1.7 до 2.3 га и широко перекрывались. Структура и характер поведения птиц позволяют считать это поселение проколониальным. Оно состояло из 5 пар (3 пары М. flava и 2 пары М. citreola) и занимало площадь около 3.5 га.
Западное поселение, помимо белых и желтоголовых трясогузок, включало 16 пар и 2 холостующих самцов жёлтых трясогузок и занимало площадь около 5 га. В качестве гнездового биотопа жёлтые трясогузки исполь- зовали приподнятую песчаную часть косы с зарослями караганы и разреженным злаковым покровом. Кормились и собирали корм для птенцов они исключительно на общем кормовом участке, включавшем в себя береговую линию оз. Торе-Холь и внутреннего озерца. Границы гнездовых участков, площадь которых колебалась от 0.09 до 0.37 га, были крайне условными. От соседей защищалось, в основном, лишь место расположения гнезда, а функции территории участки выполняли лишь до момента начала строительства гнёзд. В западном поселении у жёлтых трясогузок наблюдалось отчётливое стайное поведение в репродуктивный период, включая групповой ток самцов, описанный выше для оз. Хадын. Среди жёлтых трясогузок западного поселения первыми приступили к гнездованию пары №№ 1, 11 и 14. Интересно, что пара № 14, поселившаяся в типичной для этого вида стации (прибрежное разнотравье), роль инициатора не сыграла. Возможно, это связано с тем, что гнездовой участок этой пары оказался на узкой полосе, интенсивно используемой в качестве общего охотничьего пространства. Таким образом, западное поселение жёлтых трясогузок состояло из двух субъединиц, сформировавшихся вокруг пар № 1 и № 11, пространство между которыми заполнили пары №№ 8, 9, 19 и 20, преступившие к размножению позднее. Здесь же, ближе к краю поселения, находились территории самцов, предположительно оставшихся холостыми.
Участки обитания всех пар жёлтых трясогузок имели сходную структуру и размеры. Они состояли из гнездового участка, который лишь на ранних стадиях репродуктивного цикла являлся территорией, а в дальнейшем уступал эту функцию незначительному пространству вокруг гнезда. В участок обитания входила общая кормовая территория, которая интенсивно использовалась птицами только в пределах своей субъединицы поселения. Интенсивно перемещаясь между кормовым участком и гнездом, трясогузки постоянно пересекали пространство, занятое гнездовыми участками соседних пар. В пределах поселения все пары придерживались территории своих субъединиц, сложившихся вокруг одной из пар-инициаторов. В результате размеры их участков обитания различались не сильно. Наиболее активно использовалось пространство площадью 1.2-1.4 га, которое составило ядро участка. Периферийные зоны участков, включавшие удалённые части общего кормового участка, тростниковые заросли и т.д., использовались в меньшей степени. С учётом этого пространства участки обитания трясогузок имели площадь около 7 га.
Оценка структуры поселения, поведения птиц и использования ими пространства даёт возможность диагностировать западное поселение как смешанную колонию, элементы которой, с учётом характера пребывания здесь белых и желтоголовых трясогузок, складываются более или менее независимо.
Колония жёлтых трясогузок на оз. Торе-Холь заслуживает внимания ещё с одной точки зрения. В предыдущие десятилетия трясогузки формы М. f. beema здесь не отмечались (Редькин 2003), так что в этом случаем мы имеем дело либо с пульсацией ареала, либо с устойчивым процессом расселения данной формы на юг и освоением ею новых гнездовых стаций. В этой связи особый интерес представляет фенотипический состав поселения (табл. 1). Здесь было зарегистрировано 17 фенотипически чистых особей
Таблица 1. Состав пар и стадии их репродуктивного цикла в групповом поселении трясогузок на озере Торе-Холь в Убсунурской котловине в июне 1999
