О выборе математических моделей кинетики культивирования дрожжей saccharomyces cerevisiae в условиях дефицита кислорода

Автор: Тишин В.Б., Меледина Т.В., Головинская О.В.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Процессы и аппараты пищевых производств
Статья в выпуске: 3 (65), 2015 года.
Бесплатный доступ
В технологиях производства многих пищевых продуктов микробиологические процессы являются определяющими в экономических показателях предприятий и качестве выпускаемой продукции. Примерами тому могут служить производства, в которых конечным продуктом является биомасса. Например, производство различных штаммов дрожжей Saccharomyces для различных отраслей пищевой промышленности: хлебопекарной, пивоваренной, винодельческой, а также для фармацевтической промышленности. Разработка математических моделей развития клеток микроорганизмов - одна из сложнейших задач микробиологии. Необходимость в поисках математических моделей диктуется постоянным развитием микробиологической отрасли промышленности, повышающимися требованиями к проектированию производств, ведению и предсказанию хода технологических процессов в зависимости от изменения технологических параметров. Но для этого необходимы знания о закономерностях материального и энергетического обмена между культуральной средой и клеткой и наличие математических моделей, описывающих их. Эти знания нельзя получить, не изучив кинетику протекания биологических процессов. Кинетические закономерности развития микроорганизмов во многом определяются выбором способа проведения микробиологического процесса и типа аппаратуры, в которой эти процессы протекают. Многие биологические процессы можно описать простейшими математическими моделями, однако существуют кинетические закономерности биологических процессов, которые можно описать только уравнениями более сложного вида. Получены модели кинетики культивирования дрожжей, отражающие сложности биологических процессов, протекающих во время культивирования. На примере обработки экспериментальных данных по культивированию дрожжей Saccharomyces cerevisiae периодическим способом получена система уравнений (математическая модель), устанавливающая функциональную связь прироста биомассы и потребления клетками углеводов с их различными начальными значениями в культуральной среде в условиях дефицита кислорода без какого-либо перемешивания.
Математическая модель, кинетика, культивирование, дрожжи saccharomyces cerevisiae
Короткий адрес: https://sciup.org/14040486
IDR: 14040486 | УДК: 663.12
About a choice of mathematical models of kinetics of cultivation of saccharomyces cerevisiae yeast in the conditions of deficiency of oxygen
In the production technology of many foods microbiological processes are crucial to the economic indicators of enterprises and the quality of the products manufactured. The examples of this are the production, where the biomass is the end product. For example, the production of various strains of the yeast Saccharomyces for different branches of the food industry: baking, brewing, wine-making, as well as for the pharmaceutical industry. The development of mathematical models of microbial cells is one of the greatest challenges of microbiology. The need to search for mathematical models is dictated by the continuous development of microbiological industry, increases in the requirements for the production design, maintenance and predictions of the processes depending on the change of process parameters. However, this requires knowledge of the laws governing material and energy exchange between the culture medium and the cell and the availability of mathematical models describing them. This knowledge cannot be obtained without studying the biological processes kinetics. Kinetic regularities of microbial growth is largely determined by the selection method of the microbiological process and the type of equipment in which these processes occur. Many biological processes can be described with a simple mathematical model, but there are kinetic regularities of biological processes that can only be described by equations of more complex type. Culturing yeast kinetic models, reflecting the complexity of the biological processes occurring during the cultivation were obtained. According to the analysis of experimental data on the Saccharomyces cerevisiae yeast culturing with a batch process, a system of equations (mathematical model), giving a functional relationship of biomass growth and cells consumption of carbohydrates with their different initial values in a culture medium under conditions of oxygen deficiency without stirring is obtained.
Текст научной статьи О выборе математических моделей кинетики культивирования дрожжей saccharomyces cerevisiae в условиях дефицита кислорода
Развитие микроорганизмов происходит под действием множества факторов, функционально связанных между собой. Установить эту связь в виде математических зависимостей (моделей) – задача сложная, интересная и необходимая. Необходимость ее решения диктуется высокими темпами развития микробиологических отраслей промышленности, возросшими требованиями к проектным работам, широкой автоматизацией технологических процессов и их оптимизации. Удовлетворить эти требования будет трудно, не имея данных по кинетике развития популяции клеток и математических моделей, адекватно отражающих течение биологических процессов. И не случайно проблеме моделирования кинетики биологических процессов уделяют большое внимание многие известные ученые [1, 2, 10].
Гипотеза о пропорциональности первой степени между приростом биомассы и ее концентрацией в культуральной среде.
