Об адаптивной внугрипопуляционной дифференциации холмогорской породы крупного рогатого скота

Автор: Матюков B.C.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы, итоги
Статья в выпуске: 6 т.42, 2007 года.
Бесплатный доступ
Исследовали полиморфизм белков молока и антигенов групп крови в популяции крупного рогатого скота холмогорской породы, расселенной в нескольких природно-климатических зонах. Тенденции распределения частот генов локусов р-Сп и к-Са согласуются с их клинальным распределением в мировой популяции вида Bos taunts L. Географическая клина, полученные доказательства селективной элиминации отдельных аллелей, несоответствие внутрипопуляционной дифференциации системе разведения породы, ассоциация антигенов "простых" систем групп крови с аллелями высокополиморфного 5-локуса свидетельствуют о вовлечении проанализированных полиморфных признаков в процесс отбора.
Порода, популяция, генотип, аллель, полиморфизм, отбор, адаптация
Короткий адрес: https://sciup.org/142133225
IDR: 142133225 | УДК: 636.2:575.17
About adaptive intrapopulational differentiation of Kholmogor breed of cattle
The polymorphism of milk proteins and antigens of blood groups were investigated in cattle populations of the Kholmogor breed settled in few natural-climatic zones. The tendencies in distribution of ft-Cn and к-Сп loci frequencies are in agreement with their cline distribution in world population of Bos taurus L. The geographic cline, obtained proofs of selective elimination of some alleles, discrepancy of intrapopulational differentiation to breeding system, association of antigens of "simple" blood groups system with alleles of high polymorphous j9-locus are suggest about involving analyzed polymorphous determinants in selection process.
Текст научной статьи Об адаптивной внугрипопуляционной дифференциации холмогорской породы крупного рогатого скота
Большинство работ, посвященных анализу генетической адаптации, выполнено на природных популяциях (1-7). Сельскохозяйственные животные в этом отношении менее изучены (1, 8, 9). Их генофонд в отличие от естественных популяций, кроме эволюционного давления, испытывает влияние искусственного отбора, поэтому адаптационные комплексы генов (АКГ) формируются в результате взаимодействий с селективными факторами обоих типов. При каждом конкретном сочетании воздействующих условий и генофонда в популяции отбираются специфичные АКГ. Выявив АКГ по сцепленным с ними маркерным генам, можно реализовать программу адаптивной селекции (10).
Анализ полиморфизма белков и ферментов показал, что по средней степени и размаху генетической изменчивости природные популяции и популяции сельскохозяйственных животных сходны (8, 11). Следовательно, принципиальных различий генетического ответа генотипа на действие искусственного и естественного отбора не существует. При этом тот и другой могут действовать одновременно как на одни, так и на разные полиморфные генетические системы.
Наиболее общим индикатором адаптивности полиморфных признаков в естественных популяциях считается географическая клина, которая служит косвенным подтверждением градиента естественного отбора (3). У сельскохозяйственных видов механизмы формирования географической изменчивости частот генов (полиморфных признаков) сложнее, поскольку увеличивается число взаимодействующих факторов, от которых зависит геногеографическая характеристика популяции. Поэтому изучение изменчивости частот генов в терминах генетической адаптации у сельскохозяйственных видов возможно только в многочисленной популяции, обитающей на обширном географическом пространстве в нескольких природно-климатических зонах. Одной из таких популяций, на наш взгляд, являлась холмогорская порода крупного рогатого скота. Это, пожалуй, единственная отечественная порода, которая длительное время не подвергалась интенсивному межпородному скрещиванию. В ХХ веке она поглотила отдельные отродья и популяции крупного рогатого скота, обитающие на пространстве от Кольского полуострова до Чукотки и Камчатки и от побережья Северного Ледовитого океана до центральных районов европейской части России и Сибири. В формировании генофонда холмогорского скота на протяжении нескольких столетий главенствующую роль играли племенные стада Архангельской, позднее и в меньшей степени — Московской, Кировской и Ленинградской областей, Республики Коми, Татарстана и Карелии. Если пренебречь влиянием на генофонд холмогорской породы не поддающихся точному учету эпизодических репродуктивных связей со скотом из низменных районов Западной Европы, то вплоть до конца 60-х годов ХХ века ее разведение в первичном очаге холмогорского скотоводства характеризовалось полной эндогамией, в остальной части популяции на протяжении примерно 20 поколений — частичной или полной экзогамией, поскольку поток генов был односторонним и направленным из первичного очага к периферии ареала породы. Таким образом, до середины 70 -х годов ХХ века геногеография холмогорской породы в чистом виде отражала взаимодействие генофонда крупного рогатого скота севера России с интегрирующим потоком генов из племенных стад и дифференцирующим воздействием экологических и селекционных факторов, которые адаптировали генофонд региональных популяций к территориальным условиям.
В последние десятилетия холмогорский скот, в первую очередь его лучшие по продуктивности стада, подверглись интенсивной ассимиляции голштинской породой. Однако это, на наш взгляд, еще более повышает ценность ретроспективного обсуждения геногеографической характеристики и эколого-генетической дифференциации холмогорской породы до ее скрещивания с голштинами, что и послужило целью настоящей работы.