|
№ пары |
Видовая принадлежность |
Пол |
Форма |
Репродуктивная стадия (на 22 июня) |
|
1 |
Motacilla flava |
M |
beema * |
|
|
F |
leucocephala * |
Слётки 11-12 сут |
||
|
2 |
Motacilla flava |
M |
beema = leucocephala * |
|
|
F |
beema = leucocephala * |
Кормят |
||
|
3 |
Motacilla flava |
M |
beema = leucocephala * |
|
|
F |
beema * |
5 птенцов 7-8 сут |
||
|
4 |
Motacilla flava |
M |
beema |
|
|
F |
beema |
Кормят |
||
|
5 |
Motacilla alba |
M |
baicalensis |
|
|
F |
baicalensis |
Слётки ~ 16 сут |
||
|
6 |
Motacilla alba |
M |
baicalensis |
|
|
F |
baicalensis |
Слётки старше 18 сут |
||
|
7 |
Motacilla citreola |
M |
quassatrix |
|
|
F |
quassatrix |
Слётки ~ 14 сут |
||
|
8 |
Motacilla flava |
M |
beema |
|
|
F |
beema |
Беспокойство у гнезда |
||
|
9 |
Motacilla flava |
M |
leucocephala * |
|
|
F |
leucocephala * |
Беспокойство у гнезда |
||
|
10 |
Motacilla flava |
M |
leucocephala |
|
|
F |
leucocephala |
5 птенцов ~5 сут |
||
|
11 |
Motacilla flava |
M |
beema |
|
|
F |
beema |
Слётки 12-14 сут |
||
|
12 |
Motacilla flava |
M |
leucocephala > beema |
|
|
F |
? |
Беспокойство у гнезда |
||
|
13 |
Motacilla flava |
M |
beema > leucocephala |
|
|
F |
? |
Кормят |
||
|
14 |
Motacilla flava |
M |
leucocephala > beema |
|
|
F |
? |
Слётки ~ 12 сут |
||
|
15 |
Motacilla flava |
M |
leucocephala > beema |
|
|
F |
7 |
Самец токует |
||
|
16 |
Motacilla flava |
M |
beema |
самец токует, |
|
F |
— |
предположительно холост |
||
|
17 |
Motacilla flava |
M |
leucocephala > beema * |
|
|
F |
beema = leucocephala * |
5 птенцов 8-9 сут |
||
|
18 |
Motacilla flava |
M |
beema |
самец токует, |
|
F |
— |
предположительно холост |
||
|
19 |
Motacilla flava |
M |
leucocephala > beema* |
|
|
F |
? |
Самец токует |
||
|
20 |
Motacilla flava |
M |
beema * |
|
|
F |
beema |
Беспокойство у гнезда |
||
|
21 |
Motacilla citreola |
M |
quassatrix |
|
|
F |
quassatrix |
Слётки 12-14 сут |
Продолжение таблицы 1
|
№ пары |
Видовая принадлежность |
Пол |
Форма |
Репродуктивная стадия (на 22 июня) |
|
22 |
Motacilla flava |
M |
beema |
|
|
F |
beema * |
Слётки 14-16 сут |
||
|
23 |
Motacilla flava |
M |
beema * |
|
|
F |
beema |
Слётки 14-16 сут |
||
|
24 |
Motacilla citreola |
M |
werae = quassatrix * |
|
|
F |
werae * |
Кормят |
||
|
25 |
Motacilla citreola |
M |
quassatrix |
|
|
F |
quassatrix |
Слётки 14-16 сут |
||
|
26 |
Motacilla flava |
M |
leucocephala |
|
|
F |
leucocephala |
Беспокойство у гнезда |
||
|
27 |
Motacilla flava |
M |
beema |
|
|
F |
leucocephala |
Беспокойство у гнезда |
? — подвидовая принадлежность неясна; * — птица добыта.
Обозначения в графе “Пол”: М — самец; F — самка.
Обозначения в графе “Форма” для экземпляров с промежуточными признаками: = — равное распределением признаков обеих форм; > — преобладание признаков одного из подвидов.
М. / Ьеета, 8 М. f leucocephala и 10 птиц с промежуточными признаками. При этом из 8 жёлтых трясогузок формы leucocephala 3 птицы были самцами и 5 — самками. Кроме того, учитывая, что подвидовая принадлежность ещё 5 самок трясогузок осталась невыясненной, разница в соотношении полов у этой формы могла быть и больше.
Из 10 самцов М. f Ьеета б образовали пары с самками своего подвида, 2 — с самками М. / leucocephala и ещё 2, предположительно, остались холостыми. Из 7 самок М. / Ьеета только одна образовала пару с самцом, имеющим промежуточные признаки. Все фенотипически чистые самцы М. / leucocephala образовали пары с самками своего подвида. Перечисленные птицы составили 12 пар из 19. Ещё в двух парах оба партнера имели переходные признаки. Оставшиеся 5 пар были представлены фенотипически промежуточными самцами и самками с неустановленной подвидовой принадлежностью.
