Оценка устойчивости сортов амурской сои к цистообразующей нематоде

Автор: Тимкин П.Д., Линская П.М., Зажигалкина А.С., Пензин А.А.
Рубрика: Агрохимия, агропочвоведение, защита и карантин растений
Статья в выпуске: 1 (205), 2026 года.
Бесплатный доступ
Целью работы являлась молекулярно-генетическая оценка устойчивости четырёх сортов сои амурской селекции (Сентябринка, ВНИИС 1, Нега 1, Куханна) к соевой цистообразующей нематоде (SCN) Heterodera glycines с использованием метода ARMS-ПЦР. Проведено высокоспецифичное генотипирование однонуклеотидных полиморфизмов (SNP) в ключевых генах устойчивости Rhg1 и Rhg4. Результаты выявили чёткую генетическую дифференциацию сортов. Установлено, что сорта Сентябринка и ВНИИС 1 обладают гетерозиготным профилем, сочетающим аллели, ассоциированные с Peking-аллелем, и PI 88788-аллель по обоим локусам (Rhg1 и Rhg4), что прогнозирует потенциальную устойчивость к патогену. Однако высокий уровень гетерозиготности (92–96 %) указывает на их генетическую неоднородность и необходимость дальнейшего отбора стабильных гомозиготных форм для закрепления признака. В отличие от них сорта Нега 1 и Куханна являются гомозиготными по восприимчивому аллелю локуса Rhg1, что определяет их фенотипическую восприимчивость к SCN, несмотря на гетерозиготность по гену Rhg4. Практическая значимость работы состоит в валидации метода ARMS-ПЦР для раннего и точного генотипирования и создании научной основы для ускоренной селекции. Метод эффективен для скрининга селекционного материала и отбора перспективных генотипов, что позволяет сократить сроки создания устойчивых сортов сои, адаптированных к условиям Дальневосточного региона. Выявленные образцы с ценным генетическим профилем рекомендованы в качестве исходного материала для селекционных программ.
Соя, соевая цистообразующая нематода, ARMS-ПЦР, гены устойчивости, Rhg1, Rhg4, молекулярная селекция, генотипирование
Короткий адрес: https://sciup.org/142247793
IDR: 142247793 | УДК: 633.853.52:631.522 | DOI: 10.25230/2412-608X-2026-1-205-107-114
Assessment of the resistance of Amur soybean varieties to cyst nematodes
The aim of the work was to conduct a molecular genetic assessment of the resistance of four Amurbred soybean varieties (Sentyabrinka, VNIIS 1, Nega 1, Kukhanna) to the soybean cyst nematode (SCN) Heterodera glycines using the ARMSPCR method. Highspecificity genotyping of single nucleotide polymorphisms (SNPs) in the key resistance genes Rhg1 and Rhg4 was performed. The results revealed clear genetic differentiation between the varieties. The varieties Sentyabrinka and VNIIS 1 were found to have a heterozygous profile combining alleles associated with the Pekingallele and PI 88788 allele at both loci (Rhg1 and Rhg4), which predicts potential resistance to the pathogen. However, the high level of heterozygosity (92–96%) indicates their genetic heterogeneity and the need to further select stable homozygous forms to fix the trait. In contrast, the varieties Nega 1 and Kukhanna are homozygous for the susceptible allele of the Rhg1 locus, which determines their phenotypic susceptibility to SCN despite being heterozygous for the Rhg4 gene. The practical significance of this work lies in validating the ARMSPCR method for early and accurate genotyping and the development of a scientific basis for accelerated breeding. This method is effective for screening breeding material and selecting promising genotypes, reducing the time required to develop SCNresistant soybean varieties adapted to the conditions of the Far Eastern region. The identified samples with valuable genetic profiles are recommended as parent material for breeding programs.