Методика. Анализировали материалы, собранные за период с 1968 по 2002 годы при выборочных обследованиях скота холмогорской породы в хозяйствах Республики Коми и Карелии, Архангельской и Московской областей (ведущие стада племенных заводов, репродукторов и товарных хозяйств, расположенных в нескольких природно-климатических зонах). База данных содержала сведения о группах крови по девяти полиморфным системам у 4590 коров и 260 быков-производителей, а также о полиморфных белках молока у 3600 коров. В нескольких поколениях коров учитывали происхождение (генеалогию), молочную продуктивность за несколько лактаций, годы рождения и отела, продолжительность жизни, причины выбытия, плодовитость и другие признаки, необходимые для генетического анализа. Группы крови определяли в отделе животноводства Научно-исследовательского и проектно-технологического института агропромышленного комплекса Республики Коми, а также частично использовали результаты проведенной нами генетической экспертизы происхождения, выполненной по племенным хозяйствам в Ветеринарной лаборатории Республики Коми. Методика определения соответствовала принятым инструкциям (12). Антигены групп крови в индивидуальных образцах типировали с помощью наборов реагентов (производство Армавирской биофабрики и Самарской областной лаборатории иммуногенетики) согласно рекомендациям по применению (13). Для генетико-популяционного анализа по группам крови использовали также опубликованные данные (14) и записи в документах на производителей, завезенных в Республику Коми из других регионов Российской Федерации. Электрофоретический анализ белков молока в крахмальном геле в щелочной буферной системе проводили по общепринятой методике (15), в кислой — как описано (16) в нашей модификации (17).
Частоты антигенов, генов и генотипов рассчитывали по П.Ф. Рокицкому (18), оценку неравновесного сцепления генов выполняли по Р. Левонтину (2). Генетико-популяционные параметры по маркерным признакам, а также выживаемость и плодовитость определяли с использованием соответствующих алгоритмов (5, 19). Для подсчета антигенов, генов и генотипов в выборках результаты определения групп крови и полиморфных белков у животных заносили в таблицы с последующей сортировкой и расчетом частоты распределения животных-носителей (в долях единицы или процентах) по отдельным генотипам, генам и антигенам в программе MS Excel (зарегистрирована в отраслевом фонде алгоритмов и программ 4.12.2006). Эколого-генетический анализ заключался в сопоставлении показателей и выявлении различий между генерациями, генеалогическими (генетическими) группами животных и генотипами по количественным признакам и частотам генов в идентичных и градиентных экологических условиях. Для оценки животных по признакам молочной продуктивности использовали данные за первую лактацию, так как они в наибольшей степени характеризуют популяцию (репродуктивную группу животных) до отбора.
Как известно, при исследовании частной генетики любого объекта, особенно медленно размножающегося и долго живущего вида, требуется проведение непрерывного ряда многолетних наблюдений (20). Кроме того, поиск прямых доказательств селективного значения полиморфизма предполагает ревизию выборок до и после отбора. В экспериментах с крупным рогатым скотом применить такой подход сложно, поскольку приходится анализировать выборки, составленные из особей разного возраста, то есть в разной степени подвергшихся отбору, и происходящие от сравнительно небольших по численности групп самцов. Поэтому в настоящей работе животных аттестовали по генетическим вариантам полиморфных белков молока и группам крови, формируя по каждой природной зоне выборки из нескольких репродуктивно и пространственно разобщенных стад и продолжая наблюдения до естественного выбытия всех аттестованных животных.
Результаты. Г е н е т и к о - п о п у л я ц и о н н а я х а р а к т е- р и с т и к а х о л м о г о р с к о й п о р о д ы п о п о л и м о р ф - н ы м с и с т е м а м б е л к о в м о л о к а и г р у п п а м к р о в и. Каппа - казеин (к- Сп ). В молоке чистопородных холмогорских коров выявили два генетических варианта κ -казеина — А и В, синтез которых соответственно контролируют кодоминантные аллельные гены к-СпА и к-СпВ . Средняя частота к-СпА по обследованной популяции составляла 0,689, к -СпВ — 0,311 (табл. 1). Некоторое повышение частоты к-СпВ обнаружили в субпопуляции из Республики Коми и северо-таежной зоны. Встречаемость редкого и хозяйственно наиболее ценного генотипа к-СпВВ (20) в целом по выборке составила 0,087, в субпопуляциях Республики Коми — 0,099 и Архангельской области — 0,081. По отдельным регионам нарушения генетического равновесия по генотипам к-Сп не выявили, однако в целом по исследованной выборке нарушение равновесия было близким к достоверному за счет избытка гетерозигот (0,10 > Р > 0,05).
Бета - казеин (Р-Сп). При гель-электрофорезе в кислой буферной системе выявили три варианта в-казеина: A 1, A 2 и В. Наиболее часто встречался A 2 , затем A 1 , реже других — В. По частоте аллелей Р-Сп заводские и репродукторные стада различались незначительно. Проявлялась некоторая тенденция к повышению частоты встречаемости аллеля в-СпА1 в субпопуляциях Республики Коми и лесотундровой зоны.
Различия по частотам аллелей в-Сп между массивами холмогорского скота Республики Коми, Архангельской области и в среднем по племенным заводам оказались несущественными. Однако при формировании выборки по природным зонам для лесотундровой частота аллеля Р-СпА2 снижалась до 0,45, Р-СпВ — до 0,10, а Р-СпА1 — возрастала до 0,45 (см. табл. 1), что достоверно отличало животных этой группы от скота племенных заводов и таежной зоны (Р < 0,01).