Таким образом, с одной стороны, пары складываются в первую очередь из птиц одной формы. С другой стороны, большое число переходных экземпляров (и = 10) может рассматриваться как свидетельство того, что нарушение в соотношении полов у М. / leucocephala в поселении на озере Торе-Холь — явление постоянное. Последнее, вероятнее всего, объясняется тем, что эта форма находится здесь на северной границе своего распространения.
В июне 2000 в поселении трясогузок были проведены однодневные наблюдения. Численность и структура этого поселения остались примерно прежними. Как и в 1999 году, среди жёлтых трясогузок преобладали М. / Ьеета и особи с переходными признаками. Было встречено 3 фенотипически чистых самца М. f leucocephala, один из которых образовал пару с самкой промежуточного фенотипа, у которой преобладали признаки формы М. / leucocephala.
На берегах солёного озера Шара-Нур также была отмечена высокая плотность гнездящихся жёлтых и желтоголовых трясогузок. Берега этого водоёма низкие, местами заболоченные, с хорошо выраженной сменой растительных сообществ по мере удаления от уреза воды. К началу размножения желтоголовые трясогузки приступили во второй, а жёлтые — в третьей декаде мая. Подавляющее большинство трясогузок гнездилось вдоль восточного и южного берегов озера. Здесь первую, прибрежную, полосу растительности шириной от 50 до 80 м на подтопленных почвах составляют осоково-злаковые кочкарники и низкорослый тростник. Пространство между куртинами растений первого яруса занимают преимущественно галофиты. Вторая зона, шириной от 100 до 200 м, представляет собой растительные сообщества, развивающиеся на умеренно влажных почвах без признаков поверхностного засоления. Основу растений верхнего яруса составляют вейниковые кочкарники, а во втором ярусе растут ситники, лапчатки, примулы, чистяк и др. Проекционное покрытие растений второго яруса 80-100%. Третью зону прибрежной растительности шириной до 500 м представляют мощные, сильно разреженные куртины полупустынных злаков. Второй ярус выражен слабо и представлен бобовыми (в основном остролодочниками) и лапчатками. Его проекционное покрытие составляет от 5 до 40%. Выше располагаются типичные участки полупустынного разреженного полынно-злакового разнотравья.
Желтоголовые трясогузки гнездятся в первой, реже второй зоне, а кормятся в первой, преимущественно вдоль основного берега и на заросших тростником прибрежных лужах. Они образуют гнездовые группировки полуколониального типа, состоящие из 3-5 пар. Эти птицы используют общее охотничье пространство, применяя стратегию групповой интерференции. Участки обитания пар широко перекрываются, гнездовой участок выполняет функции территории только до момента выбора самкой места для будущего гнезда. В то же время элементов стайного поведения и полного перехода к эксплуатационной конкуренции за пищевые ресурсы, характерных для колониальных поселений, здесь не наблюдалось. Расстояние между гнёздами в поселениях колебалось от 25 до 50 м, а между поселениями — в среднем 170 м.
Жёлтые трясогузки на оз. Шара-Нур гнездятся во второй и третьей растительных зонах, а кормятся во второй, реже в первой, избегая полностью затопленных участков. Они образуют поселения проколониального типа из 3-4 пар, гнездящихся на расстоянии от 30 до 70 метров друг от друга. Среднее расстояние между поселениями составило 280 м.
Соотношение форм жёлтых трясогузок в этих поселениях было иное, чем на берегу оз. Торе-Холь. Здесь преобладали птицы белоголового подвида М. / leucocephala. Переходные признаки были отмечены только у 1 самки и 4 самцов. Фенотипически чистые самцы формы М. / Ьеета наблюдались в 1999 г. в этом месте лишь дважды. В 2000 г. здесь было добыто две самки этого подвида, самцы не отмечались.
Маскированные трясогузки Motacilla personata в Туве приступают к гнездованию первыми. Они гнездятся в городах, посёлках, вдоль берегов рек и т.п. Их численность и характер распространения, в общих чертах, совпадает с таковыми М. alba alba из средней полосы России (Цветков
1994). В Кызыле и его окрестностях первые пары начинают кладку примерно в третьей декаде апреля, тогда как массовая откладка яиц приходится на первую декаду мая. В Эрзине и на р. Тес-Хем отмечены две пары, приступившие к размножению в третьей декаде мая. Не исключено, что это были повторные кладки.