Текст научной статьи Оценка устойчивости сортов амурской сои к цистообразующей нематоде
Научная статья УДК 633.853.52
Введение. Соевая цистообразующая нематода (SCN) Heterodera glycines Ichinohe является одним из наиболее опасных патогенов сои в мировом масштабе, в том числе в Амурской области, вызывая гетеродероз [1]. В отличие от большинства болезней растений, признаки поражения SCN на надземной части культуры проявляются только спустя несколько недель после заражения, что затрудняет раннюю диагностику и своевременные меры борьбы [2]. Эта особенность способствует быстрому распространению нематоды и значительным потерям урожая, что подчеркивает необходимость разработки эффективных методов контроля и устойчивых сортов сои [3].
В основе селекции устойчивых сортов сои лежит понимание молекулярно-генетических механизмов защиты растения. На сегодняшний день ключевую роль в резистентности к соевой цистообразующей нематоде (SCN) Heterodera glycines играют два основных гена: Rhg1 (хромосома 18) и Rhg4 (хромосома 8) [4]. Эти гены формируют многоуровневую систему защиты, основанную на комбинации различных молекулярных механизмов.
Ген Rhg1 кодирует LRR-рецептор, распознающий эффекторы нематоды и запускающий программируемую клеточную смерть растений через гиперчувствительный ответ, что приводит к некрозу и изоляции зоны питания паразита [5]. Параллельно этому ген Rhg4 кодирует серпин – белок из консервативного надсемейства сериновых ингибиторов протеаз. Его природная функция заключается в ингибировании (подавлении активности) пищеварительных сериновых протеаз нематоды. Это приводит к нарушению процессов питания и развития личинок, формируя один из молекулярных барьеров устойчивости растения [5].
Совместное действие двух механизмов – распознавания патогена через LRR-рецептор и ингибирования пищеварения через серпин – обеспечивает устойчивый резистентный фенотип растения.
На молекулярном уровне это взаимодействие приводит к распознаванию эффекторов нематоды и нарушению ее метаболизма, на клеточном уровне – к программируемой гибели клеток и нарушению питания личинок. В конечном итоге это проявляется в фенотипе как изоляция паразита, значительное снижение репродуктивного потенциала нематоды и уменьшение количества цист в почве, что обеспечивает устойчивость растений и сохранение урожайности сои.
В селекции сои на устойчивость к SCN исторически доминируют два основных генетических источника, определяющие два фундаментальных типа резистентности. Наиболее распространенным является тип PI 88788, на основе которого создано более 95 % коммерческих сортов [6]. Его устойчивость контролируется преимущественно многокопийным вариантом локуса Rhg1-b ; высокое число копий генов в этом тандемном повторе обеспечивает количественный защитный эффект за счет повышенной экспрессии. Второй, более редкий и ценный источник – тип Peking (PI 548402), к которому относятся изученные в данной работе сорта. Его устойчивость является результатом синергического действия двух генов: низкокопийного аллеля Rhg1-a и гена Rhg4 , кодирующего сериновую протеазу (SHMT). Механизм устойчивости Peking-типа основан на качественном взаимодействии этих двух генетических компонентов, что часто обеспечивает более широкий спектр защиты и считается стратегически важным для преодоления расовой адаптации патогена, развившейся в ответ на повсеместное использование сортов типа PI 88788 [7].
Наиболее оптимальным методом генотипирования является ARMS-ПЦР (Amplification Refractory Mutation System PCR) [6]. Система ПЦР с рефрактерными мутациями (ARMS) представляет собой простой и эффективный метод обнаружения однонуклеотидных полиморфизмов, включающих изменения одного нуклеотида или небольшие делеции. Технология ARMS-ПЦР широко применяется для генотипирования однонуклеотидных полиморфизмов (SNP) с помощью рефрактерных прай- меров. Способность праймеров присоединяться и инициировать амплификацию зависит от наличия соответствующего аллеля: мутантный праймер рефрактерен или устойчив к обычной ПЦР и наоборот. В результате получается система, которая позволяет точно определить наличие или отсутствие конкретного SNP, основываясь на способности праймеров присоединяться и инициировать реакцию [8].
