Обзор биотехнологических подходов к биообогащению Triticum aestivum L.

Автор: Серазетдинова Ю.Р., Асякина Л.К.
Рубрика: Актуальные проблемы развития пищевых и биотехнологий
Статья в выпуске: 2 т.13, 2025 года.
Бесплатный доступ
«Скрытый голод» (длительный дефицит микронутриентов в питании человека) - глобальная проблема, затрагивающая треть населения планеты. Оптимизация питания населения выступает приоритетной задачей многих стран мира. Для решения этой задачи может применяться эффективная стратегия - биофортификация, которая представляет собой процесс повышения содержания биодоступных микроэлементов в сельскохозяйственных культурах до сбора урожая. Целью исследования является анализ современных достижений в области повышения биологической ценности зерна пшеницы. Анализ современных новых источников показал, что существует несколько походов к биофортификации. К ним относят генетическую трансформацию, а также агрономические методы. Каждый из методов обладает рядом преимуществ и недостатков. Генетические подходы являются эффективными и экономичными, однако сталкиваются с недоверием общественности. Агрономическая биофортификация активно применяется в сельскохозяйственной практике и позволяет не только обогатить культуры необходимыми микро- и макроэлементами, но также повысить урожайность культур. Однако этот способ сопряжен с большими экономическими затратами, а также негативным влиянием на состояние окружающей среды. Наиболее перспективным способом является биообогащение пшеницы с помощью микроорганизмов. Они способны повышать доступность биогенных элементов для растений благодаря выделению различных метаболитов, включая органические кислоты, фитогормоны и сидерофоры. Кроме того, они способны влиять на транспортные системы растения, усиливая поглощение определенных элементов. Микробная биофортификация представляет собой экологически безопасный и экономически эффективный метод обогащения пшеницы микроэлементами для борьбы со «скрытым голодом».
Биофортификация, пшеница, скрытый голод, генетическая трансформация, селекция, агрономический метод, микроорганизмы, пищевая ценность
Короткий адрес: https://sciup.org/147250719
IDR: 147250719 | УДК: 633.11: | DOI: 10.14529/food250201
Review of biotechnological approaches to bio-enrichment of Triticum aestivum L.
“Hidden hunger” (prolonged micronutrient deficiency in human nutrition) is a global problem affecting a third of the world's population. Optimizing the nutrition of the population is a priority task in many countries around the world. To solve this problem, an effective strategy can be applied - biofortification, which is the process of increasing the content of bioavailable trace elements in agricultural crops before harvest. The purpose of the study is to analyze modern achievements in the field of increasing the biological value of wheat grain. An analysis of modern new sources has shown that there are several trips to biofortification. These include genetic transformation, as well as agronomic methods. Each method has a number of advantages and disadvantages. Genetic approaches are effective and cost-effective, but they face public distrust. Agronomic biofortification is actively used in agricultural practice and allows not only to enrich crops with essential micro- and macronutrients, but also to increase crop yields. However, this method is associated with high economic costs, as well as a negative impact on the environment. The most promising method is bio-enrichment of wheat with the help of microorganisms. They are able to increase the availability of biogenic elements for plants due to the release of various metabolites including organic acids, phytohormones and siderophores. In addition, they are able to influence the plant's transport systems by enhancing the absorption of certain elements. Microbial biofortification is an ecologically safe and economically effective method of enriching wheat with trace elements to combat “hidden hunger”.
Текст научной статьи Обзор биотехнологических подходов к биообогащению Triticum aestivum L.
Недостаток микро- и макронутриентов в питании, называемый «скрытый голод», представляет собой глобальную проблему общественного здравоохранения, затрагивающую около 33 % населения Земли [1]. Вследствие этого оптимизация питания населения является приоритетной задачей для многих стран. В последние годы наблюдается стремительный рост алиментарно зависимых заболеваний, ассоциированных со «скрытым голодом». Беременные женщины и дети до пяти лет представляют собой группы повышенного риска развития тяжелых последствий, связанных с дефицитом микронутриентов. Согласно современным данным, около 45 % случаев детской смертности в возрасте до пяти лет в мире ассоциировано с нарушениями питания [2]. В связи с этим одним из ведущих трендов пищевой промышленности является разработка биообогащенных продуктов, процесс получения которых называют биофортификацией.
