Оценка вклада анемофильного переноса пыльцы в генетическое засорение сортов масличного льна

Автор: Зеленцов С.В., Олейник В.И., Рябенко Л.Г., Овчарова Л.Р., Зеленцов В.С.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (178), 2019 года.
Бесплатный доступ
У долгунцовых и масличных форм льна обыкновенного возможен энтомофильный и анемофильный перенос пыльцы. Однако доля вклада анемофильного переопыления в генетическое загрязнение недостаточно изучена. Для определения вероятности анемофильного переноса пыльцы между соседними сортами масличного льна, в 2017 г. на базе ФГБНУ ФНЦ ВНИИМК был заложен специальный полевой эксперимент. Коричневосемянный сорт масличного льна ВНИИМК 620 был обсеян желтосемянным сортом ВНИИМК 630. После созревания растений по восьми азимутам горизонта в сорте ВНИИМК 630 отобраны образцы растений. Полученные из них семена F1 были высеяны на следующий год. В потомствах F2 подсчитывали коричневые семена. Средняя частота встречаемости коричневых семян в F2 в желтосемянном сорте ВНИИМК 630 в 2018 г. составила 0,097 %. Этот уровень генетического засорения представляет угрозу для сохранения сортовой чистоты семян высших репродукций семян льна. Заметных закономерностей в изменении частот выщепления коричневых семян по азимутам горизонта и направлений ветров обнаружено не было...
Масличный лён, генетическое засорение, анемофильное переопыление, ветровой перенос пыльцы, роза ветров, румбы горизонта
Короткий адрес: https://sciup.org/142220378
IDR: 142220378 | УДК: 633.854.54:631.527:631.52 | DOI: 10.25230/2412-608X-2019-2-178-3-8
Assessment of the contribution of ane-mophilic pollen transfer to genetic contamination of oil flax varieties
In fiber and oil forms of common flax, entomophilous and anemophilic pollen transfer is possible. However, the share of the contribution of anemophilic cross-pollination to genetic contamination is not well understood. To determine the probability of anemophilic pollen transfer between adjacent varieties of oil flax in 2017 a special field experiment was established on the basis of the by V.S. Pustovoit All-Russian Research Institute of Oil Crops, Krasnodar. The brown-seed oil flax variety VNIIMK 620 was around sown with the yellow-seed variety VNIIMK 630. After the plants ripened, along eight azimuths of the horizon in the variety VNIIMK 630 samples of plants were taken. The F1 seeds obtained from them were sown the following year. In the F2 progeny, appeared brown seeds were counted. The average frequency of occurrence of brown seeds in F2 in the yellow-seed variety VNIIMK 630 in 2018 was 0.097 %. This level of genetic contamination poses a threat to preserve the varietal purity of the breeders and foundation seeds of flax...
Текст научной статьи Оценка вклада анемофильного переноса пыльцы в генетическое засорение сортов масличного льна
Введение. Лён обыкновенный ( Linum usitatissimum L.), включая его долгунцовые и масличные формы, традиционно относится к растениям-самоопылителям. Однако в ряде работ у этого вида фиксировалось незначительное (до 1 %) перекрёстное опыление [2; 6; 7; 13; 14]. По мнению большинства авторов, основной причиной появления генетических примесей (без учёта механического засорения) может быть энтомофильное переопыление [3; 7; 8; 11; 13; 14]. В разные годы ряд крупных исследователей льна, например, Е.В. Эллади (1940), И.А. Сизов (1955), A. Diederishsen (2003), полагали, что культурный лён привлекает насекомых-опылителей из целого ряда семейств благодаря крупным и яркоо-крашенным цветкам с легкодоступной пыльцой и нектарниками [9; 11; 12].
Ещё одним механизмом перекрёстного опыления у льна обыкновенного может быть анемофильный (ветровой) перенос некрупной пыльцы (50–68 мкм) [3; 7; 11; 13; 14]. В отдельных исследованиях, например, О.И. Рыжеевой (1967), частота анемофильного переопыления льна достигала 4,0–12,2 % [8].
Таким образом, ретроспективный литературный анализ свидетельствует о возможности энтомофильного и анемофильного переноса пыльцы у льна обыкновенного. Исходя из требований ГОСТ Р 52325–2005 «Сортовые и посевные качества семян» к семенам масличного льна, 4
сортовая чистота репродукций ОС и ЭС должна быть не ниже 99,6 %; репродукции РС – не ниже 98,0 %; репродукции РСт – не ниже 97,0 % [5]. Поэтому многочисленные факты, пусть незначительного, но переопыления льна, могут являться одной из причин снижения сортовой чистоты семенных партий семян, в том числе высоких репродукций.
Ранее, на примере ряда масличных сортов льна обыкновенного, нами были изучены биологические особенности цветения, а также размеры отдельных пыльцевых зёрен и их агломератов, подтверждающие теоретическую возможность анемофильного переноса пыльцы [6]. Однако доля возможного вклада анемофильного переопыления в общее генетическое загрязнение сортов льна осталась неизученной, что и послужило целью настоящих исследований.
Материалы и методы . Исследования проводили в 2017–2018 гг. на центральной базе ФГБНУ ФНЦ ВНИИМК, г. Краснодар. Для определения вероятности анемофильного переноса пыльцы между сортами масличного льна в 2017 г. был заложен специальный полевой эксперимент.
Сорт масличного льна ВНИИМК 620 с доминантными признаками коричневой окраски семян и голубой окраски венчика цветка был высеян на квадратном участке площадью 100 м2, ориентированном сторонами по основным направлениям горизонта (С, В, Ю и З). Сразу после завершения посева сорт ВНИИМК 620 со всех четырёх сторон был обсеян сортом ВНИИМК 630 с рецессивными признаками жёлтой окраски семян и белой окраски венчика цветка (рис. 1).
Ширина полосы обсева сорта ВНИИМК 620 сортом ВНИИМК 630 – 20 м. Перед созреванием растений на экспериментальном участке в сорте ВНИИМК 630 по компасу были определены и отмечены вешками восемь географических азимутов (С, СВ, В, ЮВ, Ю, ЮЗ, З, СЗ) для отбора сноповых образцов (рис. 2).

Рисунок 1 – Внешний вид экспериментального участка в фазе цветения сортов ВНИИМК 620 с голубой окраской цветов (в центре) и ВНИИМК 630 с белой окраской цветов, 2017 г.
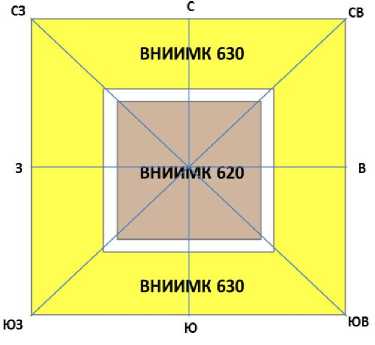
Рисунок 2 – Схема и конфигурация экспериментального участка с направлениями отборов сноповых образцов по 8-ми азимутам горизонта
В фазе созревания желтосемянного сорта ВНИИМК 630, по всем восьми азимутам, с интервалом в 2,5 м были произведены сноповые отборы с площади 1 м2 каждый. Полученные при ручном обмолоте всех отобранных снопов семена, во избежание механического засорения, вручную были высеяны в 2018 г. на отдельных делянках. После созревания по-томств F1 все делянки индивидуально, вручную были убраны и обмолочены. В полученных образцах желтоокрашенных семян подсчитывали (при наличии) количество коричневых семян, как генетиче- ских примесей вследствие переноса пыльцы от коричневосемянного сорта ВНИИМК 620.
Данные о направлениях ветра в период цветения льна – с 25.05.2017 по 25.06.2017 – получали из архива метеонаблюдений метеостанции «Круглик», расположенной на расстоянии 5600 м на юго-запад от экспериментального участка [1]. Статистическую обработку полученных результатов проводили с использованием алгоритмов проверки отклонения распределения экспериментальных выборок от нормального на основе критерия нормальности ( W ) Шапиро-Уилка [4], прямолинейной корреляции ( r ) Пирсона и непараметрической ранговой корреляции ( ρ ) Спирмена [10].
Результаты и обсуждение . Период цветения сортов льна ВНИИМК 620 и ВНИИМК 630 в 2017 г. происходил с 30 мая по 25 июня. В том числе раскрытие цветков на цветоносных осях 1-го и 2-го порядков (основное цветение) отмечали с 30 мая по 14 июня; цветение цветоносных осей 3-го и 4-го порядков – с 15 по 25 июня. В условиях центральной зоны Краснодарского края ветровой перенос подсохшей пыльцы после растрескивания пыльников у льна возможен примерно с 9:00 ч до 12:00 ч [6].
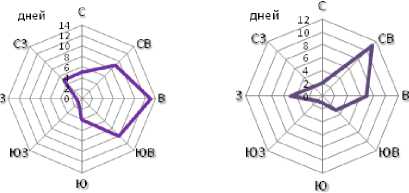
На основе метеоданных метеостанции «Круглик» [1] в период цветения льна в 2017 г. с 25 мая по 25 июня были построены розы направления ветров, дувших с 9:00 ч до 12:00 ч, и потенциального анемофильного переноса пыльцы, отдельно в период основного цветения – с 30 мая по 14 июня, и в период вторичного цветения боковых цветоносов – с 15 по 25 июня (рис. 3).
Жёлтая окраска семян у льна является моногенно наследуемым рецессивным признаком [2]. Поэтому в случае горизонтального переноса пыльцы с цветущих растений сорта ВНИИМК 620 на цветущие растения сорта ВНИИМК 630 в потомстве F2 желтосемянного сорта следует ожидать выщепления коричневых семян. Если основным вектором переноса пыль- цы в эксперименте был ветер, то наибольшее количество коричневых семян будет обнаруживаться в потомствах проб, отобранных по азимутам, соответствующим основным направлениям ветров в период цветения.
а б
Рисунок 3 – Роза направлений ветров и потенциального анемофильного переноса пыльцы с 9:00 до 12:00 ч в период с 30.05.2017 г. по 25.06.2017 г.
а – в период с 30.05 по 14.06;
б – в период с 15.06 по 25.06
Исследования частот выщепления коричневых семян (%) в потомствах F 2 желтосемянного сорта ВНИИМК 630, отобранных по восьми азимутам горизонта на дистанциях от 2,5 до 12,5 м от сорта ВНИИМК 620, показали, что наибольшая частота выщепления коричневых семян была выявлена в северо-восточном, а также в северном и юго-восточном направлениях (рис. 4).
Однако в целом каких-либо выраженных закономерностей в изменении частот выщепления коричневых семян по азимутам горизонта и направлений ветров обнаружено не было.
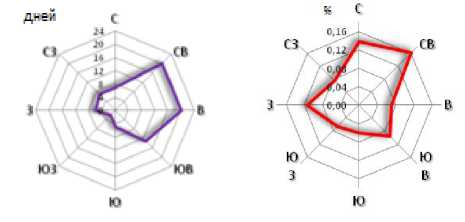
В целом за период с 30.05.2017 по 25.06.2017 г. преимущественными направлениями ветров были северовосточный, восточный и юго-восточный азимуты. Максимальные частоты выщеп-ления коричневых семян в потомствах F 2 желтосемянного сорта ВНИИМК 630, отобранных на дистанциях от 2,5 до 12,5 м от коричневосемянного сорта ВНИИМК 620, выявлены в северо-восточном и северном направ-лении, несколько меньше – в юго-6
восточ-ном и западном направлениях (рис. 5).
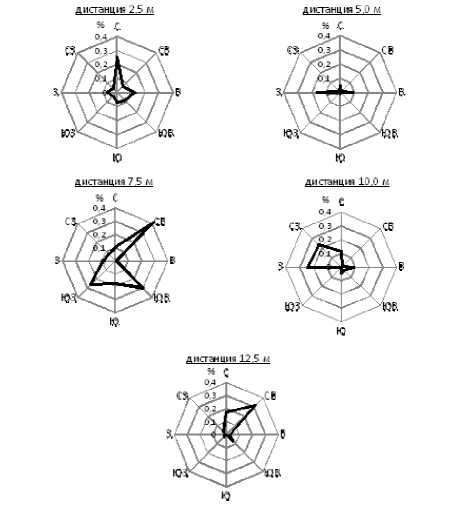
Рисунок 4 – Частоты выщепления коричневых семян (%) в потомствах F 2 желтосемянного сорта ВНИИМК 630, отобранных по 8 азимутам горизонта на дистанциях от 2,5 до 12,5 м от сорта ВНИИМК 620, 2018 г.
б
а
Рисунок 5 – Сравнительная оценка изменчивости частоты встречаемости коричневых семян в потомствах F 2 желтосемянного сорта ВНИИМК 630 от направления ветра: а – общая роза направлений ветров в период с 30 мая по 25 июня 2017 г. с 9:00 ч по 12:00 ч; б – средняя частота обнаружения коричневых семян в 2018 г. в потомствах F 2 сорта ВНИИМК 630
Совпадения между направлением ветра и увеличенной частотой выщепления коричневых семян F 2 выявлены только по северо-восточному азимуту, и, частично, по юго-восточному.
В целом, средняя из 40 точек отбора в массиве сорта ВНИИМК 630 частота встречаемости коричневых семян в F 2 в 2018 г. составила x̅ = 0,097 % при варьировании этого показателя от 0 до 0,41 %.
Для выбора алгоритма корреляционного анализа оценили степень отклонения экспериментальной выборки данных от нормального распределения с использованием критерия нормальности W Шапиро-Уилка. В экспериментальной выборке этот критерий составил W факт. = 0,899 при W табл. = 0,605. Поскольку W факт. > W табл. , следовательно, экспериментальная выборка данных имеет близкое к нормальному распределение и к ней применимы параметрические методы статистики. Однако расчётный критерий W факт. имеет значимость только при р- уровне = 0,281 (вероятность ошибки = 28,1 %), что крайне недостаточно по сравнению с принятым в биологической статистике уровнем значимости р -уровня = 0,05 (5%-ный уровень значимости или 5%-ная вероятность ошибки). Поэтому для установления корреляционных связей между частотами выщепления коричневых семян и розой направлений ветров, помимо прямолинейной корреляции Пирсона, дополнительно использовали не требующий нормализации выборки алгоритм непараметрической ранговой корреляции Спирмена.
Параметрический коэффициент корреляции Пирсона между направлениями ветров в 2017 г. и частотой выщепления коричневых семян в 2018 г., составил r = 0,380, что при р -уровне = 0,353 и коэффициенте детерминации r 2 = 0,144 указывает на отсутствие тесных корреляционных связей между ними.
Непараметрический коэффициент корреляции Спирмена между этими же показателями оказался несколько выше и составил ρ = 0,530 при р-уровне = 0,177 (17,7%-ная вероятность ошибки) и коэффициенте детерминации ρ2 = 0,281. Тем не менее непараметрическая статистика Спирмена также свидетельствует о фактическом отсутствии значимой корреляции между розой направлений ветров и частотами выщепления генетических примесей.
Выводы . Проведённые исследования показывают, что гипотеза об анемофильном (ветровом) переносе пыльцы, как основной причине выщепления коричневых семян в желтосемянном сорте масличного льна ВНИИМК 630, отвергается.
На основании метода исключения можно утверждать, что основной причиной генетического засорения льна является энтомофильное переопыление, зависящее преимущественно от частоты посещений цветков льна разными видами пчёл и шмелей, и, возможно, некоторыми другими видами насекомых.
Доля генетических примесей с признаком коричневой окраски семян масличного льна на примере сорта ВНИИМК 630 в условиях 2017 г. составила 0,097 % при варьировании этого показателя от 0 до 0,41 %. Поэтому выявленный в наших экспериментах уровень энтомофильного генетического засорения льна представляет угрозу для поддержания высокого уровня сортовой чистоты семян высших репродукций семян.
В связи с этим при выращивании оригинальных и элитных семян рекомендуется территориально разделять между собой посевы различных сортов льна на расстояние не менее половины дневных перелётов основных насекомых-опылителей.
Список литературы Оценка вклада анемофильного переноса пыльцы в генетическое засорение сортов масличного льна
- Архив погоды в Краснодаре/Круглике за 25.05.2017-25.06.2017 -. -URL: https://rp5.ru/Архив_погоды_в_Краснодаре,_Круглике (дата обращения: 31.03.2019).
- Брач Н.Б. Внутривидовое разнообразие льна (Linum usitatissimum L.) и его использование в генетических исследованиях и селекции: дис. … д-ра биол. наук. -Санкт-Петербург: Всероссийский научно-исследовательский институт растениеводства им. Н.И. Вавилова, 2007. -С. 21-22, 76.
- Галкин Ф.М. Лён масличный: селекция, семеноводство, технология возделывания и уборки/Под ред. Н.И. Бочкарёва. -Краснодар, 2008. -С. 21-26.
- ГОСТ Р ИСО 5479-2002 Статистические методы. Проверка отклонения распределения вероятностей от нормального распределения. -Введ. 2002.01.22, № 25-ст. -М.: ИПК «Изд-во стандартов», 2002. -30 с.
- ГОСТ Р 52325-2005 Семена сельскохозяйственных растений. Сортовые и посевные качества семян. Общие технические условия. -Введ. 2005.03.23, № 63-ст. -М.: Стандартинформ, 2005. -С. 8.
- Зеленцов С.В., Мошненко Е.В., Рябенко Л.Г., Овчарова Л.Р. Типы и способы естественного опыления льна обыкновенного Linum usitatissimum L.//Масличные культуры. Науч.-тех. бюл. ВНИИМК. -2018. -Вып. 1 (173). -С. 105-113.
- Минкевич И.А. Лён масличный в СССР. -Краснодар: Краевое книжн. изд-во, 1940. -С. 32.
- Рыжеева О.И. Лён масличный//В сб.: Руководство по селекции и семеноводству масличных культур/Под общ. ред. В.С. Пустовойта. -М.: Колос, 1967. -С. 122-130.
- Сизов И.А. Лен. -М.: Сельхозгиз, 1955. -С. 97-101.
- Урбах В.Ю. Биометрические методы. -М.: Наука, 1964. -С. 282-286, 345-350.
- Эллади Е.В. Linum usitatissimum (L.) consp. nov. -Лён//В сб.: Культурная флора СССР. Прядильные/Под ред. Е.В. Вульфа. -М.-Л.: Гос. изд. колх. и совхозн. лит-ры, 1940. -Т. V. -С. 109-207.
- Diederishsen A., Richards R. Cultivated flax and the genus Linum L. Taxonomy and germplasm conservation//In: Flax. The genus Linum/Ed. by A.D. Muir and N.D. Westcott. -London, Taylor & Francis Ltd, 2003. -P. 32-42.
- Jhala A.J., Hall L.M., Hall J.C. Potential Hybridization of Flax with Weedy and Wild Relatives: An Avenue for Movement of Engineered Genes?//Crop Sci. -2008. -Vol. 48. -P. 825-840.
- Williams I.H. The pollination of linseed and flax//Bee World. -1988. -Vol. 69. -P. 145-152.
