Об эволюции биоты в береговой зоне дальневосточных морей

Автор: Урусов В.М., Варченко Л.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 11, 2013 года.
Бесплатный доступ
В статье рассмотрено видообразование и возникновение эндемичных форм ниже видового ранга на российском Дальнем Востоке, связанное с зонами перехода океан-континент, лес-степь, субальпыгорные тундры, в которых химизм воздуха и почвы, солнечная радиация имеют особенности, поощряющие мутагенез. В результате в супралиторальных экосистемах и крупнотравных сообществах эндемов не менее 20 %, в субальпах от 14 %, в смешанных лесах дубравного генезиса 1,5-2 %. Причём часть эндемов супралиторали и ближайшей к ней береговой зоны обладает особенно ценными лекарственными, декоративными и пищевыми свойствами.
Супралитораль, субальпы, эндемизм, возраст эндемов, особый химизм, ионы морского происхождения в воздухе и почве
Короткий адрес: https://sciup.org/14082789
IDR: 14082789 | УДК: 581.524/527:574(571.6)
About biota evolution in the far east sea coastal zone
The speciation and the emergence of endemic forms below the species rank in the Russian Far East, associated with the transition zones ocean-continent, forest-steppe, sub-Alps-mountain tundra, where the air and soil chemism, solar radiation have peculiarities that encourage mutagenesis are considered in the article. As a result, in supralittoral ecosystems and large grass communities there are endems not less than 20 %, in sub-Alps from 14 %, in mixed oak genesis forests 1,5-2 %. Moreover, the part of endems in the supralittoral and the nearest coastal zone have a particularly valuable medicinal, ornamental and nutritional properties.
Текст научной статьи Об эволюции биоты в береговой зоне дальневосточных морей
Введение. Особый интерес представляет изучение микро- и макроэволюционных процессов на берегах дальневосточных морей с широким разнообразием сосудистых растений, таких, как карликовый вечнозелёный рододендрон родства Rhododendron sichotense , эндемичные крупноцветковые тимьяны и эдельвейсы у оз. Ханка, оз. Благодатное в береговой зоне Сихотэ-Алинского государственного заповедника в Приморье, на известняках в бассейне р. Партизанская (район Находки), крупноплодные Lonicera edulis ssp. kamtschatica на п-ове Большой Нос (охотоморская сторона), у подножия вулкана Атсонупури на о. Итуруп и в северном углу Озерновского залива на северо-востоке Камчатки, крупноплодные формы Rosa rugosa на о. Фуругельма (юг Приморья), Vaccinium yatabei в пихтарнике-черничнике на южном склоне вулкана Атсонупу-ри на высоте 250–400 м над ур.м. (Итуруп), крупношишечные формы кедрового стланика в Сахалинской и Магаданской областях и др. Это разнообразие перетекает в эндемичные расы (var.), подвиды (ssp.), например, можжевельников [17], и виды (sp.) не только у сосудистых растений. Общеизвестный меланизм у животных тоже выражен именно здесь. Зонирование морского влияния позволило установить сложность и результативность эволюционных преобразований именно в ближайшей к морю первой подзоне [10 и др.]. Во вторую очередь интересны экзотические экосистемы и виды, в особенности макротермные, приуроченные не столько к прибрежной зоне, в которой выделены 3 подзоны, сколько к следующей зоне – береговой, отстоящей от моря на 20–30 и даже 300 км и ограниченной в Приморье ледниковым рубежом океанического влияния (рис. 1). Именно к береговой зоне относятся важнейшие рефугиумы биологического разнообразия (БР) и наиболее богатые локальные флоры.
Первая подзона прибрежной зоны – подзона эдафических, экосистемных и динамических мозаик – наиболее приближена к береговой черте (даже на первые десятки метров) и находится под наиболее сложным влиянием акватории как на микроклимат, эдатопы и экотопы, так и динамику береговой линии и эволюцию биоты.
