Образование аномальных цветков в потомстве удвоенных гаплоидов кабачка (Cucurbita pepo L.)
")
Автор: Домблидес Е.А., Шмыкова Н.А., Химич Г.А., Коротцева И.Б., Домблидес А.С.
Журнал: Овощи России @vegetables
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 5 (43), 2018 года.
Бесплатный доступ
Культуру неопыленных семяпочек кабачка in vitro использовали для создания полностью гомозиготных линий с целью ускорения селекционного процесса. В результате проведенной оценки по хозяйственно ценным признакам было выделено семь перспективных DH-линий, полученных из разных селекционных образцов кабачка, отличающихся по форме и окраске плода. Все полученные линии отличались высокой однородностью, сохраняющейся при последующем размножении, и были наделены комплексом хозяйственно ценных признаков. DH-линии относились к женскому типу и имели до 96% женских цветков и лишь 4% мужских, причем мужские цветки появлялись через две недели после начала цветения женскими цветками, что представляет большую селекционную ценность. На отобранных DH-линиях было отмечено формирование морфологически аномальных женских и мужских цветков, а также пинандроморфных цветков. За период вегетации на одном растении образовывалось от 26 до 36 цветков, при этом 19-21 штук из них были нормально развитыми женскими цветками, 3-5 штук - нормальными мужскими цветками и до 10-11штук цветков развивались с той или иной аномалией...
Кабачок (cucurbita pepo l.), dh-растения, культура неопыленных семяпочек in vitro, гиногенез, гаплоиды кабачка, аномальные цветки, гинандроморфные цветки, обоеполые цветки
Короткий адрес: https://sciup.org/140238362
IDR: 140238362 | УДК: 635.621.3:575.224.234 | DOI: 10.18619/2072-9146-2018-5-13-17
Development of abnormal flowers in generation of doubled haploids of summer squash (Cucurbita pepo L.)
The culture of unpollinated ovules in vitro in summer squash was used to develop fully homozygous breeding lines with the aim of the speeding-up breeding program. As a resuit of assessment for economically valuable traits, the seven promising DH-lines obtained from summer squash accessions differed by fruit shapes and colours were selected out. All breeding lines produced showed high homogeneity that retained in following generations and also have an appropriate set of economically valuable traits. DH-lines belonging to female type have up to 96% female flowers and only 4% male flowers. It is very important for breeding when the male flowers appeared in two weeks just after the female flower began blooming. The development of morphologically abnormal female and male flowers, along with gynandromorphy flowers was noted on selected DH-lines. During vegetation period from 26 to 36 flowers appeared on the plant, where out of them 19-21 ones were normally developed female flowers, 3-5 ones were normally developed male flowers, and up to 10-11 ones showed an abnormal way of development...
Текст научной статьи Образование аномальных цветков в потомстве удвоенных гаплоидов кабачка (Cucurbita pepo L.)
Семейство Cucurbitaceae является вторым после Solanaceae большим семейством овощных культур по экономической важности в мире. Оно включает в себя такие важные культуры, как дыня (Cucumis melo), арбуз (Citrullus lanatus), огурец (Cucumis sativus) и многие виды Cucurbita со съедобными плодами (Jeffrey, 1980). Род Cucurbita (2x = 2n = 40), возникший в Северной и Южной Америке, включает в себя три экономически важных вида, таких как Cucurbita pepo, Cucurbita moschata и Cucurbita maxima, выращиваемых в умеренных, субтропических и тропических регионах (Wang et al., 2011). Вид C. pepo представлен широким ассортиментом сортов и гибридов, известных своей уникальной формой и цветом плодов, и высоко ценится за свои кулинарные свойства. Среди различных видов этого рода C. pepo, куда относится и кабачок, имеет наибольшую экономическую ценность. Род Cucurbita представлен в основном однодомными формами, однако отдельные виды могут иметь гермафродитные, полигамные или двудомные формы. В связи с таким большим разнообразием этот род представляет большой интерес с точки зрения изучения генов, регулирующих детерминацию пола у тыквенных культур. Обнаружение аномалий в развитии цветка, закрепленных генетически, представляет ценность, поскольку эти растения в дальнейшем смогут служить генетическими моделями, а использование для этих целей DH-линий особенно эффективно, поскольку они содержат все гены в гомозиготном состоянии. В литературе имеются лишь единичные сообщения об аномальных цветках у кабачка, обнаруженных спонтанно в природной популяции (Кордюм, Глущенко, 1976), полученных искусственно с использованием мутагенов (Manzano et al., 2012) и среди потомства удвоенных гаплоидов, образовавшихся в культуре пыльников (Metwally et al., 1998).