В населённых пунктах маскированные трясогузки поселяются как территориальными парами, так и формируют поселения про- и полуколониального типа, в зависимости от структурированности гнездового биотопа. В линейно вытянутых биотопах (дороги, берега рек) маскированные трясогузки поселяются отдельными парами, чаще всего на удалении друг от друга. Они избегают дорог, проходящих через степные территории, но встречаются на участках пологих, покрытых лесом перевалов, соединяющих Тувинскую котловину с Убсунурской и Урегнурской котловинами. В этих местах их плотность колеблется от 0.6 до 2 пар на 1 км маршрута. В районе Эрзина, вдоль берега р. Тес-Хем плотность маскированных трясогузок достигает 4-5 пар на 1 км маршрута. В этих условиях участки территориальных пар имеют линейную форму и могут перекрываться по периферии. Границы участков обитания маскированных трясогузок, так же как и у других форм комплекса “белых” трясогузок, определяются границами охотничьей территории. Территориальные пары используют стратегию интерференции в кормовом биотопе, но в зонах перекрывания участков обитания их поведение несколько изменяется. После ряда конфликтов и установления персональных отношений соседние самцы-резиденты либо используют эту часть охотничьей территории в отсутствии соседа, либо переходят в этой зоне к тактике эксплуатационной конкуренции, демонстрируя групповую интерференцию по отношению к изредка появляющимся здесь незнакомым птицам своего вида.
Две другие формы белых трясогузок (М. a. baicalensis, М. a. dukhunensis) и горная трясогузка на территории Тувы встречаются отдельными парами и поселений не образуют. В первом случае это связано в основном с невысокой численностью самих птиц, а во втором — с линейным характером гнездовых биотопов.
Питание гнездовых птенцов
Данные по рациону гнездовых птенцов были получены только для желтоголовой и жёлтой трясогузок.
От птенцов в возрасте 5-8 сут было собрано 89 (42 для М. citreola и 47 для М. flavd) пищевых проб. Эти пробы содержали 320 (97 и 223) пищевых объекта. Оба вида собирали корм для птенцов на общих кормовых участках, которые включали в себя береговую линию с прибрежной и водной растительностью. Тем не менее, рационы их птенцов значительно различались как по разнообразию приносимой взрослыми птицами пищи, так и по составу основных кормов (табл. 2).
В корме птенцов желтоголовых трясогузок преобладали крупные формы насекомых: личинки равнокрылых стрекоз (Odonata, Zygoptera), личинки водных полужесткокрылых (Hemiptera, Corixidae) и личинки ручейников Trichoptera. Каждая порция корма содержала от 1 до 5, в среднем 2.3
объекта. Жёлтые трясогузки кормили своих птенцов преимущественно мелкими двукрылыми: долгоножками и настоящими комарами (Diptera: Tipulidae, Culicidae). В некоторых порциях корма насчитывалось до 18, в среднем — 4.7 объекта.
Таблица 2. Состав корма птенцов жёлтой Motacilla flava leucocephala и желтоголовой М. citreola трясогузок на берегу озера Торе-Холь
|
Кормовые объекты |
М. citreola (42 пробы) |
М. flava (47 проб) |
||
|
Число экз. |
% |
Число экз. |
% |
|
|
MOLLUSCA |
3 |
3 |
2 |
0.9 |
|
Pulmonata с раковиной |
3 |
3 |
2 |
0.9 |
|
ARACHNIDA |
— |
— |
12 |
5.4 |
|
Araneidae imago |
— |
— |
6 |
2.7 |
|
Tetragnathidae imago |
— |
— |
6 |
2.7 |
|
INSECTA |
94 |
97 |
209 |
93.7 |
|
Odonata |
43 |
44.3 |
25 |
11.2 |
|
Zygoptera larvae |
38 |
39.1 |
14 |
6.3 |
|
Anisoptera larvae |
5 |
5.2 |
11 |
4.9 |
|
Orthoptera |
10 |
10.3 |
6 |
2.7 |
|
Acrididae larvae |
10 |
10.3 |
6 |
2.7 |
|
Hemiptera |
8 |
8.2 |
— |
— |
|
Corixidae larvae |
8 |
8.2 |
— |
— |
|
Trichoptera larvae |
12 |
12.4 |
1 |
0.45 |
|
Neuroptera |
— |
— |
1 |
0.45 |
|
Myrmeleontidae imago |
— |
— |
1 |
0.45 |
|
Lepidoptera |
— |
— |
3 |
1.35 |
|
Noctuidae imago |
— |
— |
1 |
0.45 |
|
Geometridae larvae |
— |
— |
2 |
0.9 |
|
Hymenoptera |
— |
— |
1 |
0.45 |
|
Myrmicidae imago |
— |
— |
1 |
0.45 |
|
Diptera |
21 |
21.8 |
172 |
77.1 |
|
Tipulidae imago |
15 |
15.6 |
94 |
