Оценка генетических параметров в субпопуляции скота джерсейской породы на основе изучения STR-маркеров и их потенциального влияния на изменчивость показателей развития животных

Автор: Олейник С.А., Лесняк А.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика, протеомика
Статья в выпуске: 2 т.60, 2025 года.
Бесплатный доступ
Изучение взаимосвязи генетических и фенотипических показателей крупного рогатого скота - важное направление селекции и генетики. Сопоставляя эти показатели, можно повысить эффективность разведения животных и обеспечить создание более высокопродуктивного стада. В настоящей работе с использованием STR-маркеров впервые получены данные по генетической структуре ремонтных телок джерсейской породы в условиях Северного Кавказа и показано влияние эффективных аллелей в исследуемых локусах на живую массу в период онтогенеза. Цель работы - изучить и охарактеризовать генетическое разнообразие и структуру субпопуляции коров джерсейской породы на Северном Кавказе по 16 микросателлитным локусам (BM1818, BM1824, BM2113, ETH3, ETH10, ETH225, INRA023, ILSTS006, SPS115, TGLA53, TGLA122, TGLA126, TGLA227, CSSM66, CSRM60, HAUT27) и идентифицировать аллельные варианты, имеющие потенциальное влияние на живую массу ремонтных телок при рождении и в возрасте 6 мес и среднесуточные приросты живой массы за период от рождения до 6 мес. Работу проводили в 2023 году. Объектом исследования были ремонтные телки ( Bos taurus taurus ) джерсейской породы линии Джестер 534585 ( n = 1074), которых разводят в племенном репродукторе ООО «Агроальянс Инвест» (Ставропольский край). Динамику живой массы определяли при взвешивании молодняка при рождении и в 6 мес (ГОСТ Р 57784-2017. М., 2017). Для генетических исследований от животных получали образцы крови. Выделение и очистку нуклеиновых кислот осуществляли на магнитном сорбенте с использованием коммерческого набора М-Сорб-кровь (ООО «НПФ Синтол», Россия). Мультиплексную амплификацию проводили на термоциклере T100 Thermal cycler («Bio-Rad», США) c использованием коммерческого набора Gene Profile Cattle (ООО «НПФ Синтол», Россия). Ампликоны, полученные в результате полимеразной цепной реакции, разделяли методом капиллярного электрофореза на автоматическом генетическом анализаторе Нанофор 05 (ООО «НПФ Синтол», Россия). Полученные данные обрабатывали с использованием программного обеспечения Gene-Marker v.3.0.1 («SoftGenetics LLC», США). Параметры полиморфизма аллелей и генетического разнообразия, включая число средних (NA), эффективных (NE) аллелей на локус, наблюдаемую (HO) и ожидаемую (HE) гетерозиготность, информационный индекс Шеннона (I), индекс фиксации (FIS), анализ основных координат (PCoA), вероятность идентичности (PI) и вероятность случайного совпадения (PISIBS), c2 (ChiSq) для равновесия Харди-Вайнберга, степень свободы (DF), вероятность (Prob) и значимость (Sigif), вычисляли с помощью надстройки GenAlEx v.6.51b2 для Microsoft Excel. Наблюдаемая и ожидаемая гетерозиготность в субпопуляции коров джерсейской породы, разводимых в условиях Северного Кавказа, по обозначенным выше локусам составляла соответственно 0,657±0,036 и 0,621±0,031, что свидетельствует о повышенной генетической адаптивности к изменениям климатических условий. FIS (-0,056±0,011) указывал на неродственное спаривание в субпопуляции. Самая высокая частота встречаемости в выборке была у аллеля 135 локуса ВМ2113 (83 %) и аллеля 117 локуса ETH3 (79 %). Среднее число аллелей на локус составило 6,875±0,446, из них среднее число эффективных аллелей - 2,846±0,171. Индекс Шеннона равнялся 1,183±0,061, что свидетельствует о высокой степени однородности стада. Обнаружено достоверно значимое влияние семи локусов (TGLA227, TGLA53, BM1824, CSSM66, TGLA122, BM1818, ILSTS006) на живую массу молодняка в возрасте 6 мес и среднесуточные приросты живой массы в период от 0 до 6 мес, а также двух локусов (ETH3, ETH10) на живую массу при рождении и в 6 мес. В результате в этих локусах были выделены аллельные варианты, при которых живая масса и ее среднесуточные приросты показывали свои пиковые значения. Профили, полученные в результате микросателлитного анализа, можно использовать в популяционных исследованиях, а также для планирования селекционно-племенной деятельности, поддержания уровня гетерозиготности в популяциях и генетического отслеживания селекционных процессов.