1. Частота аллелей по локусам в-Cn и к-Cn в субпопуляциях коров холмогорской породы в зависимости от региона, природной зоны и направления разведения
|
Группа животных |
п |
Р-Сп |
к -Сп |
|||
|
А1 |
1 А2 |
В |
A |
1 В |
||
|
Популяция в целом |
2907 |
0,37±0,008 |
0,50±0,008 |
0,13±0,006 |
0,69±0,006 |
0,31±0,006 |
|
Субпопуляция региона: |
||||||
|
Архангельская область |
1236 |
0,35±0,010 |
0,51±0,011 |
0,14±0,007 |
0,71±0,009 |
0,29±0,009 |
|
Республика Коми |
1495 |
0,37±0,012 |
0,50±0,013 |
0,13±0,009 |
0,67±0,009 |
0,33±0,009 |
|
Субпопуляция природной зоны: |
||||||
|
лесотундровая |
549 |
0,45±0,015 |
0,45±0,015 |
0,10±0,009 |
0,75 ± 0,013 |
0,25 ± 0,013 |
|
северо-таежная |
1674 |
0,34±0,010 |
0,51±0,010 |
0,15±0,008 |
0,66 ± 0,008 |
0,34 ± 0,008 |
|
средне-южная таежная |
684 |
0,35±0,017 |
0,51±0,017 |
0,14±0,012 |
0,71±0,012 |
0,29±0,014 |
|
Стадо племенных заводов |
916 |
0,34±0,011 |
0,51±0,012 |
0,15±0,009 |
0,71±0,010 |
0,29±0,010 |
П р и м е ч а н и е. По Московской области и Республике Карелия данные включены в группу «Племенные заводы», поскольку обследовали по одному племенному хозяйству.
Антигены систем групп крови (эритроцитарные антигены — ЕА ). По системе А (ЕАА ) у холмогорского скота из Республики Коми выявлены антигены А и Z' (табл. 2). Наибольшей частотой антигенов А и Z' характеризовалось голштинизированное стадо из ГУП «Южное». У чистопородного холмогорского скота трех племенных заводов частота антигена А колебалась в пределах от 0,328 до 0,471, антигена Z' была низкой (см. табл. 2).
В системе С (ЕАС) обнаружили семь антигенов: С и Е с частотой встречаемости по 0,650, W — 0,386, R — 0,234, X — 0,272, L — 0,230. Наибольшие колебания показателя по стадам установлены для антигенов W, R и X .
Система S (EAS) была представлена антигенами S 1 , H', U, U', H" с максимальной частотой антигена H' — 0,770. Вариация частот антигенов в стадах зависела от абсолютных значений частот: наибольшие величины коэффициентов вариации установили для антигенов с низкими, наименьшие — с высокими частотами.
В системе F-V (EAF-V) преобладал аллель F, соответствующий антигену F с частотой 0,843. По системам EAJ, EAL, EAM, EAZ частота представленных антигенов J, L, M, Z составила соответственно 0,214; 0,244; 0,176 и 0,317.
В н у т р и- и м е ж п о р о д н а я д и ф ф е р е н ц и а ц и я п о п у л я ц и и. Данные о распределении частот антигенов по восьми системам групп крови у животных холмогорской породы в хозяйствах Республики Коми, Архангельской и Московской областей (см. табл. 2) указывают на зависимость внутрипопуляционной дифференциации холмогорского скота от региона разведения. Так, в популяции Республики Коми частота наиболее распространенных антигенов, как правило, имела промежуточное значение относительно показателей в популяциях Архангельской и Московской областей, а в системе групп крови EAC частота антигенов W и X была соответственно существенно выше и ниже, чем в этих популяциях.
Внутри субпопуляции холмогорского скота Республики Коми генетически выделялись два стада: одно — чистопородное стадо ООО «Большая Инта» в лесотундровой зоне, другое — стадо ГУП «Южное» на крайнем юге Республики Коми (табл. 2). Анализ частоты распределения аллелей антигенов отдельных систем групп крови и генов, контролирующих синтез белков молока (см. табл. 1), показал, что внутрипопуляционная дифференциация холмогорского скота зависит также от природной зоны разведения. В частности, генетически выделялись стада лесотундровой зоны: по частотам генов, контролирующих синтез белков молока, по сравнению со скотом из таежной зоны (а также племенных заводов) они в большей степени приближались к северному европейскому (North European) скоту и чернопестрому скоту низменностей Европы (Black Pied Lowland) (12).
