Об участии холестерола, прогестерона, кортизола и липопротеинов в возрастных изменениях обмена веществ у цыплят-бройлеров промышленного кросса

Автор: Колесник Е.А., Дерхо М.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Птицеводство: наука и технологии
Статья в выпуске: 4 т.52, 2017 года.
Бесплатный доступ
По И.И. Шмальгаузену, на морфофизиологические изменения особи влияют факторы внешней среды, причем специфика реакций организма зависит от нормы его реакции и стадии онтогенеза, на которой действуют раздражители. Также известно участие гормонов в регуляции обмена веществ, роста и развития, в процессах адаптации. Мы определили концентрацию липопротеинов высокой (HDL) и низкой плотности (LDL) (непосредственно вовлечены в белковый и липидный обмен), общего холестерола (TCS), прогестерона (P4), 17-гидроксипрогестерона (17-OHP), кортизола (Cortisol) в плазме крови у цыплят-бройлеров кросса Hubbard F15 на ранних стадиях постнатального онтогенеза (четыре группы по 10 особей в каждой, возраст птицы по группам - 1, 7, 23 и 42 сут; промышленное стадо ООО «Чебаркульская птица», Челябинская обл., Россия). Роль и взаимосвязи этих факторов в метаболизме оценивали с помощью корреляционного анализа по Пирсону и факторного анализа методом главных компонент (метод вращения факторов - Varimax). Так, в возрасте 1 сут отмечали интеграцию факторов, вовлеченных в обменные процессы и регуляцию метаболизма, - HDL, LDL, TCS, P4 и Cortisol (r-Pearson: P4 и Cortisol - r = 0,69, p = 0,027; P4 и TCS - r = 0,82, p = 0,004; HDL и LDL - r = 0,83, p = 0,003; HDL и TCS - r = -0,67, p = 0,033). На 7-е сут у бройлерной птицы главными компонентами были прогестерон и кортизол (r-Pearson: P4 и Cortisol - r = 0,73, p = 0,016) и проявился фактор донорства холестерола с ведущими элементами LDL и TCS ( r = 0,73, p 4 ( r = -0,88, p 4 ( r = 0,76, p
Прогестерон, 17-гидроксипрогестерон, кортизол, липопротеины высокой и низкой плотности, холестерол, бройлерные цыплята, ранний постнатальный онтогенез, гомеостаз
Короткий адрес: https://sciup.org/142214164
IDR: 142214164 | УДК: 636.52/.58:591.1:577.121.9 | DOI: 10.15389/agrobiology.2017.4.749rus
Involvement of cholesterol, progesterone, cortisol and lipoproteins in metabolic changes during early ontogenesis of broiler chicks of an industrial cross
Morphophysiological changes in body are influenced by environmental factors, what is more, the specific nature of the body reactions depends on the reaction rate and the stage of ontogenesis on which the physiological stimuli act (I. Schmalhausen, 1982). Also it is known that hormones participate in the regulation of metabolism, growth and development, in adaptation processes. We determined the concentration of high-density (HDL) and low density (LDL) lipoproteins directly involved in protein and lipid metabolism, total cholesterol (TCS), progesterone (P4), 17-hydroxyprogesterone (17-OHP), and cortisol in blood plasma of Hubbard F15 broiler chickens at early postnatal ontogenesis using four groups of poultry of the industrial herd of Chebarkulskaya Ptitsa LLC (Chelyabinsk Province, Russian Federation), 10 animals in each, aged 1, 7, 23 and 42 days, respectively. The role and interrelations of these substances in metabolism were assessed using Pearson's correlation analysis and factor analysis by the principal components method with Varimax factor rotation. Thus, in 1-day aged chicks, the integration of factors HDL, LDL, TCS, P4 and cortisol involved in metabolic processes and metabolism regulations was noted with r -Pearson for P4 and Cortisol at r = 0.69, p = 0.027; for P4 and TCS at r = 0.82, p = 0.004; for HDL and LDL at r = 0.83, p = 0.003; and for HDL and TCS at r = -0.67, p = 0.033. On day 7 day, the principal components were progesterone and cortisol ( r -Pearson for P4 and cortisol of r = 0.73, p = 0.016), and a cholesterol donation factor with LDL and TCS as the leading elements ( r = 0.73, p < 0.05). In 23-day-old chicks the components which have become principal were HDL ( r = 0.91, p < 0.05) and 17-OHP ( r = 0.74, p < 0.05), which we attribute to growth, and also P4 ( r = -0.88, p < 0.05) and cortisol ( r = -0.77, p < 0.05) viewed as regulatory ones. On day 42 we revealed cholesterol donation factor ( r -Pearson of r = 0.86, p = 0.002 