Наиболее известной математической моделью кинетики развития микроорганизмов является экспоненциальная зависимость изменения концентрации клеток в культуральной жидкости во времени. Вывод модели основан на предположении о непрерывном делении клеток в неограниченном объеме, в условиях отсутствия дефицита питания и влияния отрицательных факторов, связанных с жизнедеятельностью клеток.
Согласно принятым условиям высказана гипотеза, что прирост биомассы А х за период времени Ат прямо пропорционален концентрации биомассы (клеток) в культуральной среде X и времени, т.е. А х = ц х Ат [1,2]. В дифференциальной форме это равенство имеет вид уравнения (1):
d x d т
ц х ,
где коэффициент пропорциональности ц носит название локальной удельной скорости прироста биомассы. После интегрирования уравнения (1) в пределах от х ,- до x J+1 и от т i до т i + 1 получим:
ц(т,,1 -т,-)
X + 1 = x i e + 1 . (2)
Кроме локальной удельной скорости, пользуются еще величиной средней удельной скорости ц определяемой в промежутке времени от т; = 0 до произвольного значения т и от x = хн до x , в таком случае из уравнения (2) следует [2]
X = Хн е цт . (3)
Чтобы при обработке опытных данных не быть зависимыми от размерностей, концентрации в культуральной жидкости в дальнейшем будем выражать в безразмерном виде, как отношение текущих концентраций к начальным: хь = х / х н. В новой записи уравнение (3) примет вид:
X b = е ^т. (4)
В конечном итоге, при переводе безразмерных величин в размерные, величина текущих значений концентраций биомассы будет зависеть от выбора размерности их начальных значений.
Уравнения (3) и (4) выражают закон экспоненциального развития микроорганизмов. Дальнейшая задача заключается в установлении зависимости удельной скорости ц от тех или иных факторов, например, температуры, pH среды, начальной концентрации микроорганизмов и субстрата; гидродинамических условий в культиваторе, последние в свою очередь зависит от интенсивности перемешивания культуральной среды, и т.п.
Обычно считается, что отклонение от экспоненциального закона говорит о несоблюдении ранее принятых условий. Удельную скорость роста при том или ином лимитирующем факторе выражают в виде различных зависимостей. В данной статье не ставится задача их анализа, а желающим детально разобраться в их сути, рекомендуем обратиться к литературе [1–5].
Гипотеза о степенной зависимости прироста биомассы от ее концентрации в культуральной среде.
Можно привести много случаев, когда развитие микроорганизмов не подчиняется экспоненциальной зависимости и условие (1) не всегда соблюдается. Более верным было бы предположение - Δ x ~. x m . Примером тому может быть развитие дрожжей Saccharomyces cerevisiae при наличии лаг-фазы, дефиците кислорода, когда принудительная подача воздуха в культуральную среду отсутствует, и кислород проникает в жидкость за счет молекулярной диффузии через ее свободную поверхность и т.п. Другим примером может служить культивирование гриба Medusomyces gisevi и др. [6–8]. Учитывая сказанное, более правильным будет записать дифференциальное уравнение (1) в ином виде:
dx = kx”. (5) dт
В уравнении (5) коэффициент k аналогичен удельной скорости μ в уравнении (1). В частном случае показатель степени m может равняться и единице. При m = 1 k = μ и в итоге получим уравнение экспоненты. Таким образом, уравнение (1) является частным случаем уравнения (5).
Интегрируя уравнения (5) в тех же пределах, что и уравнение (1), после приведения конечного выражения к безразмерному виду, получим уравнение степенного вида:
x 1 - m = 1 + (I - m ) k T .
b 1-m xн
Обозначив в правой части равенства (6) отношение параметров перед τ через δ и проведя несложные преобразования, запишем:
xb = (1+бт)1 - m.
Уравнение (7) можно привести к несколько иному виду:
xb = (1+5т) m1, где mi = —1—.(9)
1 — m
В уравнении (8) параметр 5 аналогичен ц в уравнении (3) и представляет собой среднею относительную удельную скорость прироста биомассы.
Интерес представляет анализ уравнения (9). Ранее уже было сказано, что при m = 1, интегрирование уравнения (5) дает экспоненциальную зависимость (4). При подстановке в уравнение (9) m = 1, функция m 1 ( m ) претерпевает разрыв и m 1 = ± ∞. Именно в зоне разрыва имеет место экспоненциальная зависимость (4). При m < 1, m 1 всегда будет больше нуля, и мы получим уравнение степенного вида (8).