В нашей работе мы использовали культуру неопыленных семяпочек кабачка in vitro для создания полностью гомозиготных линий с целью ускорения селекционного процесса. Среди потомства удвоенных гаплоидов (DH-растения) были обнаружены представляющие большой селекционный интерес DH-линии, образующие аномальные цветки, до этого не встречающиеся в нашей селекционной практике. Описанию и изучению новых форм аномальных цветков кабачка посвящена данная работа.
Материалы и методы Растительный материал и условия выращивания донорных растений
В работе использовали селекционные образцы кабачка из коллекции лаборатории тыквенных культур ФГБНУ ФНЦО. Донорные растения выращивали в 2014-2017 годах в Московской области в условиях пленочной теплицы и открытого грунта.
Получение DH-растений кабачка в культуре неопыленых семяпочек in vitro
Для индукции гиногенеза использовали питательную среду IМС (Induction Medium for Cucurbitaceae ), разработанную в лаборатории биотехнологии ФГБНУ ФНЦО (ВНИИССОК).
Культивирование проводили на стеллажах с люминесцентными лампами при 25°С и фотопериоде 14 часов, освещенности 2,5 тыс. люкс.
Получение растений-регенерантов
Нормально развитые образовавшиеся эмбриоиды переносили на без-гормональную среду MС (Murashige and Skoog, 1962) с 2% сахарозой и 3 г/л фитогеля и культивировали на стеллажах при тех же условиях.
Выращивание растений-регенерантов
Растения с нормально развитыми листьями и корневой системой переносили в вегетационные сосуды, заполненные смесью минерализованного торфа и перлита (7:3), накрывали перфорированными пластиковыми стаканчиками для адаптации растений к условиям in vivo. Выращивали растения-регенеранты в климатической камере при режиме 25°С круглосуточно, 16 ч. день/8 ч. ночь
Результаты
В результате проведенной работы из 9 отозвавшихся в культуре неопы-ленных семяпочек образцов C. pepo в течение 4-х лет (2014-2017) было получено 341 растение-регенерант. На этапе адаптации к условиям in vivo потери составили 25%. Около 7% растений были предположительно гаплоидными, и получить от них потомство не удалось. Все адаптированные DH-растения были переданы селекционерам для оценки и дальнейшей селекционной работы. Для включения удвоенных гаплоидов в селекционный процесс необходимо получить потомство от этих растений. У тыквенных культур на этом этапе работы встречаются трудности. Семейство Cucurbitaceae представлено в основном однодомными формами, то есть растения с раздельнополыми – мужскими (тычиночными) и женскими (пестичными) цветками, формируются на одной и той же особи. Для самоопыления индивидуального растения необходимо одновременное цветение мужскими и женскими цветками, что происходит достаточно редко (рис.1). Поэтому не всегда удается получить потомство от самоопыления. Мы можем рекомендовать использовать на этапе регенерации клональное микроразмножение, чтоб увеличить количество растений с одинаковым генотипом, и в этом случае их можно высаживать совместно и самоопылять внутри группы.

Рис. 1. Одновременное раскрытие мужского и женского цветка на растении-регенеранте R 0 .
Fig. 1. Regenerated plant R0 with simultaneous male and female flower opening.
В результате проведенной оценки по хозяйственно ценным признакам, было выделено 7 перспективных DH-линий, включенных в селекционный процесс (рис. 2). Все полученные линии отличались высокой однородностью, сохраняющейся при последующем размножении, и были наделены комплексом хозяйственно-ценных признаков. Особенно ценно, что выделенные линии, помимо высокой продуктивности, характеризовались очень большой насыщенностью женскими цветками, что нетипично для вида C. pepo. Сорта и гибриды кабачка, у которых преобладают мужские цветки, относят к мужскому типу цветения, имеющие – 50% мужских и 50% женских цветков – к смешанному типу, а с преобладанием женских цветков – к женскому типу. Все выделенные нами DH-линии относились к женскому типу и имели до 96% женских цветков и лишь 4% мужских, причем мужские цветки появлялись через две недели после начала цветения женскими цветками, что представляет большую селекционную ценность. Однако на растениях этих линий одновременно с нормально развивающимися цветками формировались и аномальные цветки, и плоды (рис. 3), что значительно снижало их семенную продуктивность.

Рис. 2. Отобранные селекционно-ценные DH-линии кабачка.
Fig. 2. Selected DH valuable breeding lines of summer squash.

Рис.3. Нормальный и аномальный цветок на растении кабачка.
Fig. 3. Normal and abnormal flowers on plant of summer squash.
В норме цветки C. pepo крупные, жёлтые или белые правильные, одиночные или собраны в пучки (мужские цветки); цветки однополые (растения однодомные), но в редких случаях способны формировать обоеполые цветки. Чашечка и венчик колокольчатые или ворончато-колокольчатые из пяти (редко четырёх-семи) долей. В мужских цветках тычинки спаялись пыльниками в головку, пыльники извитые; в женском цветке развиты три-пять стаминодиев и пестик, с толстым коротким столбиком, с трёх- или пятилопастным рыльцем и с нижней, 3-5-гнёздной многосемянной завязью. Плод – тыквина (лат. pepo, peponium ) – паракарпный многосемянный плод с сочным эндокарпием, мясистым мезокарпием и твердым экзокарпием. Плод образуется из нижней завязи, включающей обычно 3 плодолистика (рис.4, 5).
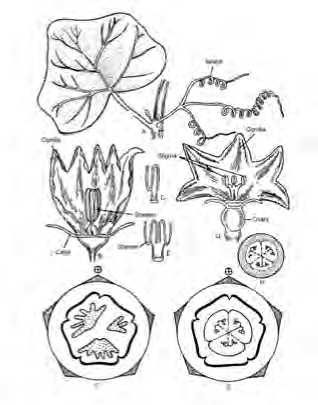
Рис. 4. Строение типичного мужского и женского цветка растений рода Cucurbita (по Sambamury, 2005).
-
А. междоузлие;
-
В. мужской цветок;
-
С. однолопастная тычинка;
-
D. женский цветок;
-
Е. дввухлопастная тычинка;
-
F. цветочная диаграмма мужского цветка;
-
G. цветочная диаграмма женского цветка;
-
H. поперечный срез завязи.
Fig. 4. Typical male and female flowers in genus Cucurbita (by Sambamury, 2005)
-
A. a twig;
-
B. a male flower;
-
C. Single lobed stamen;
-
D. a female flower;
-
E. doubled lobed stamen;
-
F. floral diagram of male flower;
-
G. floral diagram of female flower;
-
H. an ovary

Г
Д
Е
Рис. 5. Строение мужского и женского цветка кабачка. Внешний вид мужского (А) и женского (Г) цветка кабачка; вид мужского (Б) и женского (Д) цветка с удаленным околоцветником; сросшиеся в столбик тычинки и их поперечный срез (В); продольный срез женского цветка (Е).
Fig. 5. Male and female flowers of summer squash. Male (A) and female flower (Г); male (Б) and female (Д) with removed floral envelope; stamens accreted together into a style and its transverse section; longitudinal section of a female flower (E)
На растениях выделенных 7 линий, полученных из разных селекционных образцов кабачка, различающихся по форме и окраске плода, было отмечено формирование морфологически аномальных женских и мужских цветков, а также гинандроморфных цветков. За период вегетации на одном растении образовывалось от 26 до 36 цветков, при этом 19-21 штук из них были нормально развитыми женскими цветками, 3-5 штук – нормальными мужскими цветками и до 10-11 штук цветков развивались с той или иной аномалией. Процент уродливых цветков на этих линиях оставался неизменным, как при выращивании этих линий в пленочной теплице, где температура воздуха и влажность были достаточно высокими, так и в условиях открытого грунта. При самоопылении этих линий образование уродливых аномальных цветков наследовалось и проявлялось у потомства. Было отмечено, что при скрещивании DH-линий кабачка, образующих аномальные цветки с другими линиями или сортами, в потомстве образование аномальных цветков не наблюдалось. Возможно этот признак контролируется рецесивными генами, и требуется проведение дальнейших генетических исследований для выяснения природы его наследования.