for LDL and TCS) and an integral factor with the principal components HDL ( r = 0.74, p < 0.05), P4 ( r = 0.76, p < 0.05) and cortisol ( r = 0.84, p < 0.05). Thus, here we described the age-specific features of interaction between lipoproteins and hormones of cholesterol-progesterone-cortisol system involved in broiler metabolism, and found out the change of principal components and functional relationships among the hormones of progesterone group and lipoproteins during early growth, which, according to our thought, makes a physiological basis for chicken performance under commercial poultry production. In further studies, it can be reasonable to assess the role of these hormones and metabolites in the control of reaction norm and adaptive capability of broilers, and the physiological cost of adaptation (i.e. adequate or pathological response) to reproduction at commercial farms.
Текст научной статьи Об участии холестерола, прогестерона, кортизола и липопротеинов в возрастных изменениях обмена веществ у цыплят-бройлеров промышленного кросса
Еще И.И. Шмальгаузен отмечал, что морфофизиологические изменения напрямую связаны с действием факторов внешней среды на совокупность процессов развития (1). Ответ организма на внешнее воздействие становится своего рода раздражителем, обусловливающим последующие реакции, но специфика формообразовательных реакций зависит прежде всего от самого организма, его нормы реакции и этапа онтогенеза, на котором действуют раздражители (1).
Гомеостаз, который объединяет циклы синтеза и распада химических веществ в жизнеспособную морфофункциональную структуру открытой системы — организма (2), можно определить как поддержание относительного динамического постоянства внутренней среды на основе взаимосвязанных системных регуляторных и метаболических процессов с участием факторов эндогенной и экзогенной природы, определяющих онтогенетическое развитие (3, 4). У бройлерных цыплят на ранний неонатальный период развития приходятся критические стадии, когда формируются физиолого-биохимические механизмы, обеспечивающие впоследствии ус- тойчивое функционирование и адаптационные возможности организма в условиях промышленных технологий (3, 5-7). Основой гомеостаза, сформированной в процессе эволюционного развития вида, и первичным связующим звеном между эндо- и экзогенными энергетическими и пластическими потоками, сигнальными воздействиями служит эндокринная регуляция. Она включает иерархическую цепь гормонов, содержание которых в крови по типу обратной связи определяет, в том числе, количество холестерина и липопротеинов крови, отражающее как процессы синтеза гормонов надпочечников, так и их регуляторные эффекты (2, 8-10). Один из таких гормонов — прогестерон, синтезируемый из холестерина липопротеинов низкой плотности, наиболее известен участием в эндокринном контроле формирования, роста и развития плода. К основным циркулирующим метаболитам прогестерона и ведущим предшественникам стероидов относится 17-гидроксипрогестерон (11, 12). Недавно установлено что прогестерон и его синтетические производные оказывают прямое (за счет нейростероидов группы прегненолона, синтезируемых de novo из холестерола в головном мозге птиц) (12) и опосредованное (13, 14) воздействие на вегетативный и центральный отделы нервной системы, включены в нейромедиаторный контроль хронологических ритмов постнатального роста и развития за счет рецепторного взаимодействия с γ-аминомасляной кислотой (12, 13) и дофамином, влияют на органогенез сосудов, опорно-двигательного аппарата (15), скелетной мускулатуры, имеющей гипертрофированное развитие у цыплят-бройлеров (5, 9), миелинизацию нервных волокон (13). Собственно прогестерон, 17-гидроксипрогестерон и его метаболит кортизол — одни из первичных активаторов и эффекторов гипоталамо-гипофизарно-адреналовой оси (10, 16). Тем не менее, информация о роли прогестерона и его производных в липидном и белковом метаболизме (несмотря на прямую связь биосинтеза этих метаболитов, в частности липопротеинов низкой и высокой плотности, и гормонов) крайне ограничена. Имеющиеся сведения в основном получены в медицине, тогда как в отношении сельскохозяйственных животных, где подобные исследования имеют важное практическое значение, данные единичны, разрознены или почти отсутствуют.