При m > 1, m1 будет иметь отрицательное значение, и мы получим уравнение иного вида:
Sh =--- 1—
(1 + § , т) m 1 '
.
В данном случае Sb обозначает любой компонент культуральной жидкости, концен- трация которого снижается при протекании биологического процесса.
Уравнения, полученные на основе гипотезы степенного вида, требуют экспериментальной проверки. В дальнейшем будет показано, что область применения уравнений вида (8) и (10) ограничена определенными условиями. Как показали исследования по культивированию дрожжей Saccharomyces cerevisiae, уравнения типа (8) достаточно точно отражают характер накопления биомассы (рисунок 1). Однако уравнение (11), полученное на основе той же гипотезы (5), такой же достаточностью не обладает при описании опытных данных по кинетике потребления субстрата (рисунок 1а), поэтому для этой цели необходим поиск другой модели.
Эмпирическая степенная зависимость.
Уравнение степенного вида, несколько иного вида, чем (8) и (10), можно получить, основываясь не на гипотезе (5), а, скорее, из соображений логики, опираясь на экспериментальные результаты. Согласно графикам x = f ( т ) (рисунок 1), мы в праве в любой момент времени концентрацию биомассы в культуральной среде выразить в виде суммы:
x = x + A x , (11)
н где ∆x – прирост биомассы в промежутке времени от τ = 0 до τ = τi, в общем случае изменяющейся во времени. Можно записать, что ∆x ~ τn. Тогда, с учетом этой пропорциональности, уравнение (11) примет вид:
x = x + k т , (12)
н1, где k1 – коэффициент пропорциональности.
Разделив обе части равенства (12) на x н и обозначив отношение k I xH = у n , из уравнения (12) получим:
xb = 1 + ( YT ) n , (13)
где Y и n - параметры, определяемые экспери- ментально. Показатель степени в уравнении (13) может иметь как положительное, так и отрицательное значение. В последнем случае уравнение (14) примет вид:
S, =----- 1
b 1 + ( y s т ) n S
.
Таким образом, мы вместо уравнений (8) и (10) получили уравнения (13) и (14). Преимущество уравнений (13) и (14) перед уравнениями (8) и (10) заключается в том, что входящим в уравнение (13) и (14) параметрам проще придать вполне определенный физико-биологический смысл. Если в указанных уравнениях параметры m 1 и n имеют один и тот же смысл - темп изменения функции х ь (т) или Sb ( т ) , т.е. изменение скорости протекания биологического процессы во времени, то с параметрами δ и δ S дело обстоит сложнее. В отличие от δ в уравнении (8), из уравнения (13) легко доказывается, что величина ти = VY есть время удвоения функции x ( т ) , а из уравнении (14) - т5 = 1/ ys есть время снижения функции Sb ( т ) в два раза.
Время удвоения называют еще генерацией клеток. За все время культивирования дрожжей может быть несколько генераций, причем время генерации остается постоянной. Время генерации является важной характеристикой штамма дрожжей и используется в технологических расчетах.
Экспериментальное определение эмпирических коэффициентов
Прирост биомассы. Предметом исследований являлись дрожжи Saccharomyces cerevisiae штамма Л-12. Опыты проведены на мелассных растворах с начальным содержанием сахаров в культуральной среде S = 0,115; S =0,0575 и S =0.0383, в массовых долях. Растворы приготовлялись из мелассы с известным содержанием общих сахаров. Для получения необходимой концентрации сахаров в культуральной жидкости мелассу разбавляли водой. Засевная культура приготовлялась из прессованных дрожжей 75 % влажности. Для каждого начального значения субстрата было предусмотрено четыре значения начальной концентрации дрожжей в культуральной среде: x = 3,7; 10; 30; 50 кг/м3.
Культивирование проводилось в прозрачном стеклянном сосуде в течение 6-8 ч при температуре 30-31 °С при pH среды 4,2-4,6. Считалось, что изменение температуры и кислотности культуральной жидкости в указанных пределах не влияло существенным образом на процесс культивирования. Через каждый час отбирались пробы для определения общей концентрации дрожжей x – кг/м3 при 75 % влажности. Визуальные наблюдения показали, что большая часть дрожжей находилась во взвешенном состоянии и лишь незначительная – оседала на дно сосуда.
Результаты экспериментальных исследований и обработки опытных данных представлены на рисунке 1 в виде графиков зависимости x b ( т ) при . Для других значений S H графики выглядят аналогичным образом. Для сравнительной оценки уравнений (8) и (13), обработка экспериментальных даны, с целью нахождения значений коэффициентов δ, m 1 , γ и n , велась для обоих вариантов. Для облегчения работы, предварительный выбор уравнений для расчета коэффициентов производился с помощью компьютерных прграмм CurveExpert и MatchCad [9].