Чаще всего мы отмечали образование на растении аномальных женских цветков, они составляли до 85% от общего числа всех обнаруженных аномалий. Эти цветки можно было определить по внешнему виду еще до раскрытия цветка. Степень развития аномалии женского цветка могла варьировать (рис.6). Рыльце могло либо вообще отсутствовать, либо быть представленным одно- или двухлопастной формой. Вместо обычного трехлопастного рыльца на верхушке завязи формировались уродливые образования из дополнительных плодолистиков. При этом часть плодолистиков могла быть полностью стерильной и обнаруживать сходство с рыльцем, у другой части могло образовываться незначительное число семяпочек на брюшной стороне плодолистиков. В других случаях семяпочки вообще могли лежать открыто, при этом они значительно разрастались и по внешнему виду были сходны с зачатками семян. Однако, как и в случае аномалий, описанных Кордюм и Глущенко в 1976 году, они не содержали зародыша и эндосперма, а увеличение размера происходило за счет разрастания паренхимной ткани интегументов и нуцеллуса (Кордюм, Глущенко, 1976). В цветках, у которых рыльце хотя бы частично формировалось, опыление было возможно, хотя семена образовывались только в количестве 4-5 штук. Плоды из таких цветков формировались и могли достигать 30 см в длину (фаза технической спелости). Бутоны женских цветков с полностью отсутствующим рыльцем обычно не распускались, увеличивались в размере, достигая фазы технической спелости, однако через месяц верхушка плода начинала загнивать.
Кроме аномальных женских цветков, на


Рис. 7. Внешний вид стерильных мужских цветков.
Fig. 7. Sterile male flowers.

Рис. 6. Внешний вид аномальных женских цветков кабачка.
Fig. 6. Abnormal female flower of summer squash.
линиях удвоенных гаплоидов кабачка могли образовываться и аномальные стерильные мужские цветки (рис. 7). Эти цветки обычно были мельче по размеру и не образовывали фертильной пыльцы.
Кроме аномальных женских и мужских цветков, были обнаружены и обоеполые цветки. Формирование обоеполых цветков у растений в норме с однополыми цветками следует рассматривать как явление гинанд-роморфизма. На ранних этапах органогенеза культур семейства Cucurbitaceae отсутствуют четкие морфологические отличия между цветками различных половых типов (мужскими, женскими, обоеполыми) у разных форм (Whitaker, 1931, Пангало, 1943). В мужских и женских цветках большинства тыквенных элементы органов другого пола останавливаются в своем развитии, как правило, на стадии недифференцированных зачатков и обнаруживаются в сформированных цветках лишь в виде небольших бугорков (Кордюм, Глущенко, 1976). В наших исследованиях гинандроморфные цветки мы наблюдали на DH-линиях, причем степень развития гинецея и андроцея варьировала, наблюдались различные отклонения, вплоть до образования полностью стерильных цветков (рис. 8). В литературе нам встречались единичные сообщения об образовании сходных гинанд-роморфных цветков у кабачка и огурца (Кордюм, Глущенко, 1976). В наших исследованиях чаще всего тычинки гинандроморфно-го цветка имели нити нормальной длины, а

Рис. 8. Образование аномальных обоеполых (гинандроморфных) цветков.
Fig. 8. Development of abnormal hermaphrodite and bisexual (gynandromorph) flowers.
пыльники были заполнены пыльцевыми зернами. Верхушки тычиночных нитей и пыльники тесно примыкали к аномально развитому гинецею. Иногда срастание пыльников было неполное (рис.8). Бокал завязи у таких цветков чаще всего был не сформирован, и завязь производила впечатление верхней. Это могло свидетельствовать о том, что описанные гинандроморфные цветки формировались из мужских. Рыльца, как и в случае, описанном Кордюм и Глущенко в 1976 году на обнаруженном аномальном цветке кабачка сорта Грибовский 37, чаще всего имели вид уродливых образований. При самоопылении таких цветков нам иногда удавалось получить семена, однако их количество было не более четырех в одном плоде.
Обсуждения
В результате проведенной работы нам удалось описать встречающиеся аномалии в развитии мужских и женских цветков кабачка C. pepo L., обнаруженные в DH-линиях, полученных через культуру неопыленных семяпочек in vitro . Упоминание об образовании похожих гинандроморфных уродливых форм было сделано в работе Кордюм и Глущенко в 1976 году, где они описали цветки с единичного обнаруженного в сорте Грибовский растения кабачка (Кордюм, Глущенко, 1976). Аномальные цветки на растениях-регенерантах кабачка были обнаружены среди потомства, полученного через культуру пыльников (Metwally et al. (1998), однако авторы не дали подробного описания встретившихся аномалий. Также фото и описание трех мутантных линий, полученных искусственно при использовании мутагена этил метан сульфоната (ethyl methane sulfonate (EMS)) было сделано в работе Manzano с соавторами в 2012 году (Manzano et al., 2012). На этих мутантных линиях (внешне они были похожи на некоторые обнаруженныe нами гинандроморфные цветки) авторы в дальнейшем изучали гены, контролирующие чувствительность к этилену и переход женских цветков к бисексуальным и гермофродитным (Garcнa et al., 2016). Описание образования уродливых аномальных цветков среди потомства DH-линий, полученных в культуре неопыленных семяпочек в литературе нам не встречалось.
Полученные нами в культуре неопыленных семяпочек DH-линии с аномальными цветками могут служить прекрасным материалом не только для селекции, но и в исследованиях по изучению детерминации пола у тыквенных культур. Поскольку образование уродливых форм отмечалось и в последующих от самоопыления потомствах, то эти линии представляют интерес для генетических, эмбриологических и эволюционных исследований.
Список литературы Образование аномальных цветков в потомстве удвоенных гаплоидов кабачка (Cucurbita pepo L.)
- Кордюм Е.Л., Глущенко Г.И. Цитоэмбриологические аспекты проблемы пола покрытосеменных. «Наукова думка», Киев, 1976. -200 с.
- Пангало К.И. Пол и цветение у возделываемых Cucurbitaceae. Ботанический журнал СССР, 1943. -28, 1. -С.10-23.
- Garcнa А, Manzano S, Aguado E, Megнas Z, Martнnez C, Garrido D, Jamilena M. Isolation and Characterization of Three Recessive Andromonoecious Mutants of Cucurbita pepo. Proceedings of Cucurbitaceae 2016, the XIth EUCARPIA Meeting on Genetics and Breeding of Cucurbitaceae. July 24-28, 2016, Warsaw, Poland, p. 45-49.
- Jeffrey C. (1980). A review of the Cucurbitaceae. Botanical Journal of the Linnean Society, 81(3), 233-247 DOI: 10.1111/j.1095-8339.1980.tb01676
- Manzano S, Megнas Z, Marinez C, Jamilena M (2012) Generation and evaluation of a preliminary EMS mutant library in zucchini squash. In: Sari N, Solmaz I, Aras V (eds) Cucurbitaceae 2012. Adana: Cukurova University, pp 669-673.
- Metwally EI, Moustafa S.A., El-Sawy B.I. & Shalaby T.A. (1998) Haploid plantlets derived by anther culture of Cucurbita pepo. Plant Cell, Tissue and Organ Culture 52: 171-176, 1998.
- Murashige T., Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum, 1962, 15: 473-497 ( ) DOI: 10.1111/j.1399-3054.1962.tb08052.x
- Whitaker, T.W. (1931) Sex ratio and sex expression in the cultivated Cucurbits. -Amer. J. Bot., 1931, 18, p. 359.
- Wang Y.H., Behera T.K., Kole C., editors. (eds.). (2011). Genetics, Genomics and Breeding of Cucurbits. CRC Press.