Мы впервые провели системное исследование содержания липопротеинов высокой и низкой плотности (непосредственно вовлечены в обмен белков и липидов), общего холестерола, прогестерона, 17-гидроксипрогес-терона, кортизола в плазме крови у цыплят кросса Hubbard F15 в возрасте 1, 7, 23 и 42 сут в связи с параметрами роста и развития и проанализировали эти биохимические показатели методами корреляционного анализа и главных компонент (факторный анализ). Выявлены возрастные закономерности изменений в концентрации этих метаболических и регуляторных факторов, что может отражать смену их роли в процессе роста и адаптации.
Целью работы было изучение динамики содержания ряда метаболитов и гормонов (липопротеинов, холестерина, прогестерона и его производных) в плазме крови бройлеров с учетом изменения массы тела для оценки возрастных изменений этих факторов и их роли в онтогенезе.
Методика. Эксперименты выполняли на бройлерах кросса Hubbard F 15 (ООО «Чебаркульская птица», Челябинская обл.), из которых в цехе выращивания (клеточное содержание) по принципу сбалансированности сформировали группы P1, P7, P23, P42 (в каждой n = 10; возраст птицы по группам — 1, 7, 23 и 42 сут). Кормление и содержание осуществляли согласно зоогигиеническим нормам без ограничений в доступе к корму (17).
Кровь для исследования получали при декапитации птицы в 1- и 7-суточном возрасте с соблюдением принципов гуманности, изложенных в 750
директивах Европейского сообщества (86/609/ЕЕС) и Хельсинкской декларации, и прижизненно пункцией подкрыльцовой вены вакуумным методом у 23- и 42-суточных цыплят. В плазме, стабилизированной EDTA, определяли общий холестерол (TCS), липопротеины низкой плотности (LDL), липопротеины высокой плотности (HDL) — ферментативным методом с помощью коммерческих наборов Вектор-Бест (Россия) и Ольвекс Диагностикум (Россия) (18, 19); гормоны — методом твердофазного имму-ноферментного анализа (ИФА) с тест-системами сэндвич-типа («XEMA Co., Ltd», Россия) Прогестерон-ИФА К207 для прогестерона (Pregn-4-ene-3,20-dione, P4) (20), 17-ОН-Прогестерон-ИФА К217 для 17-гидроксипро-гестерона (17-OHP) и Кортизол-ИФА К210 для кортизола (Cortisol) (11). При анализе на гормоны образцы инкубировали в термостатируемом шейкере ELMI Sky Line Shaker ST-3 («ELMI, Ltd», Латвия), оптическую плотность измеряли на фотометре для иммуноферментных тест-систем MINDRAY MR-96A Elisa Microplate Reader («MINDRAY Ltd», КНР).
Определяли среднесуточный прирост массы тела за возрастные периоды: Ассп = (W1 - W0)/(Т1 - Т0), где W0 и W1 — масса тела в начале и конце учетного периода (г) в возрасте Т0 и последующем возрасте Т1 (сут).