На основе опытных данных установлено, что входящие в уравнения (8) и (13) параметры, связаны сложными функциональными зависимостями с начальными значениями концентрации дрожжей и углеводов в культуральной жидкости.
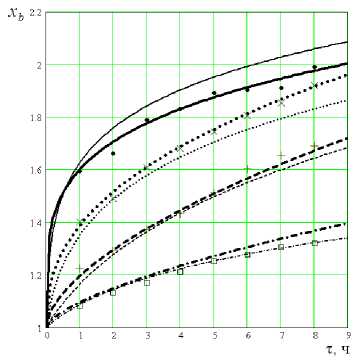
Рисунок 1. Изменение концентрации: биомассы во времени; тонкие линии соответствуют уравнению (8), толстые – (14) при S = 0,115; x = 3,7 ▬; x =10 •••; нн н
x =30 ▬ ▬; x = 50▬ • ▬.кг/м3; знакисоответствуют опытным данным.
Поиск математических моделей начнем с анализа возможности использования уравнения (8) в качестве головного. Из нескольких уравнений, предложенных программой CurveExpert для расчета величин δ и m1, при различных значениях x были выбраны следующие:
5 = к
b
a
V хн У
(8,а)
( x н b l)
m^ = a^e 2 c . (8,б)
Проанализируем полученные равенства. Прежде всего, отметим, что показатели степеней должны быть безразмерны. Значит безраз- мерными должны быть величины b , m и a , а коэффициенты a , b и c иметь размерность туже, что и x – кг/м3; так как при проведении экспериментов мы измеряли время в часах, то коэффициент k = 1, корректирующий размерность, имеет размерность, что δ – 1/ч.
Все указанные коэффициенты и показатели степеней в уравнениях (8,а) и (8,б) зависят от начальной концентрации углеводов в культуральной среде. Для их расчета на основе опытов выбирались соответствующие уравнения согласно требованиям, сформулированным выше. В результате, для расчета коэффициентов, входящих в уравнения (8а) и (8б), были выбраны следующие уравнения:
-( S н )
a = 1,62 Sh V 0,086 У ; (8,в)
b = 0,0133 • 0,85 S н S 295 . (8,г)
н
Проанализируем уравнения (8,в) и (8,г). Согласно рисунку 2а функция (8,в) имеет максимум Sн = 0,35. Далее, при 5 ^ 1, функция бун дет стремиться к нулю, что вполне логично. Возможно, значение Sн = 0,35 и соответствует тому предельному значению концентрации углеводов, при котором развитие дрожжей начинает тормозиться (требует экспериментальной проверки). При 5 ^ 0, а ^ 0, и согласно урав-н нению (8) 5 ^ 0, т.е. при отсутствии углеводов прироста биомассы, естественно, не будет.
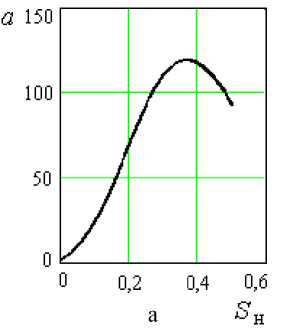
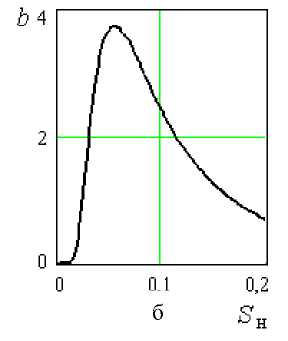
Рисунок 2. Зависимость коэффициентов a и b от S н в уравнениях (8,в) и (8,г)
Функция (8,г) при S н = 0,058 имеет максимум, следовательно и δ будет максимальным. Далее при 5 ^ 1 b ^ 0 (рисунок 2б) 5 ^ 1 . н
Таким образом влияние начального содержания углеводов S н на величину δ, выраженное уравнениями (8,в) и (8,г), не противоречит реальным условиям развития клеточной популяции и они могут быть использованы в дальнейшем при составлении математической исследуемого биологического процесса.
Недостаток функций типа (8,в) и (8,г) заключается в том, что аргумент функции входит в показатель степени, поэтому численные коэффициенты 0,086 и 1 должны иметь туже размерность, что и хн. Придать им какой-либо физико-биолгический смысл в таких уравнениях довольно сложно.