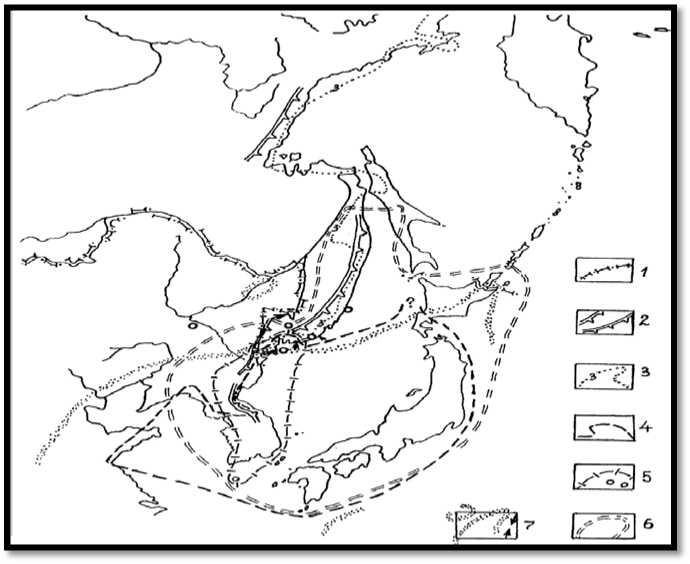
Рис. 1. Океаничность климата и ареалы лесообразователей, тяготеющих к ней. Границы и рубежи: России (1); океанического влияния в ледниковое время (2); коэффициента континентальности климата, равного 3 или меньше [15] (3).
Ареалы и изоляты Pinus densiflora (4), Abies holophylla (5), Taxus cuspidata (6), Quercus dentata (7)
Это не только современная полоса заплеска, но и её реликты, отстоящие от сегодняшней береговой черты иногда на 10-40 км, сложившиеся при более высоком (даже на 4 м) стоянии уровня Мирового океана 3,5 тыс. л.н. в суббореале и около 6 тыс. л.н. в атлантике. Экосистемы зоны заплеска в её широком понимании физиономически определяются не только супралиторальными видами, но и видами реликтовых береговых степей и предстепья Дальнего Востока России (ДВР). А поэтому к колосняку, осоке большеголовой, мер-тензии, хоризису, Rosa maximowicziana , R. rugosa , R. rugosa x R. davurica , эндемам супралиторали, включая виды родов Oxytropis, Artemisia, Dendranthema, Leontopodium, Thymus, добавляются злаково-разнотравно-полынно-тимьяновые фрагменты реликтовых степей, видимо, с Celastrus orbiculata, Fraxinus sieboldiana, F. densata, F. stenopterus . И всё же оригинальность биоты 1-й подзоны связана в основном с полосой шириной в 0,5-1,5 км, где и выпадает основная доля хлоридно-натриевых осадков и избирательно отмирает подрост [4, 5].
Цель исследований. Изучение видообразования и возникновения эндемичных форм на Дальнем Востоке.
Материалы и методы исследований . Материалы и методы относятся к сфере генэкологических исследований, выполненных авторами в береговой полосе и отчасти на верхней границе растительности в горах Приморья и Сахалина. Обращено внимание на частоту встречаемости и распределение форм с опушением листьев и побегов, белоцветковых форм на побережье и в высокогорьях. Причём, белоцветковый субальпийский Rhododendron bobrovii Д.Л. Врищ [21] выделен из круга близких Rh. sichotense рододендронов как раз по массовости его произрастания у верхней границы растительности: если бы это были отдельные образцы на тысячи кустов типичного Rh. sichotense, то можно было бы говорить об альбиносной форме. Однако перед нами массовый особенно низкий кустик с белыми цветками, субальпиец, частично поглощённый ценопопуляциями ультрабореального сихотинского рододендрона. Найдены многие критические признаки, разделяющие данные виды.
Ценность биоты 1-й подзоны прибрежной зоны для науки и практики в особых свойствах её таксонов и форм, т.е. полиморфизме, в лабильности, толерантности, урожайности, в возможности привлечь её для изучения микро- и макроэволюционных процессов, гибридизации, установления «адресов» возникновения таксономически ранжируемых новообразований, причём не только эндемичных, но и принадлежащих модифи- кационной изменчивости давно известных видов. Например, на морской террасе у оз. Благодатное в Тер-нейском районе Приморья карликовые особи Rhododendron sichotense в изобилии, однако особи с таким наследственно закреплённым свойством пока не выявлены – возраст данного конкретного экотопа вряд ли выше первой тысячи лет.