Для идентификации структуры взаимосвязей анализируемых элементов выполняли корреляционный анализ по Пирсону (Pearson Correlation, r-Pearson) и факторный анализ (Factor Analysis) (21) нормально распределенных в исследуемой выборке значений биохимических параметров (STA-TISTICA 8.0, «StatSoft, Inc.», США). Факторы выделяли методом главных компонент (Principal Сomponents, для вращения факторов применяли метод Varimax) (21). Цифровые данные представлены средней арифметической ( X ) и стандартной ошибкой средней (±SEM). Степень и достоверность различий для полученных результатов оценивали с помощью параметрического t -критерия Стьюдента в программе STATISTICA 8.0. Различия считали статистически значимыми при р ≤ 0,05.
Результаты. К 7-м сут кривые изменения концентрации TCS и ли-
1. Динамика биохимических показателей крови и массы тела у бройлерных цыплят кросса Hubbard F15 в раннем постнатальном онтогенезе ( X ±SEM, n = 10, ООО «Чебаркульская птица», Челябинская обл., 2014 год)
(см. табл. 1). Динамика концентрации 17-OHP и приростов массы тела у бройлерных цыплят имела достоверное сходство с пиком максимальных значений в возрасте 7-23 сут, а среднесуточный прирост массы тела оказался наибольшим на 23-и сут — 305,32 % (p < 0,001) (см. табл. 1). Кроме HDL, к 42-м сут содержание 17-OHP восстанавливалось до такового в возрасте 1 сут, а абсолютное значение суточного прироста живой массы за период с 23-х по 42-е сут было наиболее высоким, хотя оставалось ниже референтного, которое зарегистрировали с 7-х по 23-и сут (305,32 %, p < 0,001), составляя только 158,62 % (p < 0,001) (см. табл. 1).
Мы рассчитали взаимные корреляции (в том числе с учетом корреляций по Пирсону) и определили факторы и соответствующие главные компоненты среди изученных биохимических показателей в разные периоды роста и развития бройлеров (табл. 2).
-
2 . Результаты факторного анализа биохимических показателей крови у бройлерных цыплят кросса Hubbard F15 в раннем постнатальном онтогенезе с использованием метода главных компонент
|
Показатель |
Возраст, сут |
|||||
|
1 |
7 |
23 |
42 |
|||
|
фактор |
||||||
|
1 |
1 |
2 |
1 2 |
1 |
2 |
|
Липопротеины:
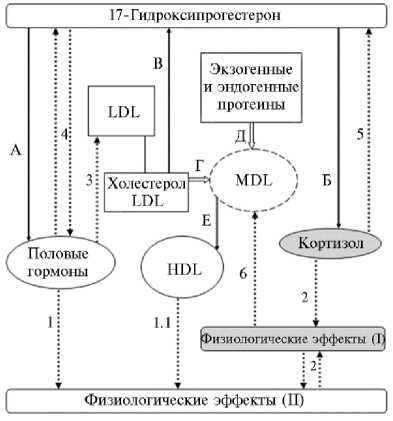
Схема, отражающая вовлеченность холестеро-ла, прогестерона, кортизола и липопротеинов в процессы роста и развития у цыплят-бройлеров кросса Hubbard F15: LDL — липопротеины низкой плотности, MDL — липопротеины промежуточной плотности, HDL — липопротеины высокой плотности; А, Б и В — синтез соответственно половых гормонов, кортизола и 17-гидроксипрогестерона; Г, Д и Е — конверсия липопротеинов низкой плотности и протеинов в липопротеины высокой плотности; 1 и 1.1 — регуляция обеспечения процессов роста и развития организма пластическими веществами, 2 — взаимная регуляция процессов адаптации, роста и развития, 3 — регуляция обмена LDL, 4 — регуляция метаболизма 17-гидрок-сипрогестерона и половых гормонов, 5 — регуляция превращения 17-гидроксипрогес-терона, 6 — регуляция синтеза HDL. Процессы биосинтеза обозначены сплошными линиями, регуляции — пунктирными.
Методом главных компонент у бройлерных кур в возрасте 1 сут мы выявили один общий фактор, за периоды с 7-х по 23-и и с 23-х по 42-е сут жизни — по два фактора (см. табл. 2). На основании полученных результатов и данных литературы нами предложена обобщенная схема взаимодействия изученных гормонов и метаболитов (рис.). На схеме процессы, в которые вовлечены эти компоненты, разделены на синтетические и регуляторные, а их эффекты проявляются последовательно: на текущей стадии развития формируется физиолого-биохимическая платформа (процессы, обозначенные нами как приспособительные; физиологические эффекты I), обеспечивающая рост и развитие на следующем этапе (физиологические эффекты II). В соответствии с данными таблицы 2 и схемой, факторы, которые выделились для каждого из изученных периодов, включали главные компоненты, отнесенные нами в основном либо к приспособительным, либо к ростовым, или совокупный фактор, объединяющий компоненты с приспособительной и ростовой функцией (см. табл. 2, рис.). Так, у цыплят в возрасте 1 сут выявлен интегративный фактор обменных и приспособительных процессов, включающий гормональные и метаболические компоненты (см. табл. 2). Это характеризует их существенную взаимосвязь (r-Pearson: P4и Cortisol — r = 0,69, p = 0,027; P4и TCS — r = 0,82, p = 0,004; HDL и LDL — r = 0,83, p = 0,003; HDL и TCS — r = -0,67, p = 0,033) в самый ранний постнатальный период, когда обмен веществ еще полноценно не сформировался (3, 5, 6, 9), и отражает напряженность всех функциональных систем при переходе от пренатальной к постнатальной стадии и воздействии факторов окружающей среды (22), что определяет гомеостатические параметры, обеспечивавшие образование de novo пластических и энергетических ресурсов для последующего роста и развития (3, 9, 23) (см. табл. 2, рис.). На 7-е сут у бройлерной птицы установлен фактор (см. табл. 2, рис.), включающий прогестерон и кортизол как главные компоненты (r-Pearson: P4 и Cortisol — r = 0,73, p = 0,016), и фактор донорства холестерола посредством активной конверсии холестерина LDL на синтез прогестерона и соответственно эндокринной цепи 17-OHP с его продуктами — половыми гормонами, кортизолом и другими стероидами (15, 16) (см. табл. 1, 2, рис.). Прогестерон (наряду с соматотропном и тиреоидными гормонами) обеспечивает прирост массы тела, прежде всего за счет регуляции синтеза белков, в основном протеинов скелетной мускулатуры и тканей внутренних органов (24). Прогестерон оказывает влияние на основной обмен у кур, повышает интенсивность метаболизма. Кортизол непосредственно участвует в комплексных приспособительных реакциях как на молекулярном, мембранном, клеточном, тканевом, органном уровнях (10, 16), так и системно, усиливает или ослабляет (в зависимости от цены адаптации в соответствии с периодом онтогенеза) адаптивные реакции, взаимно регулируя метаболизм 17-OHP (25) (см. рис.). Известно что эстрогены и другие гормональные продукты 17-гидроксипрогестерона в норме способствуют утилизации LDL и усилению синтеза HDL, вовлечены в обеспечение пластическими элементами (24, 26-28) (см. рис.).
В возрасте 23 сут у цыплят выявили два фактора — один включал HDL и 17-OHP, другой — прогестерон и кортизол (см. табл. 2), что согласуется с данными S. Rettenbacher с соавт. (29). Они показали, что повышение содержания глюкокортикоидов вследствие стресса или экзогенного экспериментального воздействия (например, введения интактным курам) может приводить к снижению превращения прогестерона в 17-OHP и его производные из-за ингибирования специфических ферментов (в том числе 17-α-гидроксилазы, 17,20-лиазы, 17p-гидроксистероиддегидрогеназы) или конкурентного взаимодействия глюкокортикоидов с ядерными и мембранными рецепторами прогестерона, участвующими в регуляции образования синтезирующих его ферментов (29). В то же время в условиях in vitro без экзогенного стрессирования глюкокортикоиды не препятствуют превращению прогестерона в инкубируемых тканях яичника курочек и тестикул петушков в 17-OHP и далее в андростендион, тестостерон и другие стероидные продукты (29). Показано (30), что стрессы ингибируют фермент, ответственный за конверсию холестерола в прегненолон. При этом, как отмечалось и нами (см. табл. 1), прогестерон, как и кортизол, имел стабильную динамику со статистически недостоверным различием содержания с 7-х по 42-е сут, а пики по 17-OHP и приросту массы, наоборот, достоверно пришлись на период с 7-х по 23-и сут (см. табл. 1). Достижение баланса интенсивных приспособительных и ростовых реакций основывается на высоких ресурсных затратах и в наибольшей степени может проявляться в начале формирования функциональных систем (3, 22, 31). Так, у цыплят яичного кросса отмечен наибольший рост массы тела во II и III декадах (8), что согласуется с нашими данными (см. табл. 1). Выявленное в возрасте 42 сут донорство холестерина (см. табл. 2) (r-Pearson: LDL и TCS — r = 0,86, p = 0,002), видимо, обеспечивает эндокринный пул синтеза прогестерона и его производных (15, 16, 25) (см. табл. 1, 2, рис.). Сообщалось (8), что у цыплят кросса белый леггорн рост стабилизируется после 30-х сут жизни. В нашем исследовании стабилизацию и консолидацию приспособительных и ростовых процессов в IV и начале V декады отражали главные компоненты, выявленные в возрасте 42 сут (см. табл. 2).
Эти сложные взаимодействия обеспечиваются через рецепторы гормонов оси прогестерона (14, 28), в том числе глюкокортикоидов. Происходит модуляция метаболизма липопротеинов и реализация их физиологических эффектов (28) (см. рис.), в частности сосудистых. В события вовлекаются внутриклеточные ядерные рецепторы (14) и так называемых внеге-номные интегральные мембранные рецепторы прогестерона (14, 15), участвующие в экспресс-ответах, не сопровождающихся генной транскрипцией (14, 15), и, видимо, необходимые в процессах роста и развития.
Таким образом, в совокупности полученные данные свидетельствуют, что при росте и развитии цыплят изменяется скорость использования холестерина для биосинтеза стероидных гормонов, что отражается на концентрации липопротеинов и гормонов в крови, а также их взаимосвязи. Возраст цыплят и, соответственно, характер физиологических процессов в их организме, в том числе опосредуемых прогестероном, 17-гид-роксипрогестероном и кортизолом, определяет формирование гомеостаза, обеспечивающего развитие устойчивости к физиологическим последствиям промышленных стрессов. Продолжением исследований может быть оценка роли этих гормонов и метаболитов в определении пределов реакции, приспособительных возможностей бройлеров, цены адаптации к факторам интенсивного воспроизводства в связи с границами нормы и патологии.
Список литературы Об участии холестерола, прогестерона, кортизола и липопротеинов в возрастных изменениях обмена веществ у цыплят-бройлеров промышленного кросса
- Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. Избранные труды. М., 1982.
- Кулаев Б.С. Эволюция гомеостазиса в биологическом пространстве-времени/Под ред. Л.М. Чайлахяна. М., 2006.
- Колесник Е.А., Дерхо М.А. Комплексная оценка роли гормональных и метаболических факторов в процессах роста и развития у цыплят-бройлеров. Проблемы биологии продуктивных животных, 2015, 4: 72-81.
- Хочачка П., Сомеро Дж. Биохимическая адаптация. М., 1988.
- Тертерян Е.Е. Особенности постнатального обмена липидов у кур в условиях континентального климата Армении. Мат. IV съезда Армянского физиологического общества им. Л.А. Орбели. Ереван, 1987: 320.
- Фисинин В.И., Сурай П. Первые дни жизни цыплят: от защиты от стрессов к эффективной адаптации. Птицеводство, 2012, 2: 11-15.
- Peng Y.Z., Wang Y.W., Ning D., Guo Y.M. Changes of haematic parameters, redox status and mitochondrial complex activity in the heart and liver of broilers fed with different density diets under low ambient temperature. Avian Pathology, 2013, 42(4): 327-334 ( ) DOI: 10.1080/03079457.2013.800941
- Тертерян Е.Е., Григорян М.С, Абрамян Г.Э., Манукян С.С. Физиологические особенности постнатального формирования естественной резистентности кур, взаимосвязь с липидным обменом и продуктивностью. Тез. XV съезда Всесоюзного физиологического общества им. И.П. Павлова (Кишинев, 1987). Ленинград, 1987, т. 2: 595.
- Колесник Е.А., Дерхо М.А. Оценка интенсивности обмена веществ и прироста массы тела у цыплят-бройлеров по липопротеиновому индексу. Ветеринария, 2014, 7: 47-51.
- Wan Y., Ma C., Wei P., Fang Q., Guo X., Zhou B., Jiang R. Dynamic expression of HSP90B1 mRNA in the hypothalamus of two Chinese chicken breeds under heat stress and association analysis with a SNP in Huainan chickens. Czech. J. Anim. Sci., 2017, 62(2): 82-87 ( ) DOI: 10.17221/8/2016-CJAS
- Gröschl M., Rauh M., Dörr H.G. Circadian rhythm of salivary cortisol, 17-OH-progesterone, and progesterone in healthy children. Clinical Chemistry, 2003, 49(10): 1688-1691 ( ) DOI: 10.1373/49.10.1688
- Tsutsui K. Neurosteroid biosynthesis and function in the brain of domestic birds. Frontiers in Endocrinology, 2011, 2(37): 1-14 ( ) DOI: 10.3389/fendo.2011.00037
- Pamir E., Ali D., Ismail C., Sanli E., Ali K., Mustafa T. The effects of high dose proges-terone on neural tube development in early chick embryos. Neurology India, 2006, 54(2): 178-181.
- Smith J.L., Kupchak B.R., Garitaonandia I., Hoang L.K., Maina A.S., Regalla L.M., Lyons T.J. Heterologous expression of human mPRalpha, mPRbeta and mPRgamma in yeast confirms their ability to function as membrane progesterone receptors. Steroids, 2008, 73: 1160-1173 ( ) DOI: 10.1016/j.steroids.2008.05.003
- Voican A., Francou B., Novac L., Chabbert-Buffet N., Canonico M., Meduri G., Lombes M., Scarabin P.-Y., Young J., Guiochon-Mantel A., Bouligand J. Pharmacology of hormone replacement therapy in menopause. In: Pharmacology/L. Gallelli (ed.). INTECH, 2012: 313-338 ( ) DOI: 10.5772/32655
- Hough D., Swart P., Cloete S. Exploration of the hypothalamic-pituitary-adrenal axis to improve animal welfare by means of genetic selection: lessons from the South African merino. Animals, 2013, 3: 442-474 ( ) DOI: 10.3390/ani3020442
- Рекомендации по кормлению сельскохозяйственной птицы/Под ред. В.И. Фисинина, Ш.А. Имангулова, И.А. Егорова, Т.М. Околеловой. Сергиев Посад, 2004.
- Титов В.Н., Востров И.А., Каба С.И., Амелюшкина В.А., Ширяева Ю.К. Липопротеины низкой и очень низкой плотности: патогенетическое и клиническое значение. Клиническая медицина, 2013, 1: 20-27.