-
2. Характеристика крупного рогатого скота холмогорской породы из разных регионов разведения и с неодинаковой кровностью по частотам антигенов систем групп крови
я 05 О &
С с 2
Я и
я
Я я
<
Республика Коми
о
ч о S X
X
чистопородный скот
я 11 я
5 я S |S 5 £
X ± s x , _ ( n = 4590)
и
1
2
3
>я
я ч
О -5
о 11
О -5
Q
ч я
СО
8^
О -5
Я S
ч
И чо
§v
О -5
А
А
0,3500
0,414
0,471
0,373
0,539
0,429±0,0343
17,8
0,333
0,401
Z´
0,001
0,0005
0
0,019
0,005±0,0037
203,4
0
0
—
0,650
0,585
0,529
0,627
0,461
0,570±0,0342
13,4
0,667
0,599
C
C
0,584
0,691
0,630
0,733
0,610
0,650±0,0273
9,4
0,619
0,814
E
0,602
0,664
0,624
0,641
0,718
0,650±0,0199
6,8
0,598
0,793
W
0,425
0,325
0,332
0,289
0,560
0,386±0,0489
28,3
0,163
0,184
R
0,215
0,188
0,155
0,398
0,214
0,234±0,0424
40,5
0,184
0,365
X
0,201
0,188
0,303
0,304
0,366
0,272±0,0339
27,8
0,348
0,503
C´
0,087
0,090
0,098
0,079
0,065
0,084±0,0056
14,9
0
0
L´
0,246
0,234
0,207
0,289
0,175
0,230±0,0191
18,6
0,116
0,405
—
0,005
0,014
0,025
0,005
0
0,010±0,0044
100,9
0,083
0,008
F
F
0,829
0,783
0,892
0,817
0,896
0,843±0,0220
5,8
0,777
0,828
V
0,171
0,217
0,108
0,183
0,104
0,157±0,0220
31,4
0,223
0,172
J
J
0,162
0,188
0,237
0,303
0,181
0,214±0,0254
57,0
0,163
0,261
—
0,838
0,812
0,763
0,697
0,819
0,786±0,0254
10,4
0,837
0,739
L
L
0,263
0,261
0,201
0,230
0,267
0,244±0,0127
11,6
0,203
0,466
—
0,651
0,739
0,746
0,770
0,733
0,728±0,0202
6,2
0,797
0,354
M
M
0,147
0,193
0,168
0,087
0,283
0,176±0,0321
40,8
0,168
0,501
—
0,852
0,807
0,832
0,914
0,717
0,824±0,0322
8,7
0,832
0,499
S
S
0,151
0,181
0,128
0,210
0,18
0,170±0,0140
18,5
0,314
0,280
H´
0,830
0,806
0,708
0,732
0,772
0,770±0,0226
6,6
0,811
0,937
U
0
0,006
0,004
0,003
0,062
0,015±0,0118
175,8
0
0
U´
0,02
0,061
0,099
0,190
0,227
0,119±0,0389
72,9
0,167
0,192
U´´
0
0,031
0
0
0
0,006±0,0062
223,6
0
0,006
H´´
0,001
0,002
0
0,001
0,006
0,002±0,0010
117,3
0
0
—
0,096
0,171
0,234
0,164
0,202
0,173±0,0230
29,7
0,083
0,016
Z
Z
0,377
0,406
0,331
0,126
0,347
0,317±0,0495
34,9
0,329
0,221
—
0,623
0,594
0,669
0,874
0,592
0,670±0,0528
17,6
0,671
0,779
-
3. Распределение дифакториальных генотипов по локусам β-Lg и β-Cn в разных популяциях скота холмогорской породы (Республика Коми)
β-Lg
β-Cn
В целом
А1А1 1
А1А2
1 А2А2
А1В
А2В 1
ВВ
С т а д о
п л е м
е н н о г о з а в о д а
О А О «П р
и г о р о д н ы й»
АА
0
2,0
6,0
1,0
6,0
1,0
16
1,3
5,6
5,3
1,2
2,0
0,6
АВ
10,0
27,0
27,0
7,0
12,0
3,0
86
6,9
30,2
28,5
6,5
10,5
3,4
ВВ
11,0
63,0
54,0
12,0
14,0
6,0
160
13,0
56,1
53,1
12,2
19,5
6,1
В целом
21
92
87
20
32
10
262
П о п у л я ц и я в
ц е л о м
АА
14,0
31,0
19,0
4,0
14,0
1,0
83
12,0
29,0
22,0
7,0
11,0
2,0
АВ
91,0
226,0
152,0
67,0
74,0
10,0
620
80,0
230,0
159,0
61,0
78,0
12,0
ВВ
135,0
415,0
301,0
107,0
144,0
27,0
1129
148,0
413,0
291,0
110,0
143,0
24,0
В целом
240
672
472
178
232
38
1832
П р и м е ч а н и е. Над и под чертой —
соответственно фактическое и теоретическое распределение генотипов. Для
П р и м е ч а н и е. 1, 2 и 3 — соответственно южная, центральная и северная часть Республики Коми. В Архангель- ской и Московской областях для расчета частот антигенов использовали племенные документы и каталоги. Прочерк означает отсутствие антигена.
Внутрипопуляционную дифференциацию холмогорской породы нельзя объяснить системой разведения, поскольку ранее было показано, что дифференциация генеалогических линий быков по частотам генов, контролирующим полиморфизм белков молока, была незначительной, а интенсивность использования производителей, судя по среднему числу дочерей от одного быка в стаде, — невысокой.
Что касается оценки генетического сходства холмогорского скота с другими породами, отметим, что по частотам генов, контролирующих синтез белков молока, полиморфных белков и ферментов сыворотки крови, антигенов «простых» систем групп крови, у холмогорского скота наибольшее сходство наблюдается с близкой по географической зоне разведения ярославской породой. От чернопестрого скота заводская популяция холмогорской породы уклоняется в сторону пород красного корня (22-24), а по частотам генов многоаллельного локуса ЕАВ она равноудалена от ярославской и черно-пестрой пород (13).
стада племенного завода ОАО «Пригородный» частоты аллелей генов β-Cn : A1 — 0,294, A2 — 0,529, B — 0,177; β-Lg :
A — 0,225, B — 0,775. Критерий соответствия и уровень значимости χ2 = 0,078; Р > 0,999. Для популяции в целом коэффициент неравновесия сцепления D1 = fAA1ŁfBA2 - - fAA2ŁfBA1 = 0,0070; D2 = fAA1ŁfBB - fABŁfBA1 =
0,0028; D 3 = f AA2 Łf BB - f AB Łf BA2 = 0,0015. Критерий соответствия и уровень значимости нарушения независимого комбинирования генотипов по локусам β-Lg и β-Cn χ 2 = 8,9; Р > 0,5.