С этой проблемой столкнулись как первоисследователи природно-ресурсного потенциала ДВР, который в те отдалённые времена был гораздо обширней географически, так и непосредственно природополь-зователи. И проблема не только в прямом влиянии морей, их ледовитости, направлении ветров, горных барьеров на их пути, но и, например, разнообразии эволюционных факторов, определяющих формирование и уцелевание БР, и в хозяйственной отдаче видов и экосистем, необходимых режимах их эксплуатации и охраны. Это относится и к арборифлоре и к флоре конкретных урочищ, и к наземной и морской фауне, и к природопользованию в целых долинах впадающих в море рек, если длина рек до 100 км (Восточное Приморье). Например, экранируемая от выноса воздушных масс с Японского моря средневысокогорным хр. Партизанский долина р. Партизанская, почти перпендикулярная летнему муссону, практически до устья перспективна для сельскохозяйственных культур и садоводства при сдерживающем влиянии наводнений.
Результаты исследований и их обсуждение . И.С. Майоровым с соавторами [10] макрозона берегов юга ДВР рассматривается как экотон прибрежных и береговых акваторий и территорий, в котором достаточно чётко выражено в т.ч. четвертичное видообразование. Причём на суше выделены 2 зоны (прибрежная современных и береговая убежищ БР и, возможно, реликтовых океанических влияний). В прибрежной зоне 3 подзоны – современных влияний моря, узкая; реликтовых влияний моря, связанная с его высоким стоянием в тёплые эпохи голоцена; выноса морских туманов. А береговая зона вбирает в себя убежища макротерм-ных видов [8, 22] и простирается даже до главных рубежей океанического влияния.
В последнем стадиале, ледниковье, этими рубежами были Восточно-Маньчжурские горы, главный водораздел Сихотэ-Алиня и хр. Джугджур. Восточноазиатская флористическая область акад. А.Л. Тахтаджяна скукоживалась, а её северная граница смещалась с 52º с.ш. на юг до 44º (рис. 2). В этом и более раннем погружении суши основная причина широкого распространения совмещённой таёжно-широколиственной растительности в регионе [15, 16].
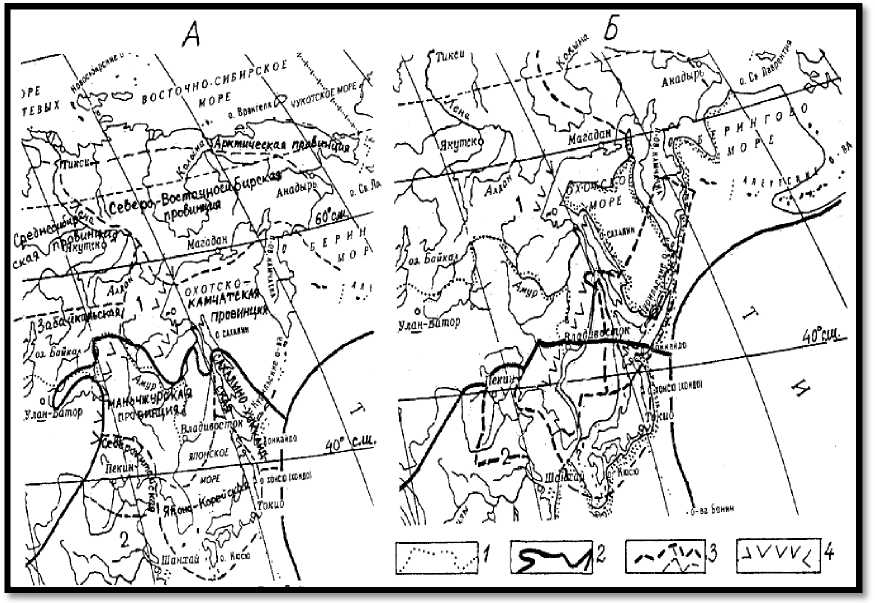
Рис. 2. Современные (а) и позднеплейстоценовые (ледниковые) с учётом регрессии моря и субконтинентализации (б) флористические области, провинции и рубежи океанического влияния.
Границы: 1 – России; 2 – флористических областей; 3 – флористических провинций;
4 – рубежи океанического влияния
В текущем межстадиале рубежи (за исключением Джугджура) сместились на запад, к Большому Хин-гану и Буреинским горам [13 и др.], т.е. в вюрме – 12–16 тыс. л.н. – океаническое влияние в Восточном Сихотэ-Алине распространялось на полосу не шире 100–150 км от современной береговой черты и только на западе Уссурийского, Октябрьского, Пограничного районов Приморья проникало в глубину материка до примерно 200–250 км. С довюрмской полосой океанического влияния в Сихотэ-Алине и Восточно-Маньчжурских горах и сейчас увязаны находки видов сахалинского генезиса, в частности, изолятов пихты сахалинской Abies x sachalinensis , зонально-ландшафтных сахалинских пихтарников на юго-востоке Хабаровского края [17], макротермных экзотов северокорейских и субтропических восточно-китайских лесов [18, 19].