- Брайнек P., Трубникова Т.Ю. Прямые методы определения холестерина липопротеидов высокой и низкой плотности. Принципы и аналитические характерис-тики. Erba Lachema s.r.o., Brno, 2015.
- De Geyter C., De Geyter M., Huber P.R., Nieschlag E., Holzgreve W. Progesterone serum levels during the follicular phase of the menstrual cycle originate from the crosstalk between the ovaries and the adrenal cortex. Hum. Reprod., 2002, 17(4): 933-939.
- Сычев С.Н. Применение метода главных компонент (факторного анализа) для анализа хроматографических данных в ВЭЖХ. Сорбционные и хроматографические процессы, 2004, 4(2): 134-143.
- Cogburn L.A., Porter T.E., Duclos M.J., Simon J., Burgess S.C., Zhu J.J., Cheng H.H., Dodgson J.B., Burnside J. Functional genomics of the chicken -a model organism. Poultry Sci., 2007, 86(10): 2059-2094 ( ) DOI: 10.1093/ps/86.10.2059
- Surai P.F. Antioxidant systems in poultry biology: heat shock proteins. Journal of Science, 2015, 5(12): 1188-1222.
- Brehme U., Bruck B., Gugel N., Wehrmann M., Hanke S., Finking G., Schmahl F.W., Hanke H. Aortic plaque size and endometrial response in cholesterol-fed rabbits treated with estrogen plus continuous or sequential progestin. Arteriosclerosis, Thrombosis, and Vascular Biology, 1999, 19: 1930-1937 ( ) DOI: 10.1161/01.ATV.19.8.1930
- Rettenbacher S., Möstl E., Groothuis T.G.G. Gestagens and glucocorticoids in chicken eggs. General and Comparative Endocrinology, 2009, 164: 125-129 ( ) DOI: 10.1016/j.ygcen.2009.05.019
- Schwenke D.C. Gender differences in intima-media permeability to low-density lipoprotein at atherosclerosis-prone aortic sites in rabbits. Lack of effect of 17 beta-estradiol. Arteriosclerosis, Thrombosis, and Vascular Biology, 1997, 17(10): 2150-2157.
- Hanke H., Hanke S., Finking G., Muhic-Lohrer A., Mück A.O., Schmahl F.W., Haasis R., Hombach V. Different effects of estrogen and progesterone on experimental atherosclerosis in female versus male rabbits. Quantification of cellular proliferation by bromodeoxyuridine. Circulation, 1996, 94(2): 175-181 ( ) DOI: 10.1161/01.CIR.94.2.175
- McCrohon J.A., Nakhla S., Jessup W., Stanley K.K., Celermajer D.S. Estrogen and progesterone reduce lipid accumulation in human monocyte-derived macrophages a sex-specific effect. Circulation, 1999, 100: 2319-2325 ( ) DOI: 10.1161/01.CIR.100.23.2319
- Rettenbacher S., Henriksen R., Groothuids T.G., Lepschy M. Corticosterone metabolism by chicken follicle cells does not affect ovarian reproductive hormone synthesis in vitro. General and Comparative Endocrinology, 2013, 184: 67-74 ( ) DOI: 10.1016/j.ygcen.2012.12.013
- Stojkov N.J., Janjic M.M., Bjelic M.M., Mihajlovic A.I., Kostic T.S., Andric S.A. Repeated immobilization stress disturbed steroidogenic machinery and stimulated the expression of cAMP signaling elements and adrenergic receptors in Leydig cells. Am. J. Physiol. Endocrinol. Metab., 2012, 302: E1239-E1251 ( ) DOI: 10.1152/ajpendo.00554.2011
- Колесник Е.А., Дерхо М.А. О кластерной системе фосфолипидов в онтогенезе бройлерных цыплят. Сельскохозяйственная биология, 2015, 50(2): 217-224 ( , 10.15389/agrobiology.2015.2.217eng) DOI: 10.15389/agrobiology.2015.2.217rus