Биофортификация представляет собой процесс повышения внутренней пищевой ценности сельскохозяйственных культур путем увеличения содержания биодоступных микронутриентов in vivo, то есть до момента сбора урожая. В отличие от традиционного обогащения, предполагающего добавление витаминов и минералов на этапе постуборочной обработки, биофортификация модифицирует метаболические процессы растения. Это достигается за счет усиления поглощения минералов из почвы, стимуляции синтеза вторичных мета- болитов, а также изменения транспорта и метаболизма микроэлементов или соединений, влияющих на их биодоступность [3].
Учитывая, что базовые продукты питания являются основным источником энергии и потребляются в значительных количествах вне зависимости от уровня дохода, их биофортификация представляется наиболее рациональной стратегией борьбы со «скрытым голодом». В связи с этим биобогащение наиболее рационально для продуктов массового употребления. В Российской Федерации к таким продуктам относятся продукты переработки зерновых культур, в частности пшеницы, обеспечивающей до 40 % суточной потребности человека в пище [4]. Несмотря на высокую энергетическую и пищевую ценность, традиционные сорта пшеницы характеризуются низким содержанием эссенциальных микроэлементов (Fe, Zn, Se, Cu) и высоким уровнем антинутриентов, лимитирующих биодоступность [2].
Целью настоящего исследования является анализ современных достижений в области повышения биологической ценности зерна пшеницы.
Объекты и методы исследований
В настоящем исследовании проведен отбор релевантных тематике научных публикаций из авторитетных отечественных и международных реферативных баз данных, включая PubMed, Scopus, ScienceDirect, Elsevier, eLIBRARY.RU и др. Поисковый запрос включал следующие слова и сочетания слов: биофортификация, биобогащение, пшеница, цинк, железо, селен, biofortification, processing and enrichment, wheat, zinc, iron, selenium, Triticum aestivum L. В ходе поиска обнаружено 104 публикации, из которых для дальнейшего анализа отобрано 33.
Обзор литературы ограничен публикациями с открытым доступом к полному тексту. Статьи, не удовлетворяющие этому критерию, а также дубликаты и публикации, не соответствующие тематике исследования (по названию, аннотации или полному тексту), не рассматривались. Критерии отбора представлены в табл. 1.
Таблица 1
Критерии отбора источников
|
Критерий |
Включено |
Исключено |
Причины |
|
Популяция |
Пшеница ( Triticum aestivum L.) |
Другие сельскохозяйственные культуры |
Целью настоящего обзора является обобщение и систематизация знаний о подходах к биообогащению пшеницы |
|
Концепция |
Исследования направленные на изучение накопления биогенных элементов в зерне пшеницы, а также способов интенсификации данного процесса |
Исследования направленные на повышение продуктивности пшеницы, защиты от инфекционных заболеваний |
Данное исследование фокусируется на методах повышения содержания биогенных веществ в пшенице. Другие агротехнические приемы выращивания пшеницы находятся за пределами рассматриваемой области |
|
Язык |
Любой |
Ограничения отсутствуют |
Ввиду глобального распространения ростостимулирующих микроорганизмов, трансформирующих биогенные вещества, и повсеместного выращивания пшеницы, лингвистические ограничения при отборе научных работ не применялись |
|
Временной период |
С 2019 по 2024 гг. |
После декабря 2024 г. |
Для обеспечения актуальности данных, поиск литературы ограничен публикациями последних пяти лет |
|
География |
Любые страны |
Ограничения отсутствуют |
Изучаемая тематика актуальна для всех стран мира |
Библиографические данные извлекали с помощью программного обеспечения Zotero (формат .ris), а также обрабатывали в VOSviewer для анализа ключевых слов. Из 391 извлеченных ключевых слов (34 публикации) отобраны 29, встречающиеся более двух раз. Визуализация взаимосвязи и частоты встречаемости ключевых слов представлена на рис. 1 и 2 соответственно. Размер точки соответствует частоте встречаемости ключевого слова. Семантические связи между ключевыми словами визуализированы линиями различного цвета.
Методологическая база исследования включала структурно-логический, аналитиче- ский и аксиоматический подходы. Обобщение сведений, полученных с помощью данных методов, приведено ниже.
Результаты и обсуждения
Понимание механизмов транспорта и накопления микроэлементов, таких как Zn, Fe, и Se, является необходимым условием для биофортификации любых культур, включая пшеницу. Как правило, процесс поглощения включает извлечение микроэлементов из ризосферы и их радиальный транспорт в ксилему корня. В период налива эти элементы транспортируются в зерно акропетально, то есть по побегу [5].
Поглощение ионов Zn2+ пшеницей осуще- grain se micronutrient
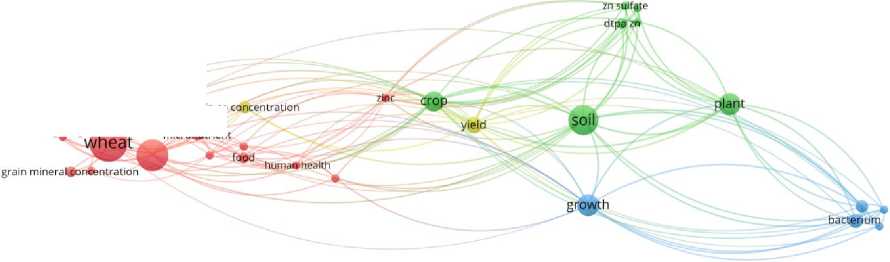
Рис. 1. Карта взаимосвязи ключевых слов

Рис. 2. Частота встречаемости ключевых слов ствляется двумя путями. Первый – активный, селективный и энергозависимый (за счет фотосинтеза) транспорт через мембрану корневых клеток. Второй механизм активируется при дефиците цинка: корни выделяют ком-плексообразователи, например, муравьиную кислоту, которые связывают Zn, увеличивая его концентрацию и подвижность в почвенном растворе. При низкой доступности Zn роль влажности почвы существенно возрастает. Ключевым фактором, определяющим доступность Zn для поглощения, является pH почвы, влияющий на растворимость этого элемента. Низкие значения pH способствуют адсорбции Zn на катионообменных участках почвенных компонентов (например, оксидах металлов), что снижает его доступность в почвенном растворе [6].
Доступность железа определяется pH и окислительно-восстановительным потенциалом почвы. В условиях высокого pH железо легко окисляется, образуя нерастворимые оксиды. Напротив, при низком pH ионы Fe3+ высвобождаются из оксидов, становясь доступным для поглощения корнями. Существуют три стратегии усвоения Fe растениями: восстановление, хелатирование а также комбинация первых двух стратегий. Пшеница для усвоения железа преимущественно использует стратегию хелатирования [7].
Среди злаковых культур пшеница демонстрирует высокую эффективность накопления селена. В настоящее время механизмы поглощения этого элемента растениями до конца не изучены, поскольку он не является для них жизненно необходимым. Однако, учитывая химическое сходство селена и серы, предполагается, что поглощение Se5+ происходит аналогично сульфату, преимущественно через активный транспорт с помощью переносчиков SULTR1;1 и SULTR1;2. При этом установлено, что селен как в форме селенита, так и селената может поглощаться не только через почву, но и с поверхности листа [8].
Накопление металлов, таких как Zn, Fe и Se в зерне является сложным физиологическим признаком, регулируемым кумулятивной экспрессией гена для поглощения, транспортирования, распределения и секвестрации в различных частях растения. Металлы аккумулируются в зерне с помощью трансмембранных транспортеров HMA и CDF, часто называемых MTP и NRAMPS. Микроэлементы откладываются в двух компартментах зер- на: в связанной форме, в виде белка ферритина (накопление Fe) и в вакуоли (Zn и Fe) [9]. Таким образом, повышение уровня или мобильности Zn, Fe и Se в почве, а также оптимизация транспортных систем микроэлементов в растении представляют собой два основных подхода к увеличению их содержания в зерне пшеницы.