Эндемиками здесь являются не менее 40 видов сосудистых растений, не менее 25 видов из них эндемичны для Восточного Сихотэ-Алиня [18, c. 32]: Festuca vorobievii, Silene olgae, Rosa maximowicziana, Potentil-la tranzschellii, Oxytropis mandshurica, O. ruthenica, Peucedanum ( Kitagawia ) litorale , несколько видов Thymus , возможно, апомиктных по генезису [12], Anaphalis pterocaulon, Heteropappus saxomarinus, H. villosus, Dendran-thema coreanum. Всего на морских берегах юга ДВР из более чем 200 видов сосудистых растений преимущественно степного и дубравного генезиса, встречающихся в зоне заплеска, облигатными и близкими к ним являются почти 100. Среди них немало полиплоидов [14], которые мы относим к особенно лабильным и вовсе не всегда геологически молодым. Самыми молодыми на супралиторалях ДВР являются Juniperus confer-ta (Сахалин, видимо, сформирован в эоплейстоцене или к рубежу плейстоцена, потому что похолодания позднего плейстоцена позволили ему расселиться по внешней гряде дюн как Западного, так и Восточного Сахалина, вдоль всего побережья Японского моря в Японии и Корее и пройти на берег Жёлтого моря в КНДР) [20, с. 291], J . x coreana = J. conferta x J. sibirica (голоценовый гибрид на береговых валах Сахалина и Кореи), Rosa rugosa x R. davurica , R. marretii x R. davurica (берега севера Сихотэ-Алиня, голоцен), подвиды можжевельников [17], возраст которых от раннечетвертичного до среднепозднечетвертичного (плейстоценового). Причём если эндемы высокогорий имеют преимущественно меловой и олигоценовый возраст, неоэн-демы Камчатки и Курил – эоплейстоценовый, то подвиды современной и реликтовой береговой зоны сформированы около 500–400 тыс. л.н. [15, 16, 17] (рис. 3).
Чаще всего особенно молоды гибриды. Некоторые таксоны обязаны своим происхождением погружению окраины Азии, например, Dendranthema coreanum , Sabina davurica ssp . maritima , может быть, немалое для Северной Пацифики в целом число Artemisia, Leontopodium, Saussurea . Погружение в сумме составило 1000–1500 м и завершилось к рубежу плейстоцена [16].
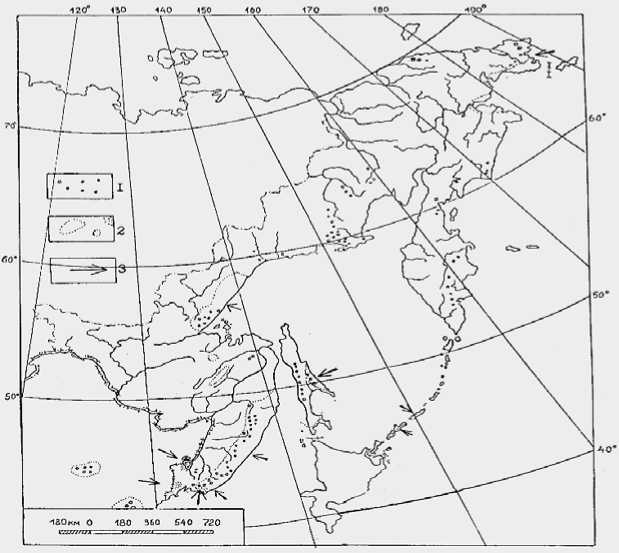
Рис. 3. Основные районы распределения на Дальнем Востоке эндемичных родов (I) и видов (2) сосудистых растений. Зоны, перспективные на открытие новых эндемичных видов (3)
Отметим и вот что: в оказавшихся из-за тектонического погружения у уровня моря популяциях Sabina sargentii (юг Сахалина, о-в Кунашир, Итуруп, Монерон (РФ), часть Хоккайдо (Япония)) как бы произошло возвращение к большей требовательности к теплу, а отчасти в ходе эволюции большинством сабин преодолённой однодомности, что позволяет предположить продолжительную дивергенцию береговых и высокогорных популяций данного вида. В этом есть хозяйственный смысл. И по крайней мере для альпинариев средних широт однодомная форма сабины перспективна.
