Оценка вклада различных популяций в генетическое разнообразие свиней корня крупной белой породы

Автор: Зиновьева Н.А., Харзинова В.Р., Сизарева Е.И., Гладырь Е.А., Костюнина О.В., Луговой С.И., Тапиха В.А., Гамко Л.Н., Овсеенко Е.В., Шавырина К.М., Эрнст Л.К.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетическое разнообразие, цитогенетика, вирусные болезни
Статья в выпуске: 6 т.47, 2012 года.
Бесплатный доступ
С использованием 11 микросателлитов (S0155, S0355, S0386, SW72, SW951, S0101, SW240, SW857, S0228, SW911 и SW936) получена характеристика аллелофонда свиней пород крупная белая и йоркшир. Изучаемая выборка была представлена 17 популяциями животных российской, венгерской, датской, канадской, литовской, немецкой, украинской и французской селекции ( n = 837). Обработку данных проводили с помощью программного обеспечения GenAlEx (v. 6.4), Structure (v. 2.3.1) и Past (v. 2.15). Установлено, что 24,2 % общей изменчивости обусловлено межпопуляционными различиями (R st= 0,242 при p = 0,01). На основании значений индексов F st и R st (AMOVA) выявлен наибольший вклад в изменчивость географически удаленных и локальных популяций. Кластерный анализ, проведенный по J.K. Pritchard (2000) и M. Nei (1983), показал, что происхождение популяций отражается в формировании кластеров.
Свиньи, крупная белая порода, йоркшир, генетическое разнообразие, микросателлиты
Короткий адрес: https://sciup.org/142133358
IDR: 142133358 | УДК: 636.42:636.082:577.2:575.174
Evaluation of the contribution of different pig populations to the genetic diversity of the Large White breed
The characteristic of the allele pool of the Large White and Yorkshire pig breeds using 11 microsatellites (S0155, S0355, S0386, SW72, SW951, S0101, SW240, SW857, S0228, SW911 and SW936) was done. The study was performed on 17 pig populations of Russian, Danish, Hungarian, Canadian, Lithuanian, German, Ukrainian and French origins ( n = 837). The data were analyzed using GenAlEx (v. 6.4), Structure (v. 2.3.1) and Past (v. 2.15) software. It was shown that 24.2 per cent of the total microsatellite diversity was due to intra population differences (R st = 0.242, p = 0.01). Based on F stand R st (AMOVA) values the greater contribution of geographically distant and local populations into microsatellites diversity was observed. The cluster analysis performed according J.K. Pritchard (2000) and M. Nei (1983) showed that the cluster structure was in concordance with populations’ origin.
Текст научной статьи Оценка вклада различных популяций в генетическое разнообразие свиней корня крупной белой породы
Оценка спектра генетического разнообразия пород требует изучения по возможности большего числа генетически изолированных популяций. Существенный вклад в генетическое разнообразие пород вносят региональные популяции, генофонд которых, как правило, формировался в условиях относительной географической изоляции и на основе местного скота — носителя собственного уникального аллелофонда (1, 2). Кроме того, на аллельные профили пород, используемых в сельскохозяйственном производстве продукции животноводства, значительное влияние оказывают стратегии селекционно-племенной работы.
Наиболее распространенная порода свиней в мире — крупная белая. На ее долю в России приходится более 85 %, во Франции и Финляндии — около 63 %, в Польше — 60 %, в Англии — 50 % от общего поголовья (3). Порода выведена в Англии в графстве Йоркшир в середине XIX века. Первоначально она называлась йоркширской, а позже от Английского королевского земледельческого общества получила название крупной белой. В ряде стран, импортировавших английскую крупную белую породу, за ней закрепилось название йоркшир.