Н е р а в н о в е с н о е с ц е п л е н и е г е н о в в п о п у -л я ц и и. Наблюдения выполняли в популяции холмогорского скота Республики Коми в целом и в стаде ОАО племенного завода «Пригородный», в котором длительное время проводилась оценка быков-производителей по качеству потомства. Методика оценки быков исключала ассортативные спаривания, то есть была гарантирована рандомизация выборок быков, и в стаде реализовывался принцип панмиксии. Полученные данные (табл. 3) опровергают гипотезу об ассоциированном комбинировании генотипов по несцепленным локусам β-Lg и β-Cn в дигибридной популяции.
Распределение генотипов в дигибридной популяции по сцепленным локусам β-Сn и κ-Cn в стаде племенного завода ОАО «Пригородный» и популяции в целом (табл. 4) достоверно отклонялось от теоретического, рассчитанного, исходя из гипотезы о независимом комбинировании аллелей этих локусов.
Анализ распределения антигенов по шести системам групп крови в пределах отдельных феногрупп EAВ -локуса выявил ассоциированное комбинирование аллелей антигенов J и М с феногруппами системы групп крови B в четырех стадах из пяти (табл. 5).
По чистопородному (племенной репродуктор, ООО «Большая Инта», лесотундровая зона) и голштинизированному стаду (ГУП «Южное», зона средней тайги) нарушение свободного комбинирования антигенов наблюдали соответственно по пяти и четырем системам из шести. Кроме того, в стаде племенного репродуктора нарушение свободного комбинирования аллелей антигенов системы ЕАА и EAB оказалось близким к достоверному. По другим стадам ассоциация антигенов была не столь очевидна и затрагивала антигены одной-двух систем групп крови. Ассоциированное распределение аллелей антигенов «простых» систем групп крови с аллелями ЕАВ -локуса в стаде ГУП «Южное», по-видимому, является следствием скрещивания скота холмогорской и голштинской пород, то есть исходным различием аллелофонда и антигенной характеристики коров холмогорской породы и голштинских быков. В этом стаде нарушение независимого комбинирования антигенов может быть обусловлено и случайным отклонением распределения генов у голштинских быков из-за малочисленности их выборки. В чистопородном стаде племенного репродуктора ООО «Большая Инта» система разведения скота исключает подобную детерминацию неравновесия сцепления.
4. Распределение дифакториальных генотипов по ло кусам в~Сп и к-Cn в разных популяциях скота холмогорской породы (Республика Коми)
|
κ -Cn |
β-Сn |
В целом |
|||||
|
А1А1 |
А1А2 |
А2А2 |
А1В |
А2В |
ВВ |
||
|
С т а д о п л е м е н н о г о |
з а в о д а |
О А О |
«П р и г |
о р о д н ы й» |
|||
|
АА |
14,0 |
59,0 |
46,0 |
1,0 |
0 |
1,0 |
121 |
|
10,2 |
43,8 |
39,2 |
9,0 |
14,6 |
4,2 |
||
|
АВ |
8,0 |
31,0 |
34,0 |
17,0 |
25,0 |
0 |
115 |
|
9,8 |
41,6 |
37,1 |
8,5 |
14,0 |
4,0 |
||
|
ВВ |
0 |
3,0 |
3,0 |
1,0 |
6,0 |
8,0 |
21 |
|
1,8 |
7,6 |
6,8 |
1,6 |
2,5 |
0,7 |
||
|
В целом |
22 |
93 |
83 |
19 |
31 |
9 |
257 |
|
П о п |
у л я ц и я |
в ц е л о м |
|||||
|
АА |
35,0 |
102 |
83,0 |
1,0 |
1,0 |
0 |
222 |
|
26,0 |
81,5 |
67,3 |
16,6 |
24,6 |
6,0 |
||
|
АВ |
24,0 |
78,0 |
61,0 |
34,0 |
43,0 |
0 |
240 |
|
29,6 |
89,0 |
70,8 |
18,0 |
26,5 |
6,0 |
||
|
ВВ |
2,0 |
7,0 |
7,0 |
3,0 |
12,0 |
13,0 |
44 |
|
5,4 |
16,4 |
12,9 |
3,4 |
4,9 |
1,0 |
||
|
В целом |
61 |
187 |
151 |
38 |
56 |
13 |
503 |
П р и м е ч а н и е. Критерий соответствия и уровень значимости нарушения независимого комбинирования генотипов по локусам β-Cn и κ -Cn для стада племзавода и популяции в целом — соответственно χ 2 = 176,6; Р < 0,001 и χ 2 = 242,1; Р < 0,001. Для стада племзавода частота аллелей генов β-Сn : А1 — 0,303; А2 — 0,564, В — 0,133; κ -Cn : A — 0,692, B — 0,308; для популяции в целом — β-Cn : А — 0,135, В — 0,865; κ -Cn : А — 0,679, В — 0,321. Над чертой — фактическое, под чертой — ожидаемое значение.
Из всех обследованных групп стадо ООО «Большая Инта» содержалось в наиболее жестких условиях, так как хозяйство расположено в лесотундровой зоне, с чем связаны трудности в обеспечении полноценного кормления и ухода за животными.