В чём же причина ускоренных мутагенеза и микро- и макроэволюционных процессов именно в этой подзоне береговой зоны? Во-первых, это разнообразие и широчайшая амплитуда климатических факторов; во-вторых, это особый и разнообразный во времени и пространстве химизм воздуха и почвы; в-третьих, радиационный фактор; в-четвёртых, постоянное наличие свободного для поселения новообразований пёстрого по составу и структуре набора эдатопов, сопоставимое с имеющимся на верхней границе леса, где также особенно многочисленны мутации, например, у Abies sibirica [6] и почти также выражены свободные участки для поселения новых форм; в-пятых, как и на верхней границе растительности прогорающие участки здесь чередуются с уцелевающими, в той или иной степени изолированными и не преобладают, а сами пожары редки и относительно слабее действуют на биоту (потому что здесь, как правило, меньше сухих растений и ветоши); в-шестых, здесь более высоко разнообразие эдатопов и экотопов; в-седьмых, всегда в наличии разнообразие физических и химических барьеров, изолирующих как экотопы, так и участки берега в некоторой степени аналогично имеющему место на верхнем пределе растительности. Вот почему Д.Л. Врищ [1], описав в своё время как самостоятельный вид Lilium sachalinensis с берегов Сахалина и Курил (зона приморских дюн), считает вероятной наследственную закреплённость карликовости у высокогорного изолята L. pensylvanicum на горе Снежная в Чугуевском районе Приморья [21, с. 56]. И здесь же удалось найти совсем древний субальпиец Rhododendron bobrovii - вечнозелёный карликовый вид с цветочными почками, равномерно распределёнными по всей ветви. Он наследует автохтонный высокогорной флоре той эпохи, когда Сихотэ-Алинь был выше на 1-1,5 км [2].
Теперь мы понимаем, что эндемизм определяется интенсивностью мутагенеза (или частотой мутаций за определённый временной интервал на определённое количество организмов данного вида) и мощностью популяционных волн, реализующихся как во времени, так и в пространстве, например, увеличивая долю новообразований. Если насыщенность эндемичными видами экосистем супралиторали и высокогорий выше в разы общего эндемизма флоры Приморья (около 6 %), то можно говорить о бесспорно лидирующем становлении новых узкоареальных форм как раз в зонах перехода океан-континент и субальпы-высокогорная тундра в том числе и потому, что высокий эндемизм крупнотравной зоны не более чем наложение условий высокогорья на изначально лесную растительность, в силу особых причин сдвинутую в микротермную зону, однако при достаточных влажности и тепле корнеобитаемого слоя почвы зимой [19 и др.].
Итак, на супралиторали новообразования формируются за 300-400 тыс. лет на уровне подвидов и полувидов, за 1 млн лет - молодых видов и хороших гибридных видов. Эндемы крупнотравья Курил и Сахалина, а также Камчатки и Японии, сложились примерно за 2,5 млн лет при единстве суши Азии от Командор до Японии. Иное дело эндемы верхнего предела растительности, возраст которых может быть олигоценовым, а при уцеле-вании на древнем кольце обрамления Японского моря ( Sabina sargentii ) - меловым. Это эндемы, маркирующие древний периметр конкретных морфоструктур центрального типа - МЦТ [9, 20 и др.]. Они, как правило, старше эндемов отдельных горных стран внутри МЦТ 3-го порядка и характерных преимущественно внутренним зонам МЦТ ландшафтных лесообразователей, к которым на юге ДВР относятся, прежде всего, Abies holophylla , Pinus koraiensis (низкогорья), A. nephrolepis , Picea komarovii (среднегорная тайга) (рис. 4-5).