В Россию первые партии свиней английской крупной белой породы были завезены в конце 1870-х—начале 1880-х годов, второй завоз из Англии произошел в период с 1923 по 1931 год с целью восстановления численности поголовья, существенно снизившейся после Первой мировой войны (3). На формирование отечественной крупной белой породы значи- тельное влияние оказали щепкинские свиньи, которыми в то время были укомплектованы все племхозы. Так, около 30 % из 48 хряков и свыше 60 % из 340 маток, занесенных в I том племенной книги, вышедшей в 1932 году под редакцией Н.Н. Завадовского, имели кровь щепкинских свиней, несущих, в свою очередь, кровь павлиновских метисных свиней (4). В большинстве племенных хозяйств животных английской крупной белой породы использовали в скрещиваниях с местными метисными свиньями, хорошо приспособленными к природно-климатическим условиям региона (5). В 1920-1930-х годах крупную белую породу представляли животные как мясосального, так и беконного типа. Первый советский стандарт, разработанный в 1934 году Н.Н. Завадовским, предусматривал отбор свиней мясосального типа. В 1940-е годы был введен новый стандарт, предполагающий создание свиней густого сального типа, что было обусловлено потребностями в жирной свинине (6). Для совершенствования мясных и откормочных качеств в последующие годы применяли освежение крови с использованием крупных белых и йоркширских животных английской, шведской, голландской, датской, французской, канадской селекции. Импорт свиней из Англии проводился в 1947-1948 годах, из Швеции и Англии — в 1970-1981 годах, из Голландии — в 1982 году (7). В последнее десятилетие повышенный спрос на постную свинину стал причиной депопуляции свиней крупной белой породы отечественной селекции в ряде племенных хозяйств и их вытеснения животными зарубежной селекции — канадской (племенные фермы Канадского центра совершенствования свиней — Canadian Centre for Swine Improvement, CCSI), датской (племенные фермы, участвующие в программе DanBred, организованной датским исследовательским центром по свиноводству — Danish Pig Research Centre), ирландской («Hermitage Genetics»), голландской («Hypor B.V.», «Topigs»), французской («Cooperl Ark Atlantique», «France Hybrid»), немецкой («Rhg Hirshmann Hybrid GmbH») и др., обладающими высокими показателями мясной и откормочной продуктивности.
В качестве инструмента для определения состояния и динамики популяций у животных находят применение ДНК-маркеры разных типов, из которых к числу наиболее распространенных относятся микросателлиты (8). Показана их прикладная значимость для характеристики аллело-фонда пород, типов и линий животных, в контроле и управлении структурой стад (9-12).
Целью настоящей работы было изучение аллелофонда и оценка вклада отдельных популяций в генетическое разнообразие свиней крупной белой породы с использованием микросателлитов.
Методика . Исследовали образцы биологического материала (ушной выщип, консервант — 96 % этанол), взятого у 837 свиней пород крупная белая и йоркшир из 17 различных популяций.
Микросателлитный анализ проводили по 11 локусам (S0155, S0355, S0386, SW72, SW951, S0101, SW240, SW857, S0228, SW911 и SW936), используя разработанную тест-систему для ДНК-экспертизы свиней (13). Разделение и детекцию продуктов амплификации выполняли на генетическом анализаторе АВ13130xl («Applied Biosystems», США) согласно рекомендациям фирмы-изготовителя.
Статистическую обработку данных проводили по Б. Вейр (14) с использованием пакета Microsoft Excel 2007 с плагином GenAlEx v. 6.4. Определяли среднее число аллелей и число эффективных аллелей в расчете на локус, наблюдаемую и ожидаемую степени гетерозиготности, индекс фиксации Fis. Изменчивость микросателлитов в изучаемых популяциях свиней оценивали по значениям индекса фиксации Fst и Rst (рассчитан с использованием функции молекулярной вариансы — AM OVA). Для выполнения кластерного анализа по методу J.K. Pritchard с соавт. (15) применяли программное обеспечение Structure v. 2.3.1, при этом значения коэффициента подобия особи в популяции (Q) рассчитывали с помощью Admixture Model и Correlation Model без введения предварительной информации о популяционной принадлежности особей для числа популяций k, равного 2, 5, 10 и 17. Генетические дистанции вычисляли по M. Nei (16). Построение филогенетического дерева осуществляли методом попарного внутригруппового невзвешенного среднего (UPGMA, программного обеспечение Past v. 2.15).
Результаты . Характеристика проанализированных популяций и число исследованных образов приведены в таблице.