Целью данного исследования является молекулярно-генетическая оценка устойчивости сортов сои селекции ВНИИ сои к цистообразующей нематоде с использованием метода ARMS-ПЦР, для чего были поставлены следующие задачи: провести молекулярно-генетическое генотипирование четырех перспективных сортов амурской сои (Сентябринка, Куханна, Нега 1, ВНИИС 1), оценить аллельное состояние ключевых генов устойчивости Rhg1 и Rhg4 , а также определить генетические профили устойчивости сортов и провести их классификацию согласно известным типам резистентности (Peking-тип).
Материалы и методы. Молекулярногенетический анализ был выполнен в лаборатории биотехнологии ФНЦ ВНИИ сои в 2025 г. В качестве объектов исследования использовались четыре сорта сои ( Glycine max (L.) Merr.) амурской селекции: Сентябринка, ВНИИС 1, Нега 1,
Куханна. Для каждого сорта было отобрано и проанализировано по 25 семян. Для выделения ДНК использовали молодые листья проростков сои (100–150 мг). Листья отбирали на стадии первого тройчатого листа. Перед этим семена проращивали в лабораторных условиях по методу бумажных рулонов: семена размещали между слоями увлажненной фильтровальной бумаги, свернутой в рулон.
Для анализа были отобраны праймеры к локусам резистентности сои к нематоде (табл. 1) [9]. Ввиду сложности использования KASP-анализа, описываемого в первоисточнике, а также недоступности заказа FRET-кассет для выполнения данного анализа, было принято решение об укорочении изначальных олигонуклеотидов и адаптации данной методики к ARMS-ПЦР.
В ходе исследования были протестированы различные температуры отжига праймеров для обеспечения специфической амплификации. Наилучшие результаты получены при температуре отжига 55 °C, при которой все пять пар праймеров эффективно гибридизовались с целевыми последовательностями. Электрофоретический анализ продуктов ПЦР подтвердил успешную амплификацию, визуализировав четкие полосы ожидаемого размера, что свидетельствует о высокой специфичности и воспроизводимости проведенных реакций.
Таблица 1
Адаптированные для ARMS праймеры к локусам резистентности к соевой цистообразующей нематоде
Table 1
Primers for soybean cyst nematode resistance loci adapted for ARMS
|
Локус |
Маркер |
Праймеры |
Длина |
Источник |
|
Rhg1 |
GSM381 |
1f: GCCAAAGAACTTGAG S AG B ATGAG (устойчивый аллель) 2f: GCCAAAGAACTTGAG S AG B ATGAT (восприимчивый аллель) |
77 |
Shi et al., 2015 |
|
r: CAAACAATAGGTCCAACCACCA |
||||
|
Rhg1 |
GSM383 |
1f: ATCTGC M AACTCTGTAAAGAGGAC (определяет тип устойчивости Peking-тип) 2f: ATCTGC M AACTCTGTAAAGAGGAG (определяет тип устойчивости PI 88788) |
102 |
|
|
r: GCTGTCCAGTCTCCAGAAGTGAA |
||||
|
Rhg4 |
GSM191 |
1f: CATCATGGGGCTAGATCTCCC (устойчивый аллель) 2f: CATCATGGGGCTAGATCTCCG (восприимчивый аллель) |
132 |
|
|
r: TAGCCGGTGGTGGAGTTTACC |
*Примечание: B – не A (то есть C, G, T или U); M – A, C; S – C, G
Реакцию полимеразной цепной реакции проводили в термоциклере CFX-96 (“Bio Rad”, США) по следующему протоколу: начальная денатурация при 94 °C в течение 5 мин; затем 35 циклов, каждый из которых включал денатурацию при 94 °C в течение 30 с, отжиг праймеров при 55 °C в течение 30 с и элонгацию при 72 °C в течение 45 с; завершающая элонгация при 72 °C в течение 5 мин. Для обеспечения достоверности результатов амплификацию для каждого образца проводили в двух независимых технических повторностях. Визуализацию выполняли в гель-документирующей системе GelDoc Ez Imager (“Bio Rad”, США). Для расчета длин полученных продуктов использовался маркер длин 100+bp («Евроген», Россия), с детекцией и расчетом длин в программе Image Lab Software.
Для количественной оценки генетической структуры сортов проведён статистический анализ данных генотипирования 25 индивидуальных растений каждого сорта. Рассчитаны доли (%) и 95%-ные доверительные интервалы для частот встречаемости генотипов. Статистическую значимость отклонения наблюдаемого распределения генотипов от гипотетических моделей (модель генетически однородного сорта и модель менделевского расщепления) оценивали с использованием критерия согласия χ² (хи-квадрат) Пирсона. Все расчёты выполнены на языке Python (версия 3.9) с применением библиотек SciPy (v1.7.3) для проведения статистических тестов и Pandas (v1.3.5) для обработки данных.
Результаты и обсуждение. Локус Rhg1, расположенный на хромосоме 18 (Chr18), является центральным компонентом устойчивости ко всем известным расам SCN. Ген кодирует белок, содержащий лейцин-бога-тые повторы (LRR), который участвует в распознавании патогена и активации защитных реакций растения. В зависимости от генотипа сои Rhg1 может присутствовать в виде нескольких аллельных вариантов, которые определяют различную степень устойчивости и специфичность к отдельным расам нематоды [10]. В данной работе маркер GSM383 был использован для дифференциации аллельных вариантов локуса Rhg1, позволяя уточнить генетический профиль устойчивости, ассоциированный с Peking-типом и PI 88788. В результате амплификации сорта Сентябринка и ВНИИС 1 являются гетерозиготами (рис. A, B).
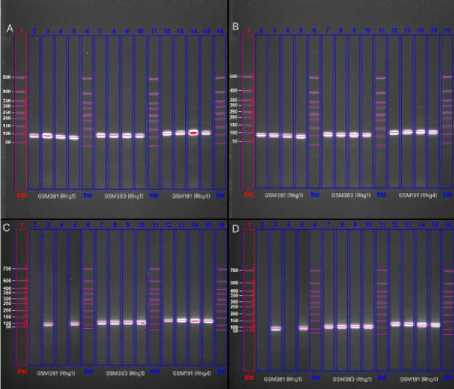
Рисунок – Результаты гель-электрофореза (представлены для визуализации целевых ампликонов):
А – электрофореграмма сорта Сентябринка; B – электрофореграмма сорта ВНИИС 1; C – электрофореграмма сорта Куханна; D – электрофореграмма сорта Нега 1;
дорожки: 1, 6, 11, 16 – маркер длин; 2, 4 – GSM381 (устойчивый аллель); 3, 5 – GSM381 (восприимчивый аллель); 7, 9 – GSM383 (Peking-аллель); 8, 10 – GSM383 (PI 88788-аллель); 12, 14 – GSM191 (устойчивый аллель); 13, 15 – GSM191 (восприимчивый аллель)
Fig. – Gel electrophoresis results (presented for visualization of target amplicons):
A – electrophoregram of the variety Sentyabrinka;
B – electrophoregram of the variety VNIIIS 1; C – electrophoregram of the variety Kukhanna; D – electrophoregram of the variety Nega 1;
lanes: 1, 6, 11, 16 – length marker; 2, 4 – GSM381 (resistant allele); 3, 5 – GSM381 (susceptible allele); 7, 9 – GSM383 (Peking allele); 8, 10 – GSM383 (PI 88788 allele); 12, 14 – GSM191 (resistant allele); 13, 15 – GSM191 (susceptible allele)
Гетерозиготность сортов Сентябринка и ВНИИС 1 по маркеру GSM381 (Rhg1), выявленная в результате амплифика- ции (рис. A, B), в сочетании с гетерозиготностью по локусу Rhg4 формирует генетическую архитектуру Peking-типа, которая ассоциируется с полной устойчивостью к соевой цистообразующей нематоде. В отличие от них, гомозиготность сортов Ку-ханна и Нега 1 (рис. C, D) по маркеру GSM381 указывает на наличие у них идентичных аллельных вариантов в обеих гомологичных хромосомах, причем, как показали результаты генотипирования, это гомозиготность по восприимчивому аллелю.
В общем случае гомозиготные генотипы обеспечивают консистентное наследование признака. Однако в данном исследовании гомозиготность сортов Нега 1 и Куханна по восприимчивому аллелю Rhg1 обусловила их стабильную восприимчивость, что и отражено в фенотипическом прогнозе (табл. 2).
Ключевым результатом исследования является выявление различного генетического базиса устойчивости у изученных сортов. Устойчивость сортов Сентябринка и ВНИИС 1 детерминирована гетерозиготным профилем (Peking-аллель и PI 88788-аллель) по локусам Rhg1 и Rhg4 (рисунок), что ассоциировано с умеренной устойчи- востью к SCN. В отличие от них восприимчивость сортов Нега 1 и Куханна однозначно обусловлена гомозиготностью по рецессивному (восприимчивому) аллелю локуса Rhg1 (рис. C, D), что нивелирует положительный эффект от гетерозиготности по Rhg4. Полученные данные чётко дифференцируют сорта по потенциалу устойчивости (табл. 2) и предоставляют селекционерам молекулярную основу для целенаправленного отбора.
Гетерозиготность по маркеру GSM383 указывает на наличие в генотипе растения различных аллельных вариантов в пределах локуса Rhg1 , которые ассоциированы с известными источниками устойчивости. В контексте селекционной популяции это может отражать процесс гибридизации и рекомбинации между линиями, несущими разные типы устойчивости.
Согласно данным, у сортов Сентяб-ринка и ВНИИС 1 наблюдается гетерозиготность по всем трём маркерам (табл. 2). В то же время у сортов Нега 1 и Куханна гомозиготность по маркеру GSM381 ( Rhg1 , восприимчивый аллель) сочетается с гетерозиготностью по остальным локусам, что определяет их восприимчивость к нематоде.
Таблица 2
Аллельное состояние генов у сортов сои по исследуемым маркерам
Table 2
Allele status of genes in soybean varieties according to the studied markers
|
Сорт |
GSM381 ( Rhg1 ) |
GSM383 (тип устойчивости) |
GSM191 ( Rhg4 ) |
Прогноз устойчивости |
|
Сентябринка |
Гетерозигота |
Гетерозигота |
Гетерозигота |
Умеренно устойчива в текущем поколении. Требует отбора гомозигот в потомстве (Peking-аллель и PI 88788-аллель) |
|
ВНИИС 1 |
Гетерозигота |
Гетерозигота |
Гетерозигота |
Умеренно устойчива в текущем поколении. Требует отбора гомозигот в потомстве (Peking-аллель и PI 8878-аллель) |
|
Нега 1 |
Гомозигота (восприимчивый аллель) |
Гетерозигота |
Гетерозигота |
Восприимчив |
|
Куханна |
Гомозигота (восприимчивый аллель) |
Гетерозигота |
Гетерозигота |
Восприимчив |
В отличие от них сорта Нега 1 (рис. C) и Куханна (рис. D) являются гомозиготными по восприимчивому аллелю локуса Rhg1 , что, несмотря на гетерозиготность по Rhg4 , определяет их статус как восприимчивых к соевой цистообразующей нематоде. При этом все четыре сорта характеризуются идентичным гетерозиготным профилем по маркеру GSM383 , что подтверждает единый механизм распознавания патогена на данном этапе защиты [11–14].
Статистический анализ распределения генотипов по ключевому локусу Rhg1 (маркер GSM381 ) выявил чёткое различие между изучаемыми сортами (табл. 3).