И всё же вполне вероятно, что в зонах перехода главным фактором эволюции является стрессирова-ние физиологии генеративных процессов на уровне ценопопуляций видов в особых, а именно крайних, периферийных условиях среды. Напомним, что «наиболее распространено, вероятно, смещение ниш у относительно генерализованных видов к периферии пространства ресурсов у границ ареала, подтверждением чему служат частые случаи интенсивного формообразования в периферических изолятах» [23, с. 171-172]. А предоставляющие ресурсы для «центробежно направленного действия отбора» [20] зоны не ограничиваются контактом континента и океана: физические (излучения разных типов, понижения и повышения температуры и влажности в т.ч. с высокой скоростью + стрессирование физиологии), физико-химические за счёт действия особых минералов и субстратов, химические (полихлорбифенилы, азотистая кислота и другие канцерогены), биологические, включая вирусы, мутагены действуют очень активно и в высокогорьях и в береговых условиях, к ним приближённых вследствие контакта с надолго замерзающими морями, солоноватыми и пресными обширными водоёмами. Если общий фон мутаций определяется динамизмом солнечной активности и радиации, то учащение мутаций в зонах перехода, таких явлений, как выраженный именно здесь меланизм у животных, стланиковость, сизый налёт, интенсивная опушенность поверхности всех частей организма иногда даже вместе у растений, вызываются другими факторами - химическими мутагенами, стрессируемой физиологией репродуктивного процесса. По крайней мере, планетарной динамикой мутаций и особым разнообразием эдатопов это не объяснить.
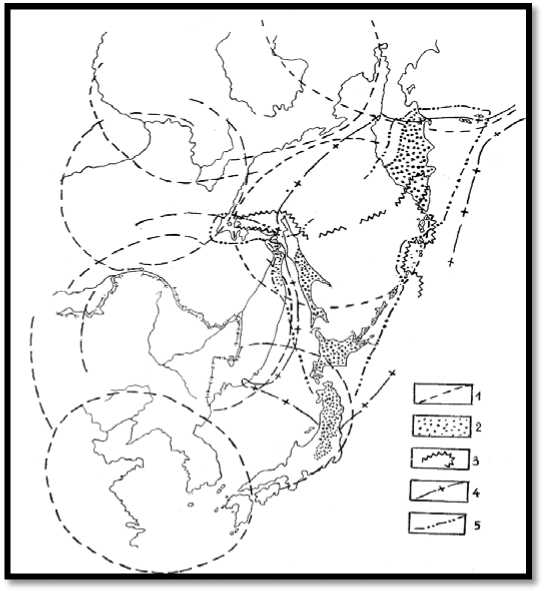
Рис. 4. Крупнотравные сообщества ДВР и ареалы некоторых связанных с ними видов: 1 – контуры основных МЦТ; 2 – зона распространения крупнотравных лугов; 3 – ареалы Lysichiton kamtschatcense;
4 – Filipendula camtschatica; 5 – Heracleum dulce
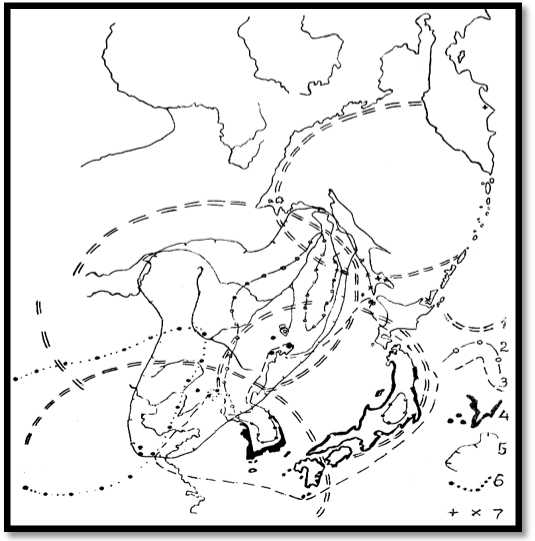
Рис. 5. Морфоструктуры центрального типа способствуют формированию берегового и высокогорного эндемизма в своих пределах, гибридизации в зонах сближения и контактов контуров морфоструктур и выживанию характерных видов БР в их центральных зонах: 1 – контуры МЦТ 3-го порядка
А.П. Кулакова; 2 – ареалы и изоляты Pinus koraiensis; 3 – P. Densiflora; 4 – P. Thunbergiana; 5 – Quercus mongolica; 6 – Pinus tabulaeformis; 7 – изоляты Abies gracilis (не путать с A. x sachalinensis) и гибридные популяции Quercus mongolica x Q. crispula
В.Ф. Максимова [11] вслед за А.Н. Качуром [4, 5 и др.] по характеристикам древостоев берегов Среднего Сихотэ-Алиня выделяет прибрежную (0-5 км), переходную (5-25 км), континентальную (25-50 км) зоны. Причём «формообразующее влияние моря на древесную растительность - кустарниковая форма роста установлена для узкой полосы шириной 70-100 м». Ионов морского происхождения - хлора и натрия на удалении 2 км от моря выпадает в 2-8 раз больше, чем в следующей полосе [4, 5 и др.], вызывая снижение продуктивности и жизненности даже у дуба монгольского. Вплоть до отмирания надземной части его возобновления (вот и причина «кустарниковой формы роста» дуба у моря, вернее, одна из причин), а так как подрост других пород хлоридно-натриевые и хлоридно-кальциевые осадки, по крайней мере, в отдельные годы полностью уничтожают, то не обязательно человек является «автором» береговых дубняков. Именно поэтому наиболее крупными вкладчиками в эндемизм флоры ДВР являются даже не высокогорья на верхнем пределе растительности, а берега окраинных морей, а это супралиторально-луговой и отчасти лесной и лугово-пойменный комплексы эндемов А.