Происхождение групп свиней пород крупная белая (КБ) и йоркшир (Й) , ис
А
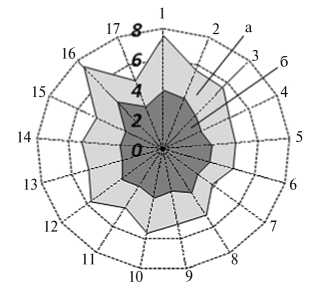
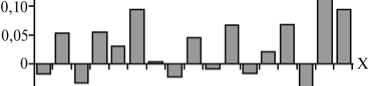
Рис. 1. Аллельные профили по 11 микросателлитам (А) и дефицит («+») /избыток (« - ») гетерозигот (Б) в изучаемых популяциях свиней пород крупная белая и йоркшир: оси Х — номер изучаемой популяции согласно таблице (А и Б); оси Y — число аллелей на локус микросателлитов (А, а — среднее число аллелей, б — среднее число эффективных аллелей; 0, 2, 4, 6 и 8 — шкала значений) и величина индекса фиксации Fis (Б).
0,25
0,20
0,15*
-0,05
-0,10-
I 3 5 7 9 II 13 15 17
пользуемых в исследованиях
|
№ популяции |
| Порода | Происхождение (регион) |
n |
|
|
1 |
КБ |
Селекция «Hypor B.V.» (Канада) |
100 |
|
2 |
Й |
Племенные фермы острова Принца Эдварда (Канада) |
15 |
|
3 |
Й |
Отечественная селекция с прилитием крови свиней европейской селекции (Республика Чувашия) |
43 |
|
4 |
КБ |
Селекция DanBred (Danish Pig Research Centre, Дания) |
26 |
|
5 |
КБ |
Отечественная селекция с прилитием крови свиней европейской селекции (Самарская обл.) |
24 |
|
6 |
Й |
Отечественная селекция с прилитием крови свиней европейской селекции (Самарская обл.) |
20 |
|
7 |
Й |
Фермы Canadian Centre for Swine Improvement (Канада) |
16 |
|
8 |
КБ |
Отечественная селекция с прилитием крови свиней европейской селекции (Белгородская обл.) |
52 |
|
9 |
КБ |
Селекция «Rhg Hirschmann Hybrid GmbH» (Германия) |
104 |
|
10 |
Й |
Отечественная селекция с прилитием крови свиней французской селекции «Cooperl Ark Atlantique» (Оренбургская обл.) |
37 |
|
11 |
КБ |
Селекция «Cooperl Ark Atlantique» (Франция) |
29 |
|
12 |
КБ |
Селекция «Hermitage Genetics» (Ирландия) |
61 |
|
13 |
КБ |
Отечественная селекция с прилитием крови свиней европейской селекции (Брянская обл.) |
78 |
|
14 |
КБ |
Отечественная селекция с прилитием крови свиней европейской селекции (Брянская обл.) |
80 |
|
15 |
Й |
Селекция «Norsvin Lietuva» (Литва) |
58 |
|
16 |
КБ |
Украинская селекция с прилитием крови свиней английской, датской и французской селекции (Украина, Запорожская обл.) |
72 |
|
17 Всего |
КБ |
Венгерская селекция (Венгрия) |
22 837 |
Б
Y
Среднее число аллелей на локус микросателлитов (рис. 1, А) варьировало от 3,8±0,5 в популяции № 7 до 7,5±0,5 в популяции № 1, число эффективных аллелей — от 2,6±0,2 в популяции № 4 до 4,3±0,6 в популяции № 16. Анализ величины индекса фиксации Fis (см. рис. 1, Б) выявил дефицит гетерозигот, превышающий 5 %, в семи из 17 изученных популяций (№№ 2, 4, 6, 11, 14, 16 и 17) при наибольшем дефиците в популяции № 16 (22,5 %). Избыток гетерозигот, превышающий 5 %, отмечали в популяции № 15 (8,6 %). В 15 из 17 исследованных групп (за исключением популяций № 2 и № 7) по ряду локусов наблюдалось высокодостоверное (р < 0,01 и р < 0,001) отклонение от генного равновесия по локусам микросателлитов. Наибольшее число локусов, в которых имелось отклонение, обнаружили в популяциях № 13 и № 16 (соответственно 6 и 7 локусов). Зависимость между характером (дефицит или избыток) гетерозиготности и смещением генного равновесия мы не выявили.