Таблица 3
Оценка генетической структуры сортов
Table 3
|
Assessment o Сорт |
f the genetic str Доля гетерозигот по GSM381 ( Rhg1 ), % |
ucture of varieties Статистическая проверка (критерий χ²) |
|
Сентябринка |
92,0 |
χ² = 21.9, p < 0,001 |
|
ВНИИС 1 |
96,0 |
χ² = 24,0, p < 0,001 |
|
Нега 1 |
0,0 |
χ² ≈ 0,0, p ≈ 1,0 |
|
Куханна |
0,0 |
χ² ≈ 0,0, p ≈ 1,0 |
Сорта Сентябринка и ВНИИС 1 формируют генетически гетерогенную группу с преобладанием гетерозиготного профиля (92–96%). Высокозначимые значения критерия χ² (p < 0,001) подтверждают, что их генетическая структура статистически достоверно отличается от модели чистой линии, что характерно для селекционных популяций на этапе отбора.
Напротив, сорта Нега 1 и Куханна продемонстрировали абсолютную (100%) гомозиготность по восприимчивому аллелю. Статистический анализ (p ≈ 1,0) не выявил отклонений от модели генетически однородной восприимчивой линии, что однозначно подтверждает их фенотипическую предрасположенность.
Генотипирование по маркерам GSM383
(определяющий тип устойчивости) и GSM191 (локус Rhg4 ) показало единообразие: все 100 проанализированных растений обладали гетерозиготным профилем, что свидетельствует о стабильной фиксации данных аллелей в генетическом материале.
Выявленный высокий уровень гетерозиготности (92–96%) по локусу Rhg1 у сортов Сентябринка и ВНИИС 1, формально являющихся чистыми линиями, представляет собой интересный феномен. Такое распределение генотипов статистически достоверно отличается от ожидаемого для гомогенного сорта (p < 0,001). Объяснение этого факта – будь то результат целенаправленного отбора на гетерозиготность, механизм генетического дрейфа или артефакт, связанный с ограниченной выборкой, – требует отдельного углубленного изучения.
Полученные молекулярно-генетические данные обладают значительной прогностической силой и могут служить эффективным инструментом для раннего отбора устойчивых к соевой цистообразующей нематоде генотипов. Проведенное высокоспецифичное генотипирование с использованием усовершенствованной методики ARMS-ПЦР создает научную основу для селекционной работы, позволяя на ранних этапах исключать восприимчивые формы (Нега 1, Куханна) и концентрировать усилия на линиях с определенным профилем устойчивости Peking-типа (Сен-тябринка, ВНИИС 1). Для подтверждения эффективности предложенного подхода и оценки уровня устойчивости в конкретных почвенно-климатических условиях Дальневосточного региона планируется проведение полевых испытаний с искусственным заражением и естественной ин-фестацией. Перспективы дальнейших исследований связаны с применением омиксных технологий для углубленного изучения молекулярных механизмов устойчивости, выявления новых паттернов взаимодействия «растение–патоген» и разработки расширенных панелей молекулярных маркеров для точной селекции [15].
Заключение. Проведенное молекулярно-генетическое исследование выявило чёткую дифференциацию изучаемых сортов сои по устойчивости к цистообразующей нематоде: сорта Сентябринка и ВНИИС 1 характеризуются умеренной устойчивостью (Peking-аллель и PI 88788-аллель), обусловленной гетерозиготностью по ключевым локусам Rhg1 и Rhg4 , в то время как Нега 1 и Куханна оказались восприимчивыми вследствие гомозиготности по рецессивному (восприимчивому) аллелю Rhg1 . Результаты подтвердили эффективность ARMS-ПЦР для генотипирования и подчёркивают важность селекции на основе гетерозиготного состояния генов Rhg1 и Rhg4 для создания устойчивых сортов сои, адаптированных к условиям Дальневосточного региона.