В уравнении (8,б) коэффициент a 1 определяет максимум значения функции m 1 ( х н ), а коэффициент b 1 - значение xH = хт н, соответствующее этому максимуму. С понижением начального содержания углеводов максимум темпа прироста биомассы возрастает. В тоже время он смещается в сторону увеличения x . В этой связи можно рекомендовать изменение начального засева x от 5 до 40 при отсутствии аэрации. При наличии принудительной аэрации культивирование можно проводить и при значительно более высоких значениях x [8, 9].
Для вычисления коэффициентов, входящих в уравнение (8,б) получены следующие эмпирические выражения:
_( 0,015 )
I а1 = 0,175 • 5н 1н j; (8,д)
b 1 = 17,57 + 94,46 5 н. (8,е)
Таким образом, получена система уравнений (8), (8,а)-(8,е), которую можно принять за математическую модель, так как она достаточно точно, согласно рисунку 1, описывает кинетику протекания процесса культивирования дрожжей, и позволяет прогнозировать течение процесса за пределами условий эксперимента в диапазоне изменения x от 1 до 50 и S от 0, 005 до 0,3. н н, ,
Обратимся теперь к уравнению (13). На основе экспериментальных данных была установлена линейная зависимость γ от x : н
Y = а 2 - Ь 2 • х н . (13,а)
Установлено, что в уравнении (13,а) b = 0,00186 м3/кк • ч независимо от значения S . Для расчета коэффициента a получено эмпирическое уравнение, устанавливающее его зависимость от начальной концентрации углеводов: 1
а 2 = 0,1028 • 0,983 5 н • 5 н 0,14 . (13,б)
Согласно равенству (13,б), в пределах изменения S от 0 до 0, 0013, можно принять а 2 = 0 и y = 0 . В таком случае прирост биомассы при 5 н< 0,0013 не происходит.
Таким образом, уравнения (13,а) и (13,б) устанавливают сложную функциональную связь удельной относительной скорости прироста биомассы с начальными значениями концентрации биомассы и углеводов в культуральной жидкости.
Для расчета темпа прироста биомассы n выбраны следующие уравнения:
-
a, • x
n =----—;
8,25 + x
н
(13,в)
(13,г)
a 3
0,428
0,06
Сравнивая модели уравнений (8) и (13),
Список литературы О выборе математических моделей кинетики культивирования дрожжей saccharomyces cerevisiae в условиях дефицита кислорода
- Алексеев Г.В., Вороненко Б.А., Лукин Н.И. Математические методы в пищевой инженерии. СПб: ЛАНЬ, 2012, 176 с.
- Nikolić S., Mojović L., Rakin M., Pejin J. et al. Simultaneous enzymatic saccharification and fermentation (SSF) in bioethanol production from corn meal by free and immobilized cells of Saccharomyces cerevisiae var. ellipsoideus//Journal of Chemical Science and Technology. 2012. V. 1. Issue.1. P. 21-26.
- Тишин В.Б., Оганнисян В. Г., Леонов А. В. Тепло-и массообмен между клеткой и культуральной средой при аэробном культивировании хлебопекарных дрожжей.//Научный журнал НИУ ИТМО. Серия: Процессы и аппараты пищевых производств. 2012. № 2.
- Арзамасцев А.А., Андреев А.Н. О возможности использования различных моделей кинетики биосинтеза.//Биофизика. 2001. T. 46. № 6. С. 1048-1061.
- Шлейкин А.Г., Кабанов А.В. и др. Изучение тиолдисульфидного равновесия в хлебопекарных дрожжах.//Вестник Международной академии холода. 2014. № 2. С. 62-65.
- Тишин В.Б., Головинская О.В., Леонов А.В. Математические и компьютерные методы в медицине, биологии и экологии: монография. М.: МИЭМП, 2013. С. 36-53.
- Тишин В.Б. Культивирование микроорганизмов. Кинетика, гидродинамика, тепломассообмен. СПб: РАПП, 2012. 180 с.
- Тишин В.Б., Аль Асаад Кусай М., Кхалил М.М. Исследование влияния некоторых факторов на кинетику роста дрожжей Saccharomyces cerevisiae при периодическом культивировании.//Вестник международной академии холода. 2007. № 1. С. 44-47.
- Макаров Е.Г. Инженерные расчеты в MatchCad: учебный курс. СПб.: Питер, 2005. 448 с.
- Дмитриева Ю.В., Андреев А.Н. Выбор дрожжей для технологии замороженной после формования сдобы.//Научный журнал НИУ ИТМО. Серия: Процессы и аппараты пищевых производств. 2015. № 3. С. 31-44.