-
5. Оценка ассоциированного распределения аллелей антигенов «простых» сис
тем групп крови с аллелями В-локуса по критерию χ2 в популяциях скота холмогорской породы из разных хозяйств (Республика Коми)
|
Хозяйство |
Аллель |
|||||
|
A |
F ** |
J |
L |
M |
Z |
|
|
ОАО «Пригородный» |
2,82 |
0,57 |
7,10* |
2,19 |
4,99* |
0 |
|
ООО «Ухта-97» |
0,01 |
8,79 |
23,44* |
1,05 |
1,32 |
0,01 |
|
ООО «Изваильский-97» |
5,29* |
0,01 |
0 |
2,30 |
8,90* |
0,52 |
|
ООО «Большая Инта» |
7,89 |
13,37* |
23,63* |
17,77* |
23,52* |
20,99* |
|
ГУП «Южное» |
21,80* |
24,39* |
10,53* |
1,87 |
20,33* |
0,96 |
|
* Нулевая гипотеза о соответствии фактической частоты антигена теоретической отвергается при уровне значимости Р |
||||||
< 0,05.
** Частота генов.
Как показано, у коров молочная продуктивность косвенно характеризует напряженность обмена веществ и физиологическую нагрузку на организм. Следовательно, мо-жно считать, что молочная продуктивность тесно связана с жизнеспособностью и плодовитостью, то есть комплексной приспособленностью генотипов, обусловленной балансом действия искусственного и естественного отбора. При этом естественный отбор направлен на стабилизацию обильномолочности на уровне, достаточном для оптимального воспроизводства популяции в конкретных условиях среды, а искусственная селекция — на максимальную продукцию молока. Анализу взаимосвязей полиморфных систем с признаками приспособленности посвящены многочисленные исследования, которые дали противоречивые результаты. На наш взгляд, одной из основных причин неопределенности, кроме сложности механизмов генетической детерминации маркерных эффектов, является отсутствие эколого-генетического подхода при интерпретации результатов. Попытаемся показать это на примере взаимосвязей вариантов в-Сп с уровнем молочной продуктивности.
Известно, что компоненты молока синтезируются «на экспорт», иначе говоря, затраченные на синтез питательные вещества не могут быть повторно использованы организмом в качестве резервных или для покрытия возникающего дефицита. Показано, что полиморфные локусы, ответственные за синтез белков молока, тесно сцеплены. Генетические варианты белков имеют дифференцированную технологическую (20) и, следовательно, биологическую ценность для вскармливания приплода. Аминокислотные различия генетических вариантов белков в зависимости от экологических (например, кормовых) условий могут коррелировать с уровнем продуктивности и дифференциальной жизнеспособностью матери и приплода.
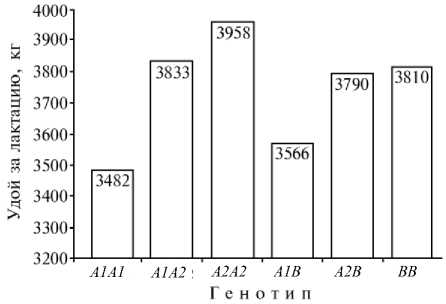
Молочная продуктивность у коров холмогорской породы в зависимости от генотипа по локусу β-казеина. Выборка сформирована из животных пяти ведущих племенных заводов по холмогорской породе из таежной зоны (Архангельская и Московская области, Республика Карелия).
Данные об удое за 305 сут первой лактации в зависимости от генотипов по в-Сп (получены на выборке, составленной из коров пяти ведущих племенных заводов по холмогорской породе из таежной зоны) (рис.) показывают, что максимальный удой был получен от особей-гомозигот с генотипом в-СпА2А2 , минимальный — с генотипом в-СпА1А1 . Гетерозиготы в-СпА1А2 уклонялись в сторону наиболее обильномолочного генотипа в-СпА2А2. В гетерозиготном сочетании с аллелем в-СпВ ранги маркерных эффектов в-СпА2 и в-СпА1 не менялись, но удои у животных соответствующих генотипов были ниже полусумм удоев альтернативных гомозигот (25). Результаты популяционного анализа взаимосвязей полиморфного и количественного признаков в целом совпали с результатами сегрегационного анализа в потомстве 11 гетерозиготных по локусу в-Сп ( А1А2 ) быков: у 64 дочерей с аллелем А1 удой и продукция молочного жира за лактацию составили соответственно 3716±99 и 139,1±3,8 кг, у 58 дочерей, унаследовавших аллель А2 , — соответственно 3930±103 и 150±4,1 кг. При оценке зависимости удоя коров за 305 сут первой лактации от их генотипа по локусу в-Сп в трех стадах из лесотундровой зоны ( п = 648) оказалось, что у животных с генотипами в-СпА1А1, в-СпА1А2, в-СпА2А2, в-СпА1В и в-СпА2В показатель составил соответственно 2306±65, 2450±86, 2374±120, 2245±173 и 2500±181 кг (различия между генотипами по удою недостоверны).
Эти данные и результаты, представленные на рисунке, свидетельствуют, что ранги удоев различных генотипов по локусу в-Сп в стадах лесотундровой зоны хорошо согласуются с рангами, установленными на выборке коров из пяти племенных заводов таежной зоны.
В данном случае независимо от того, что разница в удоях коров разных генотипов недостоверна, вероятность случайного совпадения результатов крайне низкая, поскольку они получены на двух рандомизированных репрезентативных выборках из двух субпопуляций, содержащихся в разных природных зонах и хозяйственных условиях.