Субпопуляция, джерсейская порода, селекция, генетика, генотипирование, онтогенез, фенотип, живая масса
Короткий адрес: https://sciup.org/142245109
IDR: 142245109 | УДК: 636.2.034:636.082:575.174.015.3 | DOI: 10.15389/agrobiology.2025.2.271rus
Assessment of genetic parameters in a subpopulation of Jersey cattle based on the study of STR markers and their potential impact on the variability of animal development indicators
The study of the relationship between genetic and phenotypic traits in cattle is a key area of breeding and genetics. Comparing these traits can enhance animal breeding efficiency and facilitate the development of more productive herds. Using STR markers, this study presents the first data on the genetic structure of Jersey replacement heifers in the North Caucasus region and demonstrates the influence of specific alleles at the studied loci on live weight during ontogenesis. The aim of the work was to study and characterize the genetic diversity and structure of a Jersey cow subpopulation in the North Caucasus using 16 microsatellite loci (BM1818, BM1824, BM2113, ETH3, ETH10, ETH225, INRA023, ILSTS006, SPS115, TGLA53, TGLA122, TGLA126, TGLA227, CSSM66, CSRM60, HAUT27), and also to identify allelic variants with potential influence on the live weight of replacement heifers at birth and at 6 months of age, and on average daily live weight gains from birth to 6 months. The work was conducted in 2023. The object of the study was replacement heifers (Bos taurus taurus) of the Jersey breed, Jester 534585 line ( n = 1074), bred at the breeding farm Agroalliance Invest LLC in the Stavropol Territory. Live weight dynamics were studied by weighing young stock at birth and at 6 months (GOST R 57784-2017. M., 2017). Blood samples were collected from the animals for genetic studies. Nucleic acid isolation and purification were performed using magnetic sorbent with the commercial kit M-Sorb-blood (Syntol R&D and Production LLC, Russia). Multiplex amplification was carried out on a T100 Thermal Cycler (Bio-Rad, USA) using the commercial Gene Profile Cattle kit (Syntol R&D and Production LLC, Russia). Amplicons obtained from the polymerase chain reaction were separated by capillary electrophoresis on an automated genetic analyzer Nanofor 05 (Syntol R&D and Production LLC, Russia). The obtained data were processed using GeneMarker v.3.0.1 software (SoftGenetics LLC, USA). Parameters of allele polymorphism and genetic diversity, including mean number of alleles (NA), effective number of alleles (NE), observed (HO) and expected (HE) heterozygosity, Shannon's information index (I), fixation index (FIS), Principal Coordinates Analysis (PCoA), probability of identity (PI), probability of identity between siblings (PISIBS), c2 (ChiSq) for Hardy-Weinberg equilibrium, degrees of freedom (DF), probability (Prob), and significance (Sigif), were calculated using the GenAlEx v.6.51b2 add-in for Microsoft Excel. Observed (HO) and expected (HE) heterozygosity in the Jersey subpopulation bred in the North Caucasus conditions for the aforementioned loci were 0.657±0.036 and 0.621±0.031, respectively, suggesting enhanced genetic adaptability to changing environmental conditions. The fixation index (FIS) of -0.056±0.011 indicated outcrossing (non-relative mating) within the subpopulation. The highest allele frequencies in the sample were observed for allele 135 bp of the BM2113 locus (83%) and allele 117 bp of the ETH3 locus (79 %). The average number of alleles per locus (NA) was 6.875±0.446, and the average number of effective alleles (NE) was 2.846±0.171. Shannon's information index (I) was 1.183±0.061, indicating a high degree of herd uniformity. A statistically significant influence was found for 7 loci (TGLA227, TGLA53, BM1824, CSSM66, TGLA122, BM1818, ILSTS006) on the live weight of young stock at 6 months and on average daily gains from 0 to 6 months, as well as for 2 loci (ETH3, ETH10) on live weight at birth and at 6 months. Consequently, allelic variants within these loci were identified that were associated with peak values for live weight and average daily gains. The profiles obtained from the microsatellite analysis can be used in population studies, as well as for planning breeding activities, maintaining heterozygosity levels in populations, and genetically monitoring selection processes.
Текст научной статьи Оценка генетических параметров в субпопуляции скота джерсейской породы на основе изучения STR-маркеров и их потенциального влияния на изменчивость показателей развития животных
Генетическое разнообразие крупного рогатого скота (КРС) служит
Научно-исследовательская работа выполнена в рамках реализации плановых работ по заданию Минсельхоза РФ по разработке генетико-технологической модели стада крупного рогатого скота разных пород для оптимизации производства молока на юге России и продления периода хозяйственного использования коров при высоком и среднем уровнях продуктивности животных.
основой для успешной селекции (улучшения признаков), эффективного управления популяциями (принятия решений в племенной работе) и долгосрочного сохранения пород. Повышенная частота инбридинга (близкородственного скрещивания) или избыточная гомозиготность приводят к потере генетического разнообразия (1, 2).
В России наблюдается тенденция к интенсификации процесса селекции в молочном скотоводстве, направленная на повышение продуктивности сельскохозяйственных животных за счет оптимизации хозяйственно полезных признаков различных пород. Этот процесс включает разработку и внедрение эффективных адаптивных технологий, в том числе способных снизить негативное воздействие антропогенного фактора. Целенаправленная работа по улучшению племенных и продуктивных качеств молочного скота как внутри страны, так и за ее пределами позволила сформировать значительный генетический потенциал, способствующий увеличению продуктивности и устойчивости животных к различным факторам.
Джерсейская порода имеет европейское происхождение и занимает второе место по распространенности (82 страны) среди лучших молочных пород мира (3), чем демонстрирует свою адаптивность к разнообразным климатическим и географическим условиям. При этом она характеризуется высокими показателями содержания жира (5-7 %) и белка (4-5 %) в молоке-сырье, имеет ранний возраст первого осеменения (13-14 мес) и оптимальные технологические параметры вымени с высокой скоростью молокоот-дачи (2,5-3,5 кг/мин). Коровы джерсейской породы, как правило, относительно не крупного размера (средняя масса полновозрастных животных составляет около 410 кг) и имеют более высокий коэффициент молочности (1714 кг молока/100 кг живой массы) в сравнении с широко распространенной голштинской породой, у которой этот же показатель составляет 1240 кг молока/100 кг живой массы (4, 5). При производстве молочных продуктов (сыры, творог, сливочное масло) их процентный выход выше из молока коров джерсейской породы, чем голштинской на 6-10 % (6).
Учитывая все преимущества джерсейской породы КРС, можно сделать вывод о ее потенциале стать одной из ведущих пород в животноводстве. Это касается не только производства жирного молока с высоким содержанием белка, но и создания сыра и сливочного масла.
В соответствии с рекомендациями ФАО, для создания эффективных национальных стратегий по управлению генетическими ресурсами сельскохозяйственных животных крайне важно повсеместно внедрять в практику животноводства микросателлитные эталонные маркеры, такие как короткие тандемные повторы (short tandem repeat, STR) (7).
Согласно отчету ISAG (International Society for Animal Genetics), рекомендуемые 12 STR-маркеров для КРС имеют высокую точность от 97,3 до 99,8 % в тестах на достоверность происхождения .
Практическое значение STR-маркеров для описания генетического разнообразия и структуры джерсейской породы продемонстрировано во множестве публикаций (8, 9). Подтверждение, обнаружение и корректировка происхождения и связанных родственников позволяют создавать более полные родословные, которые, в свою очередь, увеличивают число используемых генетических и фенотипических записей (10, 11).
Микросателлитные локусы представляют собой преимущественно нейтральные молекулярно-генетические маркеры. Однако существует множество свидетельств, указывающих на возможную связь между определенными аллелями в микросателлитных локусах и хозяйственно полезными 272
признаками у сельскохозяйственных животных.
Так, аллель 136 п.н. в микросателлитном локусе BM1500 служит маркером наивысшего удоя, а аллель 138 п.н. в гомозиготном виде этого же локуса связан с повышенным содержанием жира в молоке (12, 13). При исследовании коров голштинской породы наличие гомозиготного аллеля 268 п.н. локуса BM6438 было связано с наиболее высокими надоями (7001 кг молока) (14), в то время как коровы имели низкие показатели молочной продуктивности при аллельных вариантах 256/258 (6478 кг молока) и 258/268 (6389 кг молока) в этом локусе. Локус BM1905 по генотипам 178/192 и 188/188 достоверно ассоциирован со средним значением соматических клеток в молоке, также отмечены локус MB026 c генотипами 228/252 и 238/252, локус BM1258 c генотипом 110/120, локус BM1443 с генотипами 154/182 и 154/178 (15).
Аналогичные исследования в мясном скотоводстве показали, что наличие аллеля 142 п.н. в микросателлитном локусе BM2113 обеспечивает больший выход чистого мяса, а аллель 171 п.н. в этом же локусе положительно связан с толщиной филейной части туши (16). Животные помеси пород Ангус ½ Брахман с аллельным вариантом 220/222 в локусе ETH10 имели более высокие характеристики мраморности мяса и большую убойную массу (17). У КРС южной мясной породы аллель 83 п.н. в локусе TGLA227 и аллель 258 п.н. в локусе BM1818 служат маркерами интенсивной скорости роста живой массы у молодняка от рождения до отъема, в то время как у ремонтных телок, имеющих аллель 141 п.н. в локусе BM2113, отмечается медленный рост живой массы в разные возрастные периоды (18).
Была отмечена положительная связь с основными промерами у КРС с рядом микросателлитных локусов. Так, локус BM2113 показал максимальное пиковое значение в связи с шириной груди (19), гомозиготность аллеля 205 п.н. в локуcе IDVGA46 в значительной степени связана с развитием мускулатуры и скелетом, животные имели больший размер тела (20).
При этом остается малоизученной возможная взаимосвязь между генотипом и динамикой живой массы у коров молочного направления (21).
Неодинаковая интенсивность роста у разных животных в период от рождения до 6 мес может как негативно, так и положительно повлиять на развитие молочных желез и, соответственно, на формирование молочной продуктивности (22, 23). У коров джерсейской породы, разводимых в условиях Северного Кавказа, при среднесуточном приросте 811-961 г в период от 0 до 6 мес наблюдались наиболее высокие удои по результатам 305 сут первой лактации в сравнении со сверстницами, у которых среднесуточный прирост за аналогичный период был в диапазоне 456-806 г (24).
Результаты изучения влияния отдельных микросателлитных локусов на показатели интенсивности роста живой массы в период от 0 до 6 мес можно использовать как дополнительный инструмент в маркер-ориентиро-ванной селекции, что позволит реализовывать весь заложенный генетический потенциал животных джерсейской породы. Такой подход в селекции КРС молочных пород сможет повысить показатель эффективности национального молочного скотоводства.
В настоящей работе с использованием STR-маркеров впервые получены данные по генетической структуре ремонтных телок джерсейской породы в условиях Северного Кавказа и показано влияние эффективных аллелей в исследуемых локусах на живую массу в период онтогенеза.
Цель работы — изучить и охарактеризовать генетическое разнообразие и структуру субпопуляции коров джерсейской породы на Северном Кавказе по 16 микросателлитным локусам (BM1818, BM1824, BM2113, ETH3,
ETH10, ETH225, INRA023, SPS115, TGLA53, TGLA122, TGLA126, TGLA227, CSSM66, CSRM60, HAUT27), а также идентифицировать аллельные варианты, имеющие потенциальное влияние на живую массу ремонтных телок при рождении и в возрасте 6 мес и среднесуточные приросты живой массы за период от рождения до 6 мес.
Методика. Работу проводили в 2023 году. Объектом исследования были ремонтные телки ( Bos taurus taurus ) джерсейской породы линии Джестер 534585 ( n = 1074), которых разводят в племенном репродукторе ООО «Агроальянс Инвест» (Ставропольский край, Александровский р-н). Эти животные — потомки коров, завезенных на Северный Кавказ в 2017 году из племенных предприятий Дании.
Все животные содержались в равных условиях и на однотипном рационе для выращивания молодняка на уровне стандарта породы, рассчитанном для конкретного стада, что позволило не учитывать влияние кормления и условий среды на результаты исследования.
Динамику живой массы изучали посредством взвешивания молодняка при рождении и в 6 мес (ГОСТ Р 57784-2017. М., 2017).
Биоматериал от ремонтных телок (образцы цельной крови) отбирали из яремной вены в вакуумные пробирки с антикоагулянтом К3ЭДТА. Генетические исследования биоматериала проводили в ФГБОУ ВО Ставропольский ГАУ (номер госрегистрации в племенном регистре РФ ¹ 262704803000, Свидетельство о регистрации в государственном племенном регистре, серия ПЖ 77; ¹ 010649).
Выделение и очистку нуклеиновых кислот осуществляли на магнитном сорбенте с использованием коммерческого набора М-Сорб-кровь (ООО «НПФ Синтол», Россия).
Мультиплексную амплификацию проводили на термоциклере T100 Thermal cycler («Bio-Rad», США) c использованием коммерческого набора Gene Profile Cattle (ООО «НПФ Синтол», Россия), содержащего праймеры для амплификации 16 микросателлитных (STR) локусов. Из них 12 локусов (BM1818, BM1824, BM2113, ETH3, ETH10, ETH225, INRA023, SPS115, TGLA53, TGLA122, TGLA126, TGLA227) рекомендованы Международным обществом генетики животных (International Society for Animal Genetics, ISAG), а 3 локуса (CSSM66, CSRM60, ILSTS006) — Продовольственной и сельскохозяйственной организацией ООН (Food and Agriculture Organization, FAO) для генетических исследований домашних животных. Дополнительно был включен высокополиморфный локус HAUT27. Амплификацию проводили в следующем режиме: 2 мин при 95 ° С (денатурация); 5 с при 95 ° С, 1 мин при 60 ° C, 20 с при 72 ° C (отжиг) (30 циклов); 10 мин при 59 ° C (элонгация).
Ампликоны, полученные в результате полимеразной цепной реакции, разделяли методом капиллярного электрофореза на автоматическом генетическом анализаторе Нанофор 05 (ООО «НПФ Синтол», Россия).
Полученные данные обрабатывали с использованием программного обеспечения GeneMarker v.3.0.1 («SoftGenetics LLC», США).
Параметры полиморфизма аллелей и генетического разнообразия, включая число средних (N a ), эффективных (N e ) аллелей на локус, наблюдаемую (H o ) и ожидаемую (H e ) гетерозиготность, информационный индекс Шеннона (I), индекс фиксации (F is ), анализ основных координат (PCoA), вероятность идентичности (PI) и вероятность случайного совпадения (PI sibs ), X2 (ChiSq) для равновесия Харди-Вайнберга, степень свободы (DF), вероятность (Prob) и значимость (Sigif), вычисляли с помощью надстройки GenAlEx v.6.51b2 для Microsoft Excel (25).
Однофакторный дисперсионный анализ (ANOVA) влияния аллельных вариантов микросателлитных локусов на живую массу и ее среднесуточный прирост проводили в программе STATISTICA 10.0 («StatSoft Inc.», США). Определяли средние значения показателей ( M) и стандартные ошибки средних (±SEM).
Результаты. Все особи из выборки ( n = 1074), рожденные в период с 2021 по 2022 год, имели подтвержденное происхождение по материнской и отцовской линиям с вероятностью 99,9 %. Значительный размер выборки генотипированных особей разного происхождения позволяет сделать вывод о том, что обнаруженные закономерности характерны для джерсейской породы.
Анализ основных компонент (PCoA) показал сходство индивидуумов между собой в субпопуляции (рис. 1), что позволяет сделать заключение о принадлежности всех животных в выборке к генотипу одной породы.
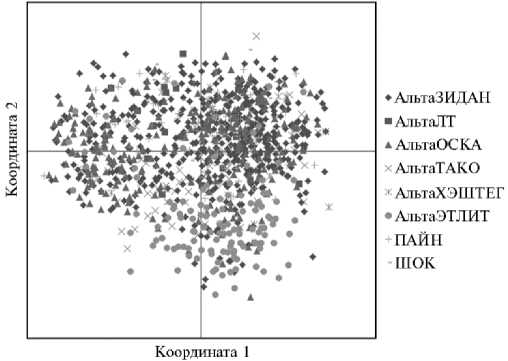
Рис. 1. Проекция индивидуальных генотипов животных, полученных в субпопуляции ремонтных телок джерсейской породы, с распределением по быкам-отцам на плоскости двух координат по данным PCoA-анализа ( n = 1074, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год) .
Животные в выборке происходили от 8 быков-отцов линии быка Джестер 534585 (см. рис. 1). На момент исследования число ремонтного молодняка этой линии преобладало над сверстницами других линий. Тем самым было дополнительно исключено линейное влияние на полученные результаты. При дальнейшем анализе субпопуляции джерсейского скота учитывали совокупные данные по выборке без разделения на быков-отцов.
Анализ распределения частот генотипов в соответствии с генетическим балансом Харди-Вайнберга по всему набору данных (табл. 1) показал небольшие отклонения от состояния генетического равновесия в некоторых локусах исследуемой субпопуляции (TGLA227, TGLA122).
1. Результаты теста χ2 на соответствие равновесию Харди-Вайнберга для 16 STR-локусов в субпопуляции ремонтных телок ( Bos taurus taurus ) джерсей-ской породы ( n = 1074, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год)
|
Локус |
DF |
ChiSq |
Prob |
Signif |
|
TGLA227 |
45 |
25,879 |
0,990 |
ns |
|
BM2113 |
15 |
1080,876 |
0,000 |
* |
|
TGLA53 |
55 |
1113,041 |
0,000 |
* |
|
ETH10 |
28 |
1128,056 |
0,000 |
* |
|
CSRM60 |
45 |
298,469 |
0,000 |
* |
|
SPS115 |
21 |
82,411 |
0,000 |
* |
Продолжение таблицы 1
|
TGLA122 |
21 |
23,655 |
0,310 |
ns |
|
BM1818 |
10 |
55,646 |
0,000 |
* |
|
HAUT27 |
21 |
48,506 |
0,001 |
* |
|
CSSM66 |
66 |
1215,593 |
0,000 |
* |
|
BM1824 |
15 |
1180,518 |
0,000 |
* |
|
ETH3 |
10 |
1088,698 |
0,000 |
* |
|
TGLA126 |
10 |
61,709 |
0,000 |
* |
|
ETH225 |
15 |
46,299 |
0,000 |
* |
|
INRA023 |
28 |
60,325 |
0,000 |
* |
|
ILSTS006 |
45 |
106,502 |
0,000 |
* |
Примечание. DF — число степеней свободы, ChiSq — значение χ2, Prob — p-значение (вероятность), Signif — уровень значимости; ns — различия несущественны (p > 0,05).
* р < 0,001.
Отклонение от генетического равновесия может быть обусловлено смещением частот аллелей относительно предыдущего поколения. К этому можно добавить еще и генетический отбор животных в сторону улучшения селекции по хозяйственно полезным признакам (26, 27). Учитывая результаты, представленные выше, мы определили полиморфизм микросателлит-ных локусов субпопуляции коров джерсейской породы (табл. 2).
-
2. Частота встречаемости аллельных вариантов микросателлитных локусов в субпопуляции ремонтных телок джерсейской породы ( Bos taurus taurus ), разводимых в условиях Северного Кавказа ( n = 1074, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год)
3. Гетерозиготность, F-статистика и полиморфизм по микросателлитным локусам в субпопуляции ремонтных телок джерсейской породы (
Bos taurus taurus
), разводимых в условиях Северного Кавказа
(
n
= 1074, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год)
Локус Аллель Вариативность аллелей в локусах
|
CSSM66 |
Размер, п.н. Частота |
159 0,001 |
179 0,003 |
181 0,061 |
183 0,006 |
185 0,133 |
187 0,052 |
189 0,018 |
193 0,606 |
197 0,114 |
199 0,005 |
|
TGLA53 |
Размер, п.н. |
153 |
160 |
162 |
168 |
170 |
172 |
182 |
186 |
||
|
Частота |
0,001 |
0,240 |
0,027 |
0,412 |
0,307 |
0,001 |
0,008 |
0,001 |
|||
|
INRA023 |
Размер, п.н. |
198 |
200 |
202 |
206 |
208 |
210 |
214 |
218 |
||
|
Частота |
0,312 |
0,010 |
0,004 |
0,466 |
0,131 |
0,002 |
0,053 |
0,022 |
|||
|
ILSTS006 |
Размер, п.н. |
284 |
286 |
288 |
290 |
292 |
294 |
296 |
298 |
||
|
Частота |
0,006 |
0,161 |
0,078 |
0,001 |
0,431 |
0,288 |
0,028 |
0,006 |
|||
|
CSRM60 |
Размер, п.н. |
92 |
94 |
96 |
98 |
100 |
102 |
104 |
|||
|
Частота |
0,304 |
0,003 |
0,177 |
0,025 |
0,004 |
0,272 |
0,212 |
||||
|
ETH10 |
Размер, п.н. |
198 |
213 |
215 |
217 |
219 |
221 |
223 |
|||
|
Частота |
0,001 |
0.016 |
0,347 |
0,225 |
0,128 |
0,279 |
0,004 |
||||
|
HAUT27 |
Размер, п.н. |
140 |
146 |
148 |
150 |
152 |
154 |
156 |
|||
|
Частота |
0,001 |
0,246 |
0,286 |
0,027 |
0,001 |
0,438 |
0,001 |
||||
|
TGLA227 |
Размер, п.н. |
77 |
81 |
85 |
91 |
93 |
97 |
101 |
|||
|
Частота |
0,024 |
0,417 |
0,001 |
0,188 |
0,342 |
0,009 |
0,019 |
||||
|
SPS115 |
Размер, п.н. |
248 |
252 |
254 |
256 |
260 |
261 |
||||
|
Частота |
0,264 |
0,461 |
0,068 |
0,003 |
0,186 |
0,017 |
|||||
|
TGLA122 |
Размер, п.н. |
143 |
149 |
151 |
161 |
169 |
181 |
||||
|
Частота |
0,453 |
0,180 |
0,287 |
0,005 |
0,072 |
0,002 |
|||||
|
BM1824 |
Размер, п.н. |
178 |
180 |
182 |
183 |
188 |
190 |
||||
|
Частота |
0,005 |
0,578 |
0,143 |
0,001 |
0,240 |
0,034 |
|||||
|
ETH225 |
Размер, п.н. |
140 |
144 |
146 |
148 |
150 |
152 |
||||
|
Частота |
0,088 |
0,397 |
0,001 |
0,307 |
0,200 |
0,008 |
|||||
|
BM2113 |
Размер, п.н. |
125 |
129 |
133 |
135 |
137 |
|||||
|
Частота |
0,001 |
0,044 |
0,105 |
0,826 |
0,023 |
||||||
|
BM1818 |
Размер, п.н. |
258 |
260 |
262 |
264 |
266 |
|||||
|
Частота |
0,089 |
0,110 |
0,174 |
0,074 |
0,554 |
||||||
|
TGLA126 |
Размер, п.н. |
115 |
117 |
121 |
123 |
||||||
|
Частота |
0,353 |
0,466 |
0,092 |
0,088 |
|||||||
|
ETH3 |
Размер, п.н. |
115 |
117 |
121 |
125 |
||||||
|
Частота |
0,001 |
0,789 |
0,004 |
0,206 |
В результате проведенного анализа ДНК у животных джерсейской породы было выявлено 10 аллелей в локусе CSSM66 длиной от 159 до 199 п.н., по 8 аллелей в локусах TGLA53 (153-186 п.н), INRA023 (198-218 п.н.), ILSTS006 (284-298 п.н.), что свидетельствует о высокой полиморфности указанных локусов. Наименьший полиморфизм был обнаружен в локусах ETH3 (115-125 п.н.) и TGLA126 (115-123 п.н.). С наибольшей частотой в нашей выборке животных встречались аллель 135 локуса BM2113 (83 %) и 276
аллель 117 локуса ETH3 (79 %).
Чтобы оценить степень генетического разнообразия популяций и видов, обычно обращают внимание на два ключевых показателя: уровень полиморфизма и степень гетерозиготности (28).
В локусах BM1818, ETH3 и TGLA126 наблюдалось минимальное число аллелей, равное 5. Максимальное значение, равное 12, было зафиксировано в локусе CSSM66. Среднее значение числа аллелей на локус составляло 7,688. Значение эффективных аллелей на локус (N е ) варьировало от 1,436 (BM2113) до 4,102 (CSRM60) (табл. 3).
Информационный индекс Шеннона (I) — еще один показатель, отражающий степень полиморфизма и генетическое разнообразие исследуемой субпопуляции, учитывая ее однородность. Он считается сильным показателем эволюционного потенциала популяции (29), однако служит косвенным показателем разнообразия без чрезмерного акцента на редкие и доминирующие аллели, если размер выборки мал ( n < 50) (30). Чем меньше видовое разнообразие, тем больше индекс Шеннона стремится к нулю (31). Его минимальные значения были выявлены нами в локусах ETH3 (0,543) и BM2113 (0,629), а максимальное значение — в локусе CSRM60 (1,497) при среднем значении по выборке 1,198, что означает высокую степень однородности в исследуемой субпопуляции животных.
В ряде исследований для характеристики популяции используют данные наблюдаемой (H o ) и ожидаемой (H e ) гетерозиготности (32-34). В нашем случае H o находилась в пределах от 0,313 для локуса BM2113 до 0,790 для локуса CSRM60. Средний уровень (выше 0,5) ожидаемой гетерозиготности был зафиксирован в 14 локусах. В числе исключений находились локусы ETH3 и BM2113, в которых этот показатель достиг минимальных значений — 0,335 и 0,304.
Согласно утверждениям некоторых исследователей, уменьшение гетерозиготности способно вызвать снижение приспособленности организмов к изменению климатических условий, поэтому такой индикатор обладает явными экологическими эффектами (35, 36). Индекс фиксации (Fis) — это статистический показатель, который используется для оценки степени инбридинга (близкородственного скрещивания) в субпопуляциях. Он поз- воляет определить, существует ли недостаток или избыток гетерозигот в популяции.
Гетерозиготность — это наличие разных аллелей одного гена у одной особи. Инбридинг приводит к уменьшению генетического разнообразия и увеличению гомозиготности в популяции. Если значение F is > 0, то это говорит о недостатке гетерозигот, что служит признаком родственного скрещивания. То есть в популяции преобладают особи, которые имеют одинаковые аллели одного гена. Значение F is < 0 указывает на избыток гетерозигот и неродственное скрещивание. Это означает, что в популяции преобладают особи, которые имеют разные аллели одного гена. При значении F is = 0 происходит случайное скрещивание, при котором вероятность образования гомозигот и гетерозигот одинакова (37). Следовательно, F is служит важным инструментом для изучения генетической структуры популяций и оценки последствий инбридинга.
Во всех 16 локусах был отмечен сдвиг баланса в сторону избытка гетерозигот, что указывает на низкий процент инбридинга в исследуемой выборке.
Среди изучаемых локусов максимальный уровень полиморфизма обнаружили в CSRM60 (N a = 10, N e = 4,102, I = 1,497). Стоит отметить, что по числу аллелей на локус первое место занимал CSSM66 (N a = 12), но при этом он имел значительно меньший уровень полиморфизма (N e = 2,473, I = 1,306). Максимальное генетическое разнообразие отмечали в локусе CSSM66 (H o = 0,790, H e = 0,756).
В исследованиях Н.В. Ковалюк с соавт. (38) при изучении генетической структуры джерсейского скота южных регионов России высокий уровень полиморфизма был установлен в локусах BM1818 (N a = 6, N e = 5), ETH10 (N A = 6, N E = 4), INRA023 (N A = 7, N E = 3), TGLA122 (N A = 7, N e = 4), TGLA227 (N a = 9, N e = 5), что согласуется с нашими данными, за исключением локуса BM1818, который в настоящем исследовании содержал наименьшее количество аллелей (N a = 5, N e = 2,7). Наблюдаемая (H o ) и ожидаемая (H e ) гетерозиготность составляла соответственно 0,62 и 0,60 при F is - 0,03. В Северной Америке при изучении генетической структуры джерсейского скота ( n = 2922) установили H o = 0,26 и H e = 0,36, что согласуется с эффективным размером популяции в сравнении с голштинской и бурой швицкой породой в этом регионе (39). При исследовании популяции джерсейской породы на острове Джерси (место происхождения породы) средние показатели H o и H e равнялись 0,63 и 0,64, при этом наиболее полиморфными микросателлитными локусами были TGLA227 (N a = 11), ILSTS006 (N a = 7), BM1818 (N a = 6), HEL5 (N a = 6) (40). Авторы более ранних работ по изучению генетической структуры популяции джерсейской породы установили значение H e = 0,41 (41), 0,63 (42).
Проведя сравнительный анализ результатов наших исследований и данных, представленных в работах вышеупомянутых авторов, можно сделать вывод, что генетическое разнообразие в породе обусловлено совокупностью сложных демографических факторов, и текущая или недавняя численность популяции лишь один из них.
Не менее важным аспектом в изучении генетической структуры субпопуляций можно считать достоверность происхождения. Так, доля ошибок в идентификации производителя в племенных и непородистых стадах может варьировать от нескольких до 22 % (43, 44). Правильная информация о родословной имеет первостепенное значение для успешной программы разведения, и ее важность для точной генетической оценки молочного скота с каждым годом увеличивается (45). Ошибка в родословной, например не- правильно идентифицированный производитель, отрицательно повлияет на родословную полученной от него коровы и всех ее родственников.
Для исключения вероятности случайного совпадения генотипов мы сравнили эффективность индивидуальной идентификации особей (рис. 2).
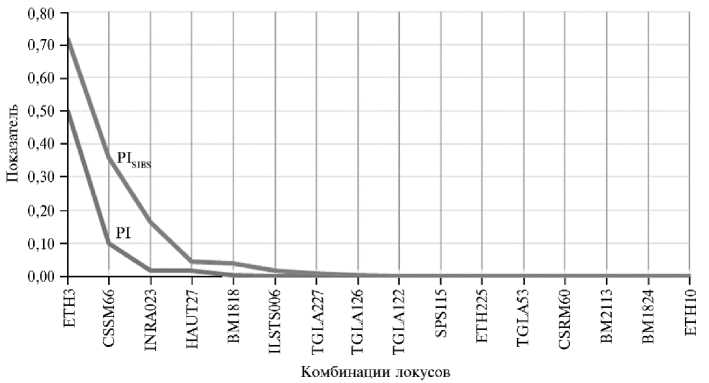
Рис. 2. Вероятность идентичности (PI) и случайного совпадения (PI SIBS ) для возрастающих комбинаций из 16 локусов в субпопуляции ремонтных телок джерсейской породы ( Bos taurus taurus ), разводимых в условиях Северного Кавказа ( n = 1074, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год).
Анализ зависимости вероятности идентичности (PI) и вероятности случайного совпадения (PI sibs ) от числа используемых локусов позволяет сделать вывод о том, что при применении восьми локусов вероятность случайного совпадения генотипов становится практически нулевой. Это подтверждают и другие исследования, в которых также было установлено, что для надежной идентификации происхождения необходимо использовать не менее восьми микросателлитных локусов (46). Применение всех шестнадцати локусов обеспечивает высокую точность результатов и их статистическую значимость.
В научной литературе встречаются исследования влияния аллельных вариантов микросателлитных локусов на показатели продуктивности КРС (26, 47). На основании этого мы изучили влияние аллельного разнообразия в каждом локусе на динамику живой массы ремонтного молодняка джер-сейской породы, что позволило идентифицировать аллели, которые достоверно связаны с данным показателем.
Из всей выборки ( n = 1074) были отобраны животные, достигшие возраста 6 мес ( n = 651). Зависимой переменной были обозначены показатели живой массы при рождении, в 6 мес и среднесуточный прирост за период 0-6 мес. Предиктором служил исследуемый локус. Мы исключили аллельные варианты во всех 16 локусах, которые встречались у менее 5 % животных.
Из 16 локусов мы выявили 7 (TGLA227, TGLA53, BM1824, CSSM66, TGLA122, BM1818, ILSTS006), которые имели достоверно значимую связь с показателем живой массы в 6 мес и среднесуточным приростом за период от 0 до 6 мес, а также 2 локуса (ETH3, ETH10) с достоверно значимой связью с живой массой при рождении и в 6 мес.
Анализ данных показал, что при содержании в микросателлитном профиле животных указанных аллельных вариантов (табл. 4, 5) показатели живой массы при рождении, в 6 мес и среднесуточные приросты в период от рождения до 6 мес демонстрировали свои пиковые значения по всей выборке ремонтного молодняка.
-
4. Пиковые значения живой массы при рождении и в 6 мес у ремонтных телок джерсейской породы ( Bos taurus taurus ), разводимых в условиях Северного Кавказа, в зависимости от аллельных вариантов в микросателлитных локусах ( n = 651, M ±SEM, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год)
-
5. Пиковые значения среднесуточных приростов живой массы за период 0-6 мес у ремонтных телок джерсейской породы ( Bos taurus taurus ), разводимых в условиях Северного Кавказа, в зависимости от аллельных вариантов в мик-росателлитных локусах ( n = 651, M ±SEM, племенной репродуктор ООО «Агроальянс Инвест», Ставропольский край, 2023 год)
-
6. QTL и фланкирующие их микросателлитные локусы, которые связаны с живой массой и интенсивностью роста молодняка различных пород крупного рогатого скота и зебу
Хромосома1 Признак
Порода
рЛокусы, фланкирующие участок QTLр
Ссылка
BTA1
BWT
Брахман ½ Герефорд
BMS1789—UWCA46—BMS4014
(48)
BTA2
BWT
Ангус ½ Брахман
BM2113—OarFCB11
(17)
BTA5
BWT
Ангус ½ Брахман
CSSM34—RM500—ETH10 а
(17)
BTA5
ADG1
Bos taurus (линия M1)
ETH10 а—IGF1—BM1819RM29
(49)
BTA14
ADG1
B. taurus (линия M1)
CSSM66 а—BMS1747
(50)
BTA21
BWT
Брахман ½ Ангус
BMS2815—TGLA337—TGLA122 а
(51)
BTA21
BWT
Ангус ½ Брахман
TGLA337—TGLA122 а—CSSM18
(17)
BTA23
BWT
B. taurus (линия M1)
RM185—BM1818 а
(50)
BTA23
ADG
B. taurus (линия M1)
RM185—BM1818 а
(50)
|
Локус |
Аллельный вариант |
Число животных, гол. |
Показатель |
|
|
среднесуточный прирост, г |
F-критерий |
|||
|
TGLA227 |
93/93 |
56 |
820,33±7,03* |
3,21 |
|
TGLA53 |
160/160 |
37 |
819,36±8,97* |
3,01 |
|
ETH10 |
215/215 |
73 |
806,62±6,21 |
2,07 |
|
BM1824 |
180/188 |
211 |
812,79±4,05*** |
12,60 |
|
CSSM66 |
187/193 |
29 |
816,09±13,43* |
2,19 |
|
TGLA122 |
151/151 |
44 |
817,67±8,49*** |
4,82 |
|
BM1818 |
258/262 |
28 |
823,41±11,42* |
2,40 |
|
ETH3 |
125/125 |
16 |
821,87±16,62 |
2,21 |
|
ILSTS006 |
292/294 |
157 |
806,36±5,23** |
3,83 |
|
Средний показатель по выборке |
799,01±2,66 |
|||
*, ** и *** Различия средних значений среднесуточного прироста для аллельного варианта по сравнению с другими аллельными вариантами конкретного локуса статистически значимы соответственно при p < 0,05, p < 0,01 и p < 0,001.
В настоящее время актуальным остается изучение влияния локусов количественных признаков (QTL) на фенотипические показатели у сельскохозяйственных животных.
Примечание. BWT — живая масса при рождении, ADG — среднесуточный прирост живой массы от рождения до отъема; а — STR-локусы, которые использованы в настоящей работе на ремонтных телках джерсейской породы.