Е. Кожевникова [7], которые мы бы назвали супралиторально-степными эндемами, а также аркто-монтанный комплекс эндемов этого же автора и комплекс эндемов круп-нотравья [18 и др.]. В альпийском поясе средних широт Евразии видообразованию способствует понижение верхнего предела леса в связи с падением тепла или ростом абсолютных высот гор, что и сформировало ,в частности, альпийскую луговую зону и её эндемизм. Но на ДВР похолоданиям позднего плейстоцена предшествовало такое тектоническое погружение суши, что и в стадиалы верхняя граница растительности оставалась прогрессивной. И микроэволюция здесь замедлялась по сравнению с высокогорьями Кавказа, Алтая, Саян. Зато у моря осыхание шельфа в связи с понижением уровня Мирового океана на 120-130 м (вюрм!) обусловило экотон экотопов с новыми микроклиматами, стимулируя адаптивную эволюцию [19].
При общем уровне эндемизма около 6 % (у А.Е. Кожевникова 10,8 %) эндемов супралиторального комплекса и береговых скал около 20 % от общего объёма данного флороценотипа (для гольцев Сихотэ-Алиня И.Б. Вышиным [3] выявлен 14 %-й эндемизм), крупнотравного флороценотипа до 25 % [18, с. 36] при 10 %-м эндемизме флоры крупнотравных лугов, а также 6-10 % эндемов на известняковом хребте Лозовый (древний коралловый риф в Партизанском и Надеждинском административных районах Приморья) и 1-2 % эндемизме в экосистемах чернопихтарников и кедровников ДВР. Следовательно, в занимающих не более чем 1 % суши в зонах перехода мутагенез результативней в разы, а мутации случаются на порядки чаще. Добавим к этому наличие свободных для заселения субстратов.
Выводы
-
1. Эндемизм сосудистых растений ДВР на супралиторали и в целом в скально-береговой зоне примерно вдвое выше высокогорного и предполагаемого нами для древних известняковых рифов низкогорий, например, известняковых хребтов юга Приморья. Это логично увязать с большим разнообразием биохимических воздействий на генеративную сферу и жёсткостью мутагенеза и популяционных волн именно здесь.
-
2. Приходится согласиться с выводами ученых Н.С. Пробатовой и В.П. Селедца, что повышенный уровень плоидности является важным, если не важнейшим фактором формирования эндемов в береговой зоне региона и не обязательно молодых эндемов.
-
3. Высокий (даже высочайший - до 25 %) эндемизм среди сосудистых растений крупнотравного флороценотипа и крупнотравных лугов как особого типа растительного покрова, кроме ДВР свойственный границе леса в некоторых горных странах Азии (от 10 %), доказывает уникальность набора факторов-движителей эволюции, сложившихся к квартеру в пределах крупнотравной области региона.
-
4. Формирование эндемичных подвидов именно в современных, раннечетвертичных и позднеплейстоценовых экосистемах супралиторали и во вторую очередь на верхнем пределе растительности открывает возможность выявления здесь материала и облегчает построение моделей генерирования перспективных растительных форм и сортов.
-
5. Совпадающие черты эволюции до уровней подвида и вида микротермных форм, например, у овсяниц, кипарисовых, можжевеловых, тисовых, лилий, клёнов, рододендронов, жимолостей, вейгел, дендран-тем, тимьянов ДВР в зонах контакта континент-океан, лес-степь, лес-субальпы подтверждают общность действующих факторов мутагенеза у моря и высоко в горах.