42.1 |
|
Culicidae imago |
— |
— |
70 |
31.4 |
|
Tabanidae imago |
1 |
1 |
2 |
0.9 |
|
Syrphidae Imago, larvae |
5 |
5.2 |
6 |
2.7 |
|
Всего: % гидробионтов от общего числа кормовых объектов |
97 100.0 73.2 |
223 12 |
100.0 .5 |
|
В добыче желтоголовых трясогузок водные беспозвоночные встречались в 73.2% случаев, тогда как в корме птенцов жёлтых трясогузок гидробионты составили 12.5%. Различия в кормовых предпочтениях этих видов оказались гораздо более существенными, чем обнаруженные В.М. Чернышовым (1981) у этих видов в Северном Казахстане (49.6% и 40%). Различия в рационе птенцов жёлтых и желтоголовых трясогузок определяются способами добычи корма. Желтоголовые трясогузки — специализированные собирате- ли. Во время охоты они обследуют береговую линию вдоль уреза воды, часто заходят в воду, склёвывая насекомых с её поверхности. Передвигаясь по плавающим растениям, они осматривают их листья, в т.ч. и погружённые в воду. Короткие пробежки для преследования активной добычи они совершают редко, ещё реже используют пробежки с подлётом. Жёлтые трясогузки в первую очередь осматривают береговую линию и полосу прибрежных трав. Их кормодобывающее поведение включает те же элементы, что и у желтоголовых трясогузок, но в нём преобладают формы активного преследования добычи.
Экологическая разобщённость этих двух систематически близких видов на уровне кормовых предпочтений и способов кормодобывания позволяет им образовывать плотные и достаточно многочисленные совместные поселения, межвидовая конкуренция за пищевые ресурсы в которых, по всей видимости, невелика.
Заключение
Алтае-Саянский регион отличается таксономическим богатством представителей рода Motacilla. Для ряда форм в пределах Республики Тыва проходят границы гнездовых ареалов, некоторые из них представляются нам динамичными. Фрагментарность подходящих для гнездования биотопов определяет мозаичный характер распространения трясогузок и является причиной формирования ими плотных поселений в предпочитаемых стациях. Экологическая специфика форм трясогузок обусловлена различиями в численности, микробиотопических предпочтениях, сроках гнездования и особенностями рационов гнездовых птенцов. Эти различия позволяют некоторым из них занимать сходные гнездовые стации, несмотря на высокое суммарное обилие представителей группы в оптимальных местах обитания.
Особый интерес представляют данные о характере пространственных и репродуктивных отношений близкородственных форм белых трясогузок, подвидов жёлтых и желтоголовых трясогузок. Накопление таких сведений представляется чрезвычайно важным для уточнения таксономической структуры этих комплексов форм, служащих хорошей моделью для развития наших представлений о процессах видообразования.
Работа по изучению экологических и поведенческих особенностей разных видов и географических рас трясогузок в условиях ландшафтного разнообразия Тувы далека от завершения, некоторые аспекты их гнездовой биологии и экологии, репродуктивных отношений остались за рамками нашего исследования. Однако, мы надеемся, что полученные из этого ключевого региона материалы о сравнительной экологии и биологии представителей рода Motacilla позволят поставить новые вопросы и инициируют дальнейшую разработку темы взаимоотношений эволюционно молодых и генетически близких таксонов птиц.
Авторы выражают свою глубокую признательность председателю Госкомэкологии Республики Тыва Сергею Октяевичу Ондару, заместителю председателя Николаю Ивановичу Путинцеву, директору Убсу-Нурского биосферного заповедника Андриану Дугаровичу Додуку, заместителю директора по научной части Милану Маадыр-Ооловичу Кыныраа, соратникам по экспедициям 1999-2000 годов: Байлаку
Эртэнэ, Анатолию Никитину, Александру Куксину, Алексею Мосолову, Владимиру Сотникову, Андрею Манылову, Алексею Микулину, а также заведующему отделением орнитологии Зоологического института РАН Владимиру Михайловичу Лоскоту за предоставленную возможность работы с коллекциями своего учреждения.