Обнаружены QTL, расположенные на разных хромосомах, которые 280
положительно связаны с показателями живой массы животных на разных этапах развития и фланкированные микросателлитными локусами, часть которых была задействована в наших исследованиях (табл. 6).
Помимо этого, показано, что у КРС и зебу существуют как положительные, так и отрицательные взаимосвязи между аллельными вариантами в микросателлитах и такими параметрами, как живая масса тела и среднесуточные приросты на разных этапах развития животных (табл. 7).
7. STR-локусы, которые связаны с живой массой и интенсивностью роста молодняка различных пород крупного рогатого скота и зебу
|
Локус |
Порода |
Признак |
Ссылка |
|
ETH10 |
Брахман ½ Ангус |
BWT |
(52) |
|
ETH10 |
Герефорд |
BWT |
(53) |
|
IGF1-(AT)n |
Bos indicus ½ B. taurus |
BWT |
(54) |
|
BMS1248 |
Баллийский крупный рогатый скот |
BW |
(55) |
|
ETH10 |
Ангус |
WW |
(52) |
|
TGLA227 |
Южная мясная |
ADG |
(18) |
|
BM1818 |
Южная мясная |
ADG |
(18) |
|
CSSM66 |
B. taurus (линия M1) |
ADG |
(50) |
|
Примечание. |
BWT — живая масса при рождении, WW — |
живая масса во время отъема, |
BW — масса |
|
тела, ADG — среднесуточный прирост живой массы от рождения до отъема. |
|||
Так, K.L. DeAtley с соавт. (52) установили, что микросателлитный локус ETH10 тесно связан с показателем живой массы при рождении у помесных животных пород брахман и ангус и во время отъема у молодняка породы ангус, что подтвердилось в исследовании A. Rogberg-Mu n oz с соавт. (53) на герефордской породе и в нашей работе с джерсейским скотом.
P.C. Andrade с соавт. (54) отмечали, что микросателлитный локус гена IGF1 связан с фенотипической изменчивостью на ранней фазе роста у КРС породы канчим: аллель 225 п.н. в этом локусе имел положительное влияние на показатели живой массы, а аллель 231 п.н., наоборот, отрицательное влияние. При изучении баллийского крупного рогатого скота I.K. Puja с соавт. (55) пришли к заключению, что генотип 130/160 микросател-литного локуса BMS1248 имеет положительную корреляцию с живой массой тела, ростом и длиной, в то время как генотип 130/130 этого же локуса отрицательно проявляется на этих фенотипических показателях.
Аллель 198 п.н. в микросателлитном локусе CSSM66 имел положительную связь со среднесуточными приростами массы тела от рождения до отъема у коммерческой линии М1 Bos taurus (50), что согласуется с результатами, полученными в нашей работе для генотипа 187/193 локуса CSSM66, ассоциированного с более высокой живой массой в 6 мес и среднесуточными приростами (см. табл. 4, 5).
Генотип 83/83 в микросателлитном локусе TGLA227 и генотип 258/258 в микросателлитном локусе BM1818 оказывали положительное влияние на среднесуточные приросты живой массы в период от рождения до отъема у южной мясной породы, в то время как их отсутствие в локусах имело отрицательный эффект (18). Это частично согласуется с нашими результатами, где гетерозиготный генотип 258/262 в локусе BM1818 был связан с пиковыми значениями живой массы и ее приростов (см. табл. 4, 5), а в локусе TGLA227 таким эффектом обладал гомозиготный генотип 93/93 (см. табл. 4, 5). Скорее всего, это можно отнести к породным особенностям.
Анализ научных публикаций за последние два десятилетия показывает, что вопрос влияния микросателлитных локусов и их аллельных вариантов на живую массу и среднесуточные приросты у молодняка изучался в основном на мясных породах, однако этот вопрос актуален и для молочного скотоводства, поскольку интенсивность роста ремонтных телок напрямую влияет на их будущую молочную продуктивность (12-15, 23).
В связи с этим результаты нашей работы актуальны и дают научный задел для дальнейшего изучения влияния микросателлитных локусов на фенотипические признаки у пород молочного направления. Профили, полученные в результате микросателлитного анализа ДНК животных джерсей-ской породы, разводимых в условиях Северного Кавказа, можно использовать в популяционных исследованиях и в практической работе с джерсей-ским скотом, а также этот инструмент может найти применение для разработки селекционных и племенных стратегий, анализа генетического состава популяций, поддержания уровня гетерозиготности у групп особей и мониторинга генетических изменений в процессе селекции.
В будущих поколениях генетическая эволюция будет в значительной степени определяться наличием достаточного уровня генетического разнообразия. В связи с этим углубленное понимание текущего состояния генофонда и структуры джерсейской породы приобретает существенное значение для всей индустрии производства молочных продуктов.
Таким образом, степень генетического разнообразия изученной субпопуляции коров джерсейской породы, разводимых в условиях Северного Кавказа, была достаточно высокой, на что указывает наблюдаемая (0,657) и ожидаемая гетерозиготность (0,621). При этом индекс фиксации (F is ) был равен - 0,056, что свидетельствует о неродственном спаривании при высокой степени однородности стада по индексу Шеннона (1,183). Использование 16 локусов для определения достоверности происхождения позволяет подтвердить родство животных с вероятностью выше 99,9 %, учитывая, что среднее количество аллелей на локус в субпопуляции было равно 6,875, из них среднее число эффективных аллелей составило 2,846. При этом самая высокая частота встречаемости в выборке оказалась у аллеля 135 локуса ВМ2113 (83 %) и аллеля 117 локуса ETH3 (79 %). Обнаружены генотипы для 7 локусов (TGLA227, TGLA53, BM1824, CSSM66, TGLA122, BM1818, ILSTS006), которые имели достоверно значимое влияние на показатели живой массы молодняка джерсейской породы в 6 мес и среднесуточные приросты живой массы в период от 0 до 6 мес, а также 2 локуса (ETH3, ETH10), которые достоверно связаны только с показателем живой массы при рождении и в 6 мес.