Биофортификация может быть достигнута агрономическими методами (внесение удобрений в почву, листовые подкормки) или методами генетической модификации. Различные геномные подходы, такие как картирование локусов количественных признаков (QTL), селекция с помощью маркеров и геномная селекция, широко используются для биофортификации пшеницы [10]. Преимущества и недостатки этого метода биофортификации представлены в табл. 2.
Биофортификация путем селекции осуществляется для тех культур, генетическая изменчивость которых легко доступна из ее первичного, вторичного или третичного генофонда. У пшеницы есть большое количество неиспользуемых диких предков, которые могли бы способствовать генетическому улучшению пшеницы. Геном пшеницы трудно интерпретировать, поскольку он имеет широко распространенные повторяющиеся последовательности, гетерозиготность и полиплоидию [11]. Тем не менее развитие методологий секвенирования, снижение стоимости секвенирования и достижения в вычислительных ресурсах позволили сделать эти ресурсы более доступными [12].
Селекция с помощью маркеров (QTL) является потенциальной стратегией для разработки биообогащенной Fe и Zn пшеницы. Гены, действующие на одном или нескольких этапах, например, поглощение корнями, транслокация из корня в побег, хранение и ремобилизация, могут быть отражены QTL, отвечающими за концентрацию минералов в зерне. Различные исследования по картированию QTL позволили идентифицировать множество QTL как для Fe, так и для Zn. Несколько исследований выявили и картировали QTL для высоких концентраций GFe и GZn на разных хромосомах, 1A, 1B, 1D, 2A, 2B, 3A, 3B, 3D, 4A, 4B, 4D,5A, 5B, 5D, 6A, 6B, 7A, 7B и 7D, обнаруженных в разных диплоидных, тетраплоидных и гексаплоидных видах пшеницы [13, 14]. Например, Gorafi и др. [15] оценили 47 линий синтетической пшеницы,
Таблица 2
Преимущества и недостатки генетической биофортификации Triticum aestivum L.
Понимание генетических механизмов, регулирующих накопление Se в зерне пшеницы, является фундаментальной предпосылкой для разработки стратегий генетической биофортификации этим микроэлементом. В литературе представлены противоречивые данные относительно степени генетической вариабельности содержания Se в зерне среди сортов пшеницы. Некоторые исследования не выявили значимой генетической изменчивости, в то время как другие указывают на наличие генотипов с повышенной аккумуляцией Se [16]. Картирование QTL представляет собой эффективный инструмент для изучения генетической архитектуры данного признака, однако количество исследований, посвященных картированию QTL, ассоциированных с концентрацией Se в зерне пшеницы, остается ограниченным [17, 18] .
Помимо недостаточного объема исследований, генетическая биофортификация пшеницы сопряжена с рядом серьезных трудностей. Так, селекция представляет собой долгосрочный и трудоемкий процесс, требующий большого количества ресурсов [19]. В то время как генная трансформация культур сравнительно быстрее, но сталкивается с общественным неодобрением, а также сложностями государственного регулирования. Например, широко известная генномо-дифицированная культура – «золотой рис», обогащенный провитамином А (β-каротин), разработан более 20 лет назад, но до сих пор не получил разрешение на реализацию в большинстве стран мира [20].
Ввиду указанных ограничений, в настоящее время для получения биофортифициро-ванных сортов пшеницы преимущественно применяются агрономические методы, основанные на внесении удобрений. Преимущества и недостатки этого метода биофортификации представлены в табл. 3.
Данный подход демонстрирует эффективность для повышения содержания ряда микроэлементов в зерне. Например, G.H. Lyons и соавторы установили, что концентрация Se в зерне коррелирует с уровнем внесения Se, независимо от способа применения (в почву или путем листовой подкормки). При этом внесение селена в форме селенита натрия в почву во время посева оказалось более эффективным, чем внекорневая обработка после цветения [21]. Yadav et al. [22] продемонстрировали возможность повышения концентрации Zn в цельном зерне пшеницы до 60 мг/кг путем внекорневой или комбинированной (почвенной и внекорневой) обработки солями Zn. При этом использование наноносителя Zn (наночастицы Zn и хитозана) показало более высокую эффективность по сравнению с сульфатом цинка. Авторы также установили, что аккумуляция Zn в зерне регу-
Таблица 3
Преимущества и недостатки агрономической биофортификации Triticum aestivum L.
В исследовании M.U. Hassan и соавторов оценивалась эффективность различных методов внесения Zn (в почву – 10 кг ZnSO 4 /га, протравливание семян – 0,3 М ZnSO 4 , внекорневая подкормка – 0,5 % ZnSO 4 ) для повышения урожайности и биофортификации пшеницы. Все методы увеличили урожайность (максимально на 6,60 т/га при внесении в почву) и концентрацию Zn в зерне (максимально на 70 % при внекорневой подкормке). Внекорневое внесение показало наибольшую эффективность использования Zn, хотя внесение в почву оказалось наиболее экономически выгодным. Авторы отметили, что для повышения урожайности пшеницы рекомендуется внесение Zn в почву, в то время как для целей биофортификации наиболее эффективна внекорневая подкормка [23].
К недостаткам агрономической биофортификации относят высокие экономические затраты, а также негативное воздействие на окружающую среду, обусловленную низкой биоусвояемостью внесенных микроэлементов. Так, только 20 % внесенного Zn доступно для растений, остальная часть адсорбируется на частицах почвы и переходит в нерастворимую форму) [24]. Продолжительное применение минеральных удобрений негативно сказывается на плодородии почв, изменяя их физикохимические свойства. Накопление основных биогенных элементов (N, P, K) нарушает агрегацию и дисперсию почвенных частиц, способствуя уплотнению почвенного профиля. Избыточное внесение удобрений также ведет к изменениям элементного состава, сдвигу pH и аккумуляции токсичных элементов, вклю- чая тяжелые металлы, что отрицательно влияет на плодородие почв [25, 26].
В связи с этим возрастает интерес исследователей к использованию микроорганизмов для повышения пищевой ценности сельскохозяйственных культур, в частности пшеницы. Преимущества и недостатки этого метода биофортификации представлены в табл. 4.
Ризобактерии, стимулирующие рост растений (PGPR), и некоторые эндофиты обладают способностью фиксировать атмосферный азот, а также способствовать усвоению других микроэлементов. Более того, для многих сельскохозяйственных культур наблюдается опосредованное PGPR стимулирование роста, повышение урожайности и увеличение содержания питательных веществ в зерне [27].
Ключевым механизмом микробной мобилизации питательных веществ в почве является продукция органических кислот и сидерофоров. Снижение pH почвы, обусловленное выделением органических кислот микроорганизмами, способствует растворению таких питательных веществ, как P, K и Zn, за счет разрушения их комплексов с другими металлами или ионами. Сидерофоры, продуцируемые микроорганизмами, повышают биодоступность Fe путем хелатирования с образованием комплекса сидерофор-Fe, играя важную роль в усвоении Fe растениями. Продукция фитогормонов, таких как ИУК и цитокинины, микроорганизмами может оказывать существенное влияние на морфологию и анатомию корневой системы, способствуя более эффективному поглощению питательных веществ [28].
Арбускулярные микоризные (АМ) грибы, усиливая поглощение питательных веществ растениями-хозяевами, используются для биофортификации Se. Геном арбускулярно микоризных грибов кодирует высокоаффинные транспортеры фосфата, кроме того при симбиотических взаимоотношениях они спо- собны индуцировать специфичные транспортеры фосфата в растении-хозяине, усиливая поглощение и транспорт фосфата. Например, Rhizophagus irregularis и Laccaria bicolor кодируют транспортеры и пермеазы сульфата. Ввиду химического сходства Se и S, Se усваивается в форме селената/селенита и метаболизируется по пути ассимиляции серы, образуя SeCys, SeMet и другие селено-аналоги метаболитов S. Вследствие чего эти микроорганизмы способствуют повышенному накоплению Se в растениях [29].
P. Durán и др. [30] исследовали влияние совместной инокуляции пшеницы селенобактериями Stenotrophomonas sp. B19, Entero-bacter sp. B16, Bacillus sp. R12, Pseudomonas sp. R8 (отдельно и в смеси) и арбускулярным микоризным грибом Glomus claroideum на усвоение Se. Инокуляция значительно повышала концентрацию Se в тканях растений по сравнению с контролем. Независимо от присутствия G. claroideum , обработка пшеницы Enterobacter sp. B16 обеспечивала наибольшее накопление Se в зерне. Наблюдался синергетический эффект от совместного использования PGPR и АМГ на концентрацию Se: совместная инокуляция увеличивала содержание Se в зерне на 23,5 % по сравнению с растениями, обработанными только бактериальным штаммом. Этот синергизм предположительно обусловлен трофическими связями между ними.
Глюконовая и кетоглюконовая кислоты, синтезируемые бактериями филумов Firmi-cutes, Protobacteria и Actinobacteria, играют важную роль в растворении Zn [31]. Вклад в этот процесс также вносят лимонная, яблочная, щавелевая, винная, муравьиная и уксусная кислоты. Увеличение поглощения и накопления Zn в зерне пшеницы, опосредованное продукцией органических кислот, показано на примере Pseudomonas fluorescens Psd [32] и Exiguobacterium aurantiacum [33].
Несмотря на изучение физиологических и молекулярных механизмов микробного обеспечения растений питательными веществами, практическое применение микроорганизмов для управления питанием сельскохозяйственных культур все еще ограничено [34]. Дальнейшие полевые исследования необходимы для эффективной интеграции микробной биофортификации в сельскохозяйственное производство.
Микробная биофортификация представляет собой экологически безопасную, экономически эффективную и устойчивую альтернативу другим стратегиям обогащения сельскохозяйственных культур микроэлементами. Кроме того, внедрение микробных препаратов в сельскохозяйственную практику сопряжено с дополнительными преимуществами, такими как стимуляция роста растений и повышение их устойчивости к стрессам [35].
Выводы
Анализ генетических, агрономических и микробных стратегий биофортификации пшеницы выявил специфические преимущества и ограничения каждого подхода. Генетическая биофортификация, предлагая устойчивое повышение содержания микронутриентов, сталкивается с трудностями практической реализации и потенциальным неприятием генетически модифицированных организмов. Агрономические методы, несмотря на доказанную эффективность, характеризуются высокой стоимостью и негативным воздействием на агроэкосистемы. Микробная биофортификация, представляя собой экологически безопасную альтернативу, требует углубленных исследований для оптимизации и масштабирования. Макси-
Таблица 4
Преимущества и недостатки микробной биофортификации Triticum aestivum L.
Поиск и характеристика новых штаммов ризобактерий, эндофитов и арбускулярных микоризных грибов с повышенной способностью к мобилизации и трансфера микронутриентов является приоритетной задачей. Разработка комплексных микробных инокулянтов с синергетическим действием, а также оптимизация агротехнологических приемов для повышения биодоступности микроэлементов и снижения экологической нагрузки, представляют значительный научный интерес.
Необходимо дополнительное изучение геномных основ механизмов симбиотического взаимодействия ризосферных и эндофитных микроорганизмов с растением-хозяином, проведение масштабных полевых экспериментов, оценка биодоступности микронутриентов из биофортифицированного зерна и анализ социально-экономических последствий внедрения биологически обогащенных культур в питание населения. Стратегической целью является разработка комплексных, устойчивых и экономически обоснованных решений, гарантирующих повышение пищевой ценности пшеницы для обеспечения глобальной продовольственной безопасности.