Можно предположить, что дифференциация генотипов по удою связана с различиями кодируемых аллельными генами вариантов в-Сп по аминокислотному составу. Например, генетический вариант в-СпА1 отличается от в-СпА2 заменой одного остатка пролина на частично лимитирующий синтез белков молока гистидин. Для синтеза одной молекулы варианта β-казеина А 2 нужно на 20 % меньше гистидина, чем для синтеза варианта А 1 . Тогда при содержании скота на рационах, дефицитных по гистидину, повышенная частота Р-ОпА2 в стадах племенных заводов поддерживается одновременно естественным отбором и селекцией коров на обильномолочность. Однако такая гипотеза плохо согласуется с более высокой частотой в-СпА1 в стадах лесотундровой зоны, где естественный и искусственный отборы не только должны быть направлены против генотипа в-СпА1 , но их суммарная интенсивность должна превосходить давление аналогичных факторов в стадах племенных заводов.
Поскольку ранги удоев коров-аналогов по генотипам в-Сп в стадах племенных заводов и лесотундровой зоны совпадали, вероятно, направление векторов естественного и искусственного отборов по обильномолочности тоже совпадает. Возможно, в поколении лакти-рующих животных вектор направлен против аллеля в-СпА1 . Однако механизм, регулирующий частоты генов в популяции, более сложен, например не исключено, что он обусловливает дифференциальную выживаемость потомства различных генотипов на основании взаимосвязей мать—потомок. Теоретически в стадах племенных заводов элиминация аллеля в-СпА1, связанного с низкой продуктивностью, должна происходить интенсивнее, чем в стадах из зоны лесотундры, вследствие селекции по обильномолочности. В балансе сил, регулирующих частоту генов в стадах из лесотундровой зоны, скорее всего, возрастает вклад естественного отбора. При этом роль и вклад естественного отбора в поддержание внутрипородной дифференциации популяции по частотам аллелей в-Сп в зависимости от условий обитания существенно меняется и, по -видимому, выражается не только в дифференцированном эффекте аллелей в-Сп по уровню молочной продуктивности. Скорее всего, невыгодное с точки зрения селекции снижение удоя у генотипов в-СпА1А1 компенсируется более высокой выживаемостью потомства и достижением паритетной с альтернативными гомозиготами жизнеспособности за счет признаков, подконтрольных естественному отбору.
Вопрос о том, почему в стадах лесотундровой зоны частота, казалось бы, селективно невыгодного аллеля повышена по сравнению со стадами племенных заводов, пока остается открытым. В обсуждаемом случае генетико-автоматические процессы вряд ли можно рассматривать в качестве ведущего фактора, обусловившего сложившуюся геногеографическую ситуацию, поскольку выборка животных, представляющая холмогорский скот лесотундровой зоны, была сформирована из нескольких стад, репродуктивно изолированных между собой, но связанных опосредованно через заводскую популяцию. Анализ показал отсутствие упорядоченной динамики частот генов в зависимости от возраста (26).
Отметим, что распределение частот аллелей β-Cn в популяции холмогорского скота из Республики Коми согласуется с общей тенденцией распределения их частот в мировой популяции (хотя нельзя исключить случайность такого совпадения). Так, с юга на север в мировой популяции прослеживается увеличение частоты встречаемости β-CnA1 и снижение — β-CnA2 ; в мировой популяции индийского зебу частота встречаемости варианта β - CnA2 составляет 0,9 и более, а варианта β-CnA1 — не превышает 0,1; у североевропейских пород и пород низменностей черно-пестрого корня частота β - CnA2 снижается до 0,4-0,5; повышение частоты «селективно невыгодного» аллеля β-CnA1 установлено у многочисленных и наиболее обильномолочных пород крупного рогатого скота, хотя прямая селекция по генетическим вариантам β-казеина никогда не проводилась.
Лесотундровая субпопуляция на геногеографической карте холмогорской породы выделялась не только по структуре локусов, контролирующих полиморфизм белков молока (22, 25, 27) и групп крови. П.Н. Шубин (1968 год, персональное сообщение) у коров из совхоза «Большая Инта» (лесотундровая зона) обнаружил чрезвычайно редкий для европейских пород полиморфизм сывороточного альбумина и гемоглобина, который не был выявлен у чистопородного холмогорского скота в других природных зонах. Кроме того, в массиве чистопородный холмогорский скот из лесотундры отличался от скота племенных заводов высокой частотой TfЕ (24). Наконец, установленная нами ассоциация аллелей антигенов «простых» систем групп крови c аллелями несцепленного высокополиморфного В -локуса локально приурочена к той же природной зоне и субпопуляции. Накопленные факты позволяют сделать вывод о том, что распределение генотипов и частот генов в обследованной популяции холмогорского скота не согласуется с системой его разведения, поскольку длительный по времени и непрерывный поток генов из заводской популяции не нивелировал генетическую специфику массива чистопородного холмогорского скота лесотундровой зоны. Такую внутрипородную дифференциацию нельзя объяснить случайными причинами, поэтому повышенная частота аллелей β-LgA и β-СnА1 , а также ассоциированное распределение генов несцепленных локусов, контролирующих полиморфные группы крови, в региональной экзогамной субпопуляции могут служить индикатором селективного давления.