В 11 из 17 популяций были найдены приватные аллели, большинство из которых встречались с частотой менее 5 %. Наибольшее число таких аллелей (11 из 34) идентифицировалось в украинской популяции свиней (№ 16), что, по всей видимости, обусловлено отсутствием обмена генами с другими исследованными популяциями. В 4 популяциях (№№ 1, 2, 12 и 16) имелись приватные аллели с частотой > 5 % (их можно рассматривать как специфичные для этих популяций): № 1 — аллель 243 в локусе SO355 (5,6 %), № 2 — 244 в локусе SO228 (6,7 %), № 12 — 119 в локусе SW72 (5,1 %), № 16 — 97 в локусе SW72, 118 в локусе SW951, 87 в локусе SW240 и 93 в локусе SW936 (соответственно 8,7; 7,6; 5,5 и 6,3 %).
Анализ характера изменчивости аллелофонда микросателлитных локусов показал, что на долю межпопуляционных различий приходилось 24,2 % общей изменчивости (Rst = 0,242 при р = 0,01). Величина индекса фиксации Fst при парном сравнении, использованного в качестве критерия, свидетельствовала, что наибольший вклад в генетическую изменчивость всей выборки свиней вносили популяции канадской и французской селекции (для № 7 и № 11 значения Fst соответственно 0,134±0,007 и 0,123±0,006), а также региональные популяции из Брянской области (для № 13 и № 14 Fst соответственно 0,125±0,006 и 0,122±0,006) и Украины (для № 16 и № 17 Fst соответственно 0,115±0,005 и 0,119±0,005). Примененный в качестве критерия оценки индекс Rst (AMOVA), при расчете которого учитываются различия не только в частоте аллелей микросателлитов, но и в их длине, указывал на наибольший вклад популяции «Hypor B.V.» (№ 1, Rst = 0,338±0,054) в изменчивость изучаемой выборки. Следует отметить существенное влияние свиней датской селекции (популяция № 4, Rst = 0,269) на общую изменчивость. В остальном отмечались тенденции, аналогичные описанным выше: значения Rst для популяций №№ 7, 11, 13, 14 и 17 составили соответственно 0,281; 0,263; 0,251; 0,244 и 0,267, в то время как для других популяций индекс Rst варьировал от 0,139 до 0,223.
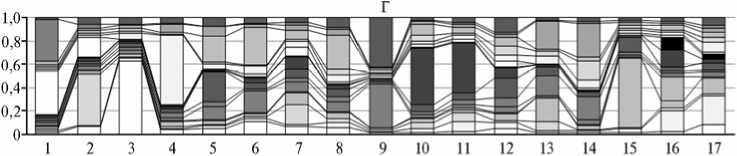
Результаты кластерного анализа изучаемых популяций свиней по J.K. Pritchard с соавт. (15) для числа кластеров k = 2, 5, 10 и 17 представлены на рисунке 2, данные филогенетического анализа по M. Nei (16) в виде филогенетического дерева обобщены на рисунке 3.
При k = 17 (см. рис. 2, Г), что соответствует числу изучаемых популяций, ни одна из них не формировала независимые кластеры при Q = 0,75. Это можно рассматривать как указание на единое происхождение групп свиней, обусловленное их принадлежностью к породам одного корня. Анализ кластерной структуры при k = 2 (см. рис. 2, А) выявил генетическую обособленность двух канадских популяций свиней от других исследованных групп: величина Q2/2 составляла соответственно 0,949 (№ 1) и 0,916 (№ 2). Относительно высокие значения, характеризующие членство в обсуждаемом кластере, имели популяции свиней с острова Принца Эдварда (Канада, популяция № 7), «Cooperl Ark Atlantique» (Франция, популяция № 10), а также отечественная и украинская популяции с прилитием крови от животных французской селекции (популяции № 11 и № 16): значения Q2/2 равнялись соответственно 0,452; 0,554; 0,642 и 0,771. На генеалогическом дереве (см. рис. 3) перечисленные популяции формировали общий подкластер. Распределение кластеров при k = 5 (см. рис. 2, Б) подтверждало иное происхождение популяции свиней «Hypor B.V.» (№ 1, Q2/5 = 0,858), что может быть следствием замкнутой системы их разведения в ряде генераций. Генетически эта популяция обособлена от 15 других, на что указывали

Номер популяции
Рис. 2. Кