О теории параллелизма устойчивости картофеля к двадцативосьмипятнистой картофельной коровке Henosepilachna vigintioctomaculata (Сoleoptera:Соссinelladae) и колорадскому жуку Leptinotarsa decemlineata (Сoleoptera:Chrysomelidae) И.М. Гонтюрова: аналитический очерк
 и колорадскому жуку Leptinotarsa decemlineata (Сoleoptera:Chrysomelidae) И.М. Гонтюрова: аналитический очерк")
Автор: Мацишина Н.В., Фисенко П.В., Собко О.А., Ермак М.В., Киру С.Д.
Журнал: Овощи России @vegetables
Рубрика: Агрохимия, агропочвоведение, защита и карантин растений
Статья в выпуске: 1 (87), 2026 года.
Бесплатный доступ
Актуальность. Установление параллелизма устойчивости картофеля к колорадскому жуку и картофельной коровке было важным этапом для отечественного селекционного процесса в 60-е годы XX века. В своём труде «О параллелизме устойчивости картофеля к эпиляхне Epilachna vigintioctomaculata Motsch) и колорадскому жуку (Leptinotarsa decemlineata Say)» И.М. Гонтюров утверждал, не взирая на уже существующие публикации Ивановой, Куренцова и других исследователей, прямо указывающих на полифагию картофельной коровки, что полное развитие обоих вредителей возможно только на растениях рода Solanum, а личинки коровки не многоядны. Однако, еще А.И. Куренцов отмечал, что H. vigintioctomaculata типичный представитель маньчжурской фауны, приуроченный к области распространения смешанных и широколиственных лесов на Дальнем Востоке, и до начала земледельческой культуры в крае картофельная коровка обитала на травянистой растительности лесных полян, что вступает в прямое противоречие с постулатами И.М. Гонтюрова. Анализ экологических особенностей видов И.М. Гонтюровым был проведен неверно, что повлекло досадные ошибки при постановке, проведении и интерпретации результатов эксперимента. Цель данной работы – провести анализ существующих подходов к определению экологической валентности колорадского жука и картофельной коровки, в т.ч. и по нашим данным. Результаты. Описываемого в литературе параллелизма экологических валентностей между колорадским жуком и картофельной коровкой нет. Это связано с тем, что картофельная коровка значительно отличается своими биоэкологическими особенностями от колорадского жука: широкий полифаг, смена трофической и зимовочной стаций, саморегуляций плотности популяции, более высокие репродуктивный коэффициент и норма чистого потребления. Кроме того, значительно отличаются сортовые предпочтения картофеля, что говорит о разном влиянии вторичных метаболитов на онтогенез фитофагов. Всё это обеспечивает широкий фронт работ сотрудникам соответствующих селекционных учреждений.
Сорт, картофель, иммунитет, селекция, устойчивость, картофельная коровка, колорадский жук
Короткий адрес: https://sciup.org/140314219
IDR: 140314219 | УДК: 635.21:631.524.86(048) | DOI: 10.18619/2072-9146-2026-1-75-80
On the theory of the parallelism of potato resistance to the twenty-eight-spotted potato ladybird beetle Henosepilachna vigintioctomaculata (Сoleoptera:Соссinelladae) and the Colorado potato beetle Leptinotarsa decemlineata (Сoleoptera:Chrysomelidae) by I.M. Gontyurov: analytical essay
Relevance. Establishing the parallelism of potato resistance to the Colorado potato beetle and the potato ladybird beetle was an important step for Russian breeding in the 1960s. In his scientific work “On the parallelism of potato resistance to Epilachna vigintioctomaculata Motsch and Leptinotarsa decemlineata Say”, I.M. Gontyurov stated that the complete metamorphosis of the potato ladybird beetle and the Colorado potato beetle was possible only on plants belonging to the genus Solanum and that the larvae of the potato ladybird beetle were not polytrophic, which was contrary to the existing publications by Ivanova, Kurentsov, and other researchers reporting the polyphagous nature of the potato ladybird beetle. A.I. Kurentsov noted that H. vigintioctomaculata was a typical representative of the Manchuria fauna, populated the areas bordering on mixed coniferous and temperate broad-leaf forests in the Far East, and inhabited grasses in forest clearings before the onset of mass agricultural practices. This directly contradicted the postulate of I.M. Gontyrov. His analysis of the ecological characteristics of the species was erroneous, which led to further unfortunate mistakes in the design and performance of the experiment as well as in the interpretation of the research results. Materials and Methods. Our research goal was to analyze the existing methods for determining the ecological plasticity of the Colorado potato beetle and the potato ladybird beetle, including the use of our data. Results. There are no studies reporting the parallelism of the ecological plasticity of the Colorado potato beetle and the ladybird potato beetle. This is due to the fact that the bioecological characteristics of the potato ladybird beetle significantly differ from those of the Colorado potato beetle: polyphagous insect, migrates between trophic and winter hibernation sites, self-regulation of the population density, higher reproductive coefficient, and higher net consumption rate. Additionally, there are differences in the feeding preferences of the insect species for potato varieties. This suggests different influence of the secondary metabolites on the ontogeny of the phytophagous insects. All this provides a wide range of work for researchers and breeding institutions.
Текст научной статьи О теории параллелизма устойчивости картофеля к двадцативосьмипятнистой картофельной коровке Henosepilachna vigintioctomaculata (Сoleoptera:Соссinelladae) и колорадскому жуку Leptinotarsa decemlineata (Сoleoptera:Chrysomelidae) И.М. Гонтюрова: аналитический очерк
Обзор / Review
Н ачало земледельческой культуры стало переломным моментом в переходе насекомых-фитофагов в массовые вредители. Этому способствовало то, что окультуривание и селекция используемых в пищу растений шли по пути их обогащения питательными веществами в легко усвояемой человеком и животными форме, а также в направлении снижения содержания в растениях продуктов вторичного обмена, ухудшавших их пищевые качества и служивших иммунологическими барьерами в отношении вредных организмов. Следствием такого направления отбора растений явилось понижение физиологического иммунитета культурных растений к вредителям и одновременно улучшение физиологического состояния насекомых-фитофагов. Таким образом, селекция сельскохозяйственных растений, направленная на повышение продуктивности и пригодности в пищу для человека и домашних животных, явилась мощным фактором микроэволюции многих видов фитофагов. Между тем, устойчивые сорта стали мощным фактором, с помощью которого человек может регулировать численность и вредоносность многих фитофагов из числа насекомых. Принципиальные трудности в создании устойчивых к вредителям сортов сельскохозяйственных растений, характеризующихся высокой пищевой ценностью, в значительной степени преодолимы благодаря иммунологическому подходу селекции. В связи с тем, что такие методы должны занимать всё большую роль в сельскохозяйственной науке, очень важно правильно оценить реальные перспективы создания устойчивых сортов.
Установление параллелизма устойчивости картофеля к колорадскому жуку и картофельной коровке было важным этапом для отечественного селекционного процесса в 60-е гг. XX века. Исследования устойчивости картофеля к колорадскому жуку проводили Л.К. Антипова в 1952-1954 годах и Г.П. Слепушкина в 19551956 годах на Экспериментальной станции ВИР (Санкт-Петербург). Изучение устойчивости сортов картофеля к картофельной коровке проводил в 1957-1964 годах на Дальневосточной опытной станции ВИР (г. Владивосток) Гонтюров И.М. [1, 2]. В своём труде «О параллелизме устойчивости картофеля к эпиляхне
Epilachna vigintioctomaculata Motsch) и колорадскому жуку ( Leptinotarsa decemlineata Say)» И.М. Гонтюров утверждал, не взирая на уже существующие публикации Ивановой [3-5], Куренцова [6] и других исследователей, прямо указывающих на полифагию картофельной коровки, что полное развитие обоих вредителей возможно только на растениях рода Solanum , а личинки коровки не многоядны. Однако еще А.И. Куренцов [6] отмечал, что H. vigintioctomaculata типичный представитель маньчжурской фауны, приуроченный к области распространения смешанных и широколиственных лесов на Дальнем Востоке, и до начала земледельческой культуры в крае картофельная коровка обитала на травянистой растительности лесных полян, что вступает в прямое противоречие с постулатами И.М. Гонтюрова.
Как известно, кроме картофеля картофельная коровка сильно повреждает томат, огурец, тыкву, арбуз, кабачок, баклажан. Питание жуков на бахчевых культурах проходит весной и осенью, а летом жуки и личинки питаются главным образом листьями картофеля. Жуки и личинки выгрызают паренхимную ткань, скелетируют листья. Повреждения имеют вид "дорожек", идущих в разных направлениях от жилок, места повреждения приобретают сетчатый вид [7]. Кроме того, наблюдения А. Ивановой [5] показали, что жуки картофельной коровки могут развиваться без резкого угнетения на лопухе крупном Arctium lappa и тладианте сомнительной Thladiantha dubia .
Данные факты подтверждаются и на современном этапе исследований. Так, наши эксперименты убедительно доказывают, что картофельная коровка способна развиваться и давать плодовитое потомство на широком спектре кормовых растений [8, 9]. С точки зрения экологии, картофельная коровка классифицируется как эвритопный широкий полифаг, что само по себе означает способность получать жизнеспособное потомство на различных пищевых ресурсах.
Колорадский жук, в отличие от коровки, является узким стенотопным олигофагом [10]. Он трофически связан только с растениями семейства пасленовых (Solanaceae), относящимся к ряду видов и форм родов Solanum и Lycopersion . Таким образом, анализ экологических особенностей видов И. М. Гонтюровым был про-
Таблица. Степень расщепления сеянцев картофеля по устойчивости к колорадскому жуку и эпиляхне (при избирательном питании жуков) (цит. по Гонтюров, 1964)
Table. Degree of the divergence of resistance to the Colorado potato beetle and the potato ladybird beetle in potato seedlings (selective feeding of beetles) (cited from Gontyurov, 1964)
|
Род Solanum |
Выделено устойчивых сеянцев |
Процент устойчивых сеянцев |
||
|
к колорадскому жуку |
к эпиляхне |
к колорадскому жуку |
к эпиляхне |
|
|
S. laplaticum |
2 |
0 |
0,6 |
0 |
|
S.chacoense |
0 |
0 |
0 |
0 |
|
S.gibberulosum |
36 |
8 |
2,9 |
1,8 |
|
S. parodii |
10 |
1 |
3,1 |
1,9 |
|
S.schickii |
35 |
1 |
3,5 |
1,2 |
|
S. dolichostigma |
18 |
3 |
5,7 |
1,3 |
|
S. boergeri |
19 |
0 |
5,3 |
0 |
|
S. horovizii |
5 |
0 |
1,6 |
0 |
|
S. demissum |
0 |
23 |
0 |
1 |
Таблица I
Устойчивость различных аилов картофеля к колорадскому жуку и эпиляхне
|
Род Solan um |
Испытано образцов |
Процент образцов co средней гибелью личинок |
||||||||
|
0—20 |
21-50 |
51—80 |
81-100 |
|||||||
|
6 ° £ C S £ la |
с ж 212 |
И С 3 л — |
о ас • ж >» ° Ц * ° 2 9 Ж С. и |
се Е 2 |
■ «с о ж П -I * О 2 О |
а л X |
С ж ж |
08 = 3 л 3 |
||
|
S. commersonii С О — — — — — '00 '00 — S. laplaticum ПГ 21.1 33 36.8 67 10.6 — 31,5 — S. chacocnse 6 7 16.6 1 2.8 50 42.8 16.7 — 16.7 14.4 S. gibbcrulosum 79 60 7,7 33 18 . 27 45 21.7 29.3 18.3 S. parodii 25 26 3.0 34.6 27 42.2 63 15.5 7 7.7 S. scbickii 36 25 25 40 25 41 36 12 14 4 S. dolicliostigma 10 24 10 33 10 33 80 31 — — S. boergcri 12 9 8.3 22 25 67 25 11 41.7 — S. horovitzii 21 13 4.9 38.4 — 30,8 23.9 30.8 71.2 — S. garciac 3 7 — — — 71.4 100 28.6 — — S. demissum 59 125 8.5 16 27,1 48 28.9 28 35.5 8 . S. antipoviezii 7 44 — 50 28.6 27.2 28.6 20.5 42.8 2.3 S. jamesii 11 — — — — — — 100 100 |
||||||||||
Рис. 1. Страница 103, таблица 1 публикации «О параллелизме устойчивости картофеля к эпиляхне Epilachna vigintioctomaculata Motsch) и колорадскому жуку (Leptinotarsa decemlineata Say)» Fig. 1. Page 103, Table 1 of the publication «On the parallelism of potato resistance to Epilachna vigintioctomaculata Motsch and Leptinotarsa decemlineata Say»
веден неверно, что повлекло досадные ошибки при постановке, проведении и интерпретации результатов эксперимента.
Так, результаты опытов, приведенные в публикации, не соответствуют выводам (табл.). К сожалению, в тексте публикации для фактических данных не приведена статистическая обработка, и не указано, в абсолютном ли количестве или среднем арифметическом данные сведены в таблицы. Нами был проведен корреляционный анализ по Спирмену выделенных И.М. Гонтюровым устойчивых сеянцев дикорастущих представителей рода Solanum, приведенных автором (таблица). Расчёты показали отрицательную техническую корреляцию, значение R составило 0,1318, что указы- вает на очень слабую связь между переменными.
В своей работе сам автор отмечает, что около половины гибридов повреждались коровкой сильнее, чем колорадским жуком, при этом наиболее сходное поведение коровки и колорадского жука проявлялось только на 2 видах ( S. commersonii и S. jamesii ) из 13, что составляет 15% схожести [1]. При этом, в тексте статьи, на с. 102, автор указывает, что исследованные виды обладали большей устойчивостью к колорадскому жуку, чем к картофельной коровке. Однако, в выводах (с. 105 публикации) приводится дословно следующее: «Сопоставление устойчивости к колорадскому жуку и эпиляхне различных видов и отдельных образцов в пределах видов свидетельствует о наличии, за
Уход в диапаузу в начале октября, после питания на дикоросах и антропофитах
Уход в диапаузу в фазе «ягодообразование - усыхание» (конец августа)
Первые яйцекладки в фазе «образование листьев и стеблей»
Выход в фазе «образование листьев и стеблей» и «рост растений в длину»
Первые яйцекладки в фазы «всходы - смыкание рядков»
Зимует в листовом опаде. Для зимовки откочёвывает
Выход до всходов картофеля на дикоросы
Зимует в почве на поле, где питался
Широкий полифаг
Питается картофелем
Узкий олигофаг
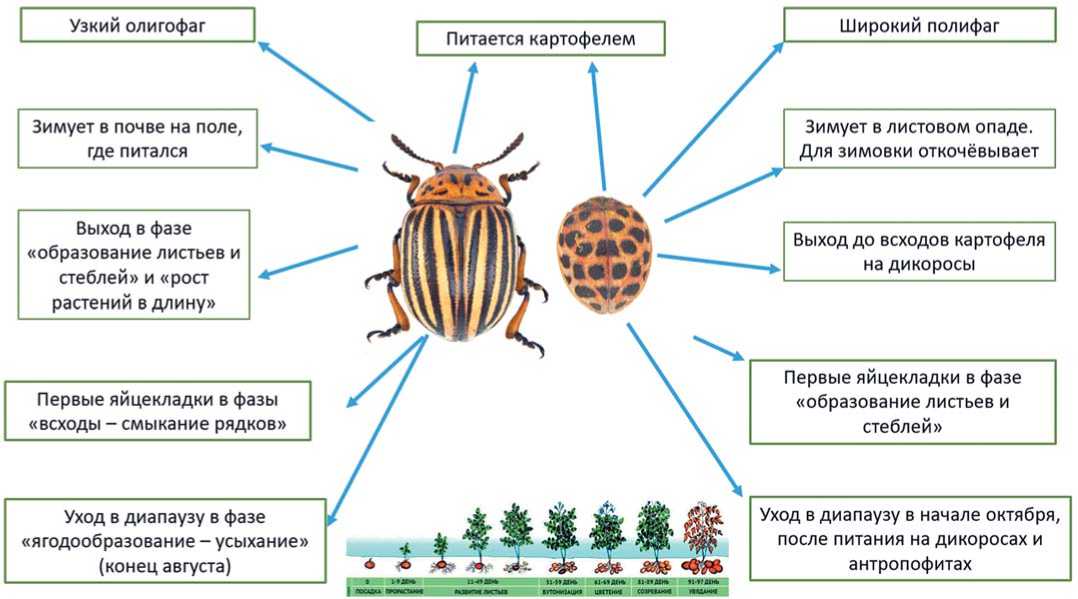
Рис. 2. Сравнительный анализ картофельной коровки и колорадского жука Fig. 2. Comparative analysis of the potato ladybird beetle and the Colorado potato beetle
небольшим исключением, параллелизма устойчивости картофеля к этим вредителям». Такое безапелляционное постулирование вызывает вопрос, является ли коэффициент схожести норм реакции картофеля на разных вредителей, выражаемый 15%, достаточным для безоговорочного принятия данной теории в качестве рабочей для селекционного процесса на устойчивость к листогрызущим вредителям.
Недоумение вызывает также содержание таблицы 1 «Устойчивость различных видов картофеля к колорадскому жуку и эпиляхне» (с. 103), публикации, где в графе «Испытано образцов» приводится различных объем экспериментальной выборки для каждого из фитофагов. Более того, для отдельных видов рода Solanum выборка представлена всего 1 образцом, что не допустимо для биологического исследования (рисунок 1).
В тексте публикации автор приводит факты о большей устойчивости образцов картофеля к колорадскому жуку, нежели к картофельной коровке, однако в 3 пункте выводов вновь утверждает об установлении параллелизма устойчивости картофеля к обоим вредителям.
По нашему мнению, данный постулат следует признать не только неверным, но и опасным. В результате сложилось ложное мнение, определившее на многие годы стратегию селекции картофеля на устойчивость к листогрызущим вредителям. Между тем, картофельная коровка обладает более широкой нормой реакции. Её пищеварительная система готова к встрече с довольно обширным числом веществ иммунного ответа растений именно в силу полифагии.
Кроме того, эти два вредителя достоверно различаются также особенностями экологии (рисунок 2). Проведенный анализ показал, что различия между про- должительностью стадии онтогенеза находятся на высоком уровне статистической значимости (p≤0,01). В целом, картофельная коровка проходит стадии онтогенеза быстрее колорадского жука, сходство отмечено лишь в сроках развития яйца [11].
Исследования фенологии колорадского жука показали, что в южных районах Приморского края выход жуков из почвы весной наблюдается в период начала вегетации картофеля, а в северных – в начале бутонизации. На юге Приморья выход первых перезимовавших жуков отмечается в третьей декаде мая, а интенсивное отрождение начинается при прогревании воздуха до +20°C и выше, и продолжается, как правило, в течение 6-9 дней. В Приморье период яйцекладки у колорадского жука очень продолжителен и длится до середины августа. Интенсивность яйцекладки зависит главным образом от температуры. Холодная погода со средней температурой +18°C заметно тормозит откладку яиц [12]. Наивысшая интенсивность яйцекладки совпадает с периодом самых длинных дней. В условиях Приморья это отмечается сразу после выхода жуков из зимовки, в конце мая – начале июня. Молодые самки первой генерации откладывают яйца, из которых развивается второе поколение фитофага, которое в Приморском крае закончить свое развитие не успевает. По нашим многолетним наблюдениям, фенология вредителя связана со сроками вегетации картофеля, что также является существенным отличием от картофельной коровки. Массовый уход имаго в диапаузу может наблюдаться уже в начале августа, когда листья картофеля поражаются грибными и псевдогрибными болезнями, что приводит к усыханию кустов, в то время как картофельная коровка продолжает своё питание на бахчевых культурах, других растениях семейства пас-
□ Картофельная коровка
□ Колорадский жук
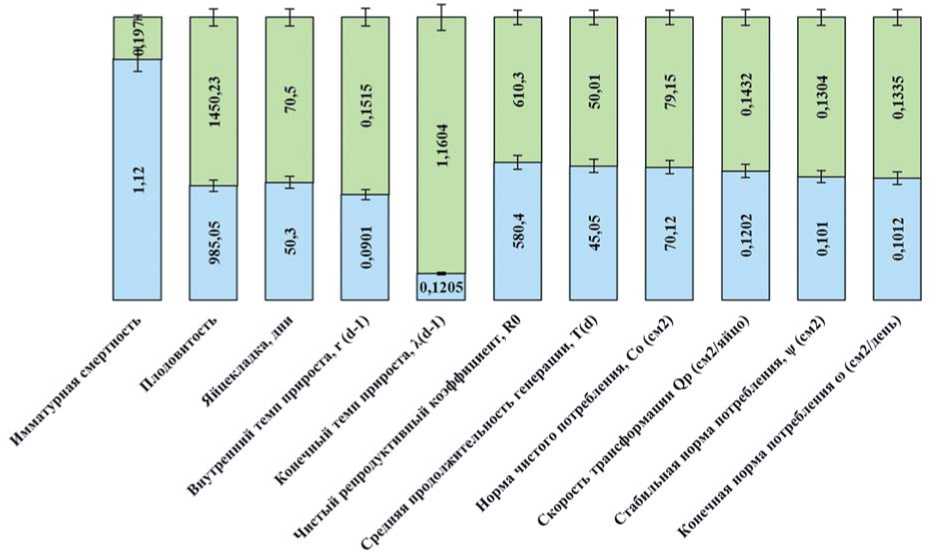
Рис. 3. Популяционные параметры исходных выборок картофельной коровки и колорадского жука (2008-2012 годы) (цит. по Matsishina et al., 2023)
Fig. 3. Population parameters of the original samples of the potato ladybird beetle and the Colorado potato beetle (2008-2012) (cited from Matsishina et al., 2023)
лёновые, сорняках, плодовых деревьях [13]. Расчётные показатели популяционных параметров двух фитофагов для периода 2008-2012 годов, представленные на рисунке 3, показывают, что внутренний темп прироста, чистый репродуктивный коэффициент, норма чистого потребления и коэффициент трансформации у картофельной коровки выше, чем у колорадского жука, даже в благоприятные для развития последнего годы [11].
Таким образом, описываемого в литературе параллелизма экологических валентностей между колорадским жуком и картофельной коровкой нет. Это связано с тем, что картофельная коровка значительно отличается своими биоэкологическими особенностями от колорадского жука: широкий полифаг, смена трофической и зимовочной стаций, саморегуляций плотности популяции, более высокие репродуктивный коэффициент и норма чистого потребления. Кроме того, значительно отличаются сортовые предпочтения картофеля, что говорит о разном влиянии вторичных метаболитов на онтогенез фитофагов. Всё это обеспечивает широкий фронт работ сотрудникам соответствующих селекционных учреждений.
С 1964 года селекционная работа на устойчивость картофеля к листогрызущим вредителям велась с опорой на теорию Гонтюрова. Это привело к созданию генотипов, не способных противостоять картофельной коровке, но позиционируемых как устойчивые. Игнорирование фактов, свидетельствующих об обрат- ном, спровоцировало увеличение пестицидной нагрузки на агроценозы вследствие потерей урожая от коровки. В данный момент остро стоит вопрос целенаправленной селекции на устойчивость сортов картофеля по отношению к H. vigintioctomaculata, поскольку численность колорадского жука на Дальнем Востоке ничтожна [12].
Мировой опыт растениеводства свидетельствует, что использование иммунных сортов сельскохозяйственных культур должно быть положено в основу интегрированной защиты растений [14]. Потребность в сортах, иммунных к вредителям, будет увеличиваться, причем в особенности возрастет значение таких сортов при переходе от интегрированной защиты растений к управлению агроэкосистемами, как это обусловлено приоритетными направлениями Доктрины продовольственной безопасности. Одна из причин нарушения механизмов саморегуляции в агроэкосистемах Дальнего Востока – это существенное влияние пестицидной нагрузки, обусловленное применением неустойчивых сортов, либо сортов, демонстрирующих устойчивость к патогенам и вредителям иного региона. Иммунитет растений к вредным организмам – это важнейшее биологическое свойство, эволюционно возникшее в результате длительного сосуществования с консументами и поддерживающее стабильность взаимоотношений фитофагов и их растений-хозяев в естественных экосистемах [15, 16].
Chrysomelidae ) in Primorsky Kray. Far Eastern Entomologist. 2023;(480):23-28.
Об авторах:
Петр Викторович Фисенко – кандидат биологических наук, ведущий научный сотрудник, исполняющий обязанности заведующего лаборатории селекционно-генетических исследований полевых культур, ,
Scopus Author ID: 26532574300, SPIN-код: 9916-1382
Ольга Абдулалиевна Собко – аспирант, научный сотрудник лаборатории селекционно-генетических исследований полевых культур, , Scopus Author ID: 57218617568, SPIN-код: 8082-5318,
Nataliya V. Matsishina – Dr. Sci. (Biology), Senior Researcher, Laboratory of Breeding and Genetic Research on Field Crops, ,
Scopus Author ID: 57218616526,
Petr V. Fisenko – Cand. Sci. (Biology), Leading Researcher, Acting Head of the Laboratory of Breeding and Genetic Research on Field Crops, ,
Scopus Author ID: 265325743001,
SPIN-code: 9916-1382
Olga A. Sobko – Post-Graduate Student, Researcher,
Laboratory of Breeding and Genetic Research on Field Crops, ,
Scopus Author ID: 57218617568,
Marina V. Ermak – Junior Researcher,
Laboratory of Breeding and Genetic Research on Field Crops, , SPIN-code: 1508-8155
Stepan D. Kiru – Dr. Sci. (Biology), Professor,
ISSN 2618-7132 (Online) Овощи России №1 2026
[ 80 ]
Vegetable crops of Russia №1 2026 ISSN 2072-9146 (Print)