Выводы о вовлечении различных маркерных систем в селективные процессы, полученные в Приполярье, по существу сходны с выводами о популяционно-генетических последствиях акклиматизации и скрещивания зебу с местным скотом на Африканском континенте, сделанными при оценке распределения частот аллелей мтДНК в африканской популяции Bos taurus (28).
Таким образом, результаты многолетних исследований полиморфизма белков молока и антигенов групп крови в популяции скота холмогорской породы, расселенной в нескольких природно-климатических зонах в Республике Коми, подтверждают возможность разработки важного для практики способа генетико-популяционного прогноза перспектив акклиматизации популяций (пород) крупного рогатого скота в новых экологических условиях по этим маркерным системам.
Настоящая работа стала возможной благодаря бескорыстной помощи зоотехников-практиков В.В. Яковенко, Н.А. Артеевой, В.К. Доможирова, главного государственного инспектора Республики Коми в области племенного животноводства А.П. Захарова, за что автор выражает им искреннюю благодарность.
Л И Т Е Р А Т У Р А
-
6.
-
7.
А л т у х о в Ю.П. Генетические процессы в популяциях. М., 1989.
Л е в о н т и н Р. Генетические основы эволюции. М., 1978.
М а й р А. Популяция, виды и эволюция. М., 1974.
М а р и н к о в и ч Д., Т у ч и ч Н., К е к и ч В. Популяционно-генетическая изменчивость и способность к экологической адаптации. В сб.: Вопросы общей генетики. М., 1981: 34-45.
С о л б р и г О., С о л б р и г Д. Популяционная биология и эволюция. М., 1982.
Т о ц к и й В.Н., Х а у с т о в а Н.Д., А л ш и б л и Н.М. и др. Генетико-биохимические механизмы онтогенетической и филогенетической адаптации. Цит. и ген., 2002, 3: 69-75.
Ш м а л ь г а у з е н И.И. Факторы эволюции. М., 1968.
Г л а з к о В.И. Генетическая изменчивость при искусственном и естественном отборе и механизмы формообразовательного процесса. Журн. общ. биол., 1986, 4: 457-459.
О х а п к и н С.К., Д у н и н И.М., Р о ж к о в Ю.И. Селекция и эволюционный процесс. М., 1995.
Б о р о е в и ч С. Можем ли мы создавать сорта растений и породы животных по заранее разработанным моделям? В сб.: Генетика и благосостояние человечества. М., 1981: 154-166.
Ш у б и н П.Н., Е ф и м ц е в а Э.А. Полиморфизм белков и ферментов и оценка степени гетерозиготности генома у северного оленя. Тез. докл. Всес. сов. «Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих». Пущино, 1983: 206-207.
Временная инструкция по генетическому контролю достоверности происхождения сельскохозяйственных животных. М., 1983.
Правила генетической экспертизы племенного материала крупного рогатого скота. М., 2003: 4-15.
П о п о в Н.А., Е с к и н Г.В. Аллелофонд пород крупного рогатого скота по ЕАВ-локусу. Справ. католог. М., 2000.
S c h m i d t D.C. Starch-gel electrophoresis of κ -casein. Biochem. Biophys. Acta, 1964, 90(2): 411-414.
A s c h a f f e n b u r g R. Procedure for simultaneous phenotyping of β -casein andβ β -lactoglobulin variants in cows milk. J. Dairy Sci., 1967, 50(8): 1320.
М а т ю к о в В.С. Полиморфизм β -казеина у холмогорского скота. С.-х. биол., 1975, 10(3): 463-464.
Р о к и ц к и й П.Ф. Биологическая статистика. Минск, 1964.
С е р е б р о в с к и й А.С. Генетический анализ. М., 1970.
З а х а р о в И.А. Генетические карты высших организмов. Л., 1979.
С т р е к о з о в Н.И., С и в к и н Н.В., И о л ч и е в Б.С. Белковый состав молока и биохимический полиморфизм его фракций. Вест. РАСХН, 1996, 1: 52-53.
М а т ю к о в В.С., Ш у л ь г а Л.П. Геногеография холмогорского скота. Мат. Междунар. конф. Баренц-Евро-Арктического региона «Фундаментальные проблемы и перспективы развития». Петрозаводск, 1996: 102-103.
М а ш у р о в А.М. Генетические маркеры в селекции животных. М., 1980.
Ш у б и н П.Н. Изменение частоты аллелей трансферинового локуса у крупного рогатого скота Коми АССР. В сб.: Физиология и экология. Сыктывкар, 1969: 86-90.
М а т ю к о в В.С. Селекционный статус полиморфизма β -казеина у крупного рогатого скота. С.-х. биол., 1983, 12: 73-78.
М а т ю к о в В.С. Эколого-генетический анализ селективного значения полиморфизма β -казеина крупного рогатого скота. Вест. РАСХН, 2004, 3: 76-78.
М а т ю к о в В.С. Система β -лактоглобулина и генетическая адаптация крупного рогатого скота в условиях северо-восточной Европы. С.-х. биол., 1986, 15(5): 92-94.
M a c H u g h D.E., S h r i v e r M.D., L o f t u s R.T. e.a. Microsatellite DNA variation and the evolution, domestication and phylogeography of Taurine and Zebu Cattle (Bos tau rus and Bos indicus). Gen. Soc. of America, 1997, 146: 1071-1086.
Научно-исследовательский и проектно-технологический институт агропромышленного комплекса
Республики Коми,
167003 г. Сыктывкар, ул. Ручейная, 27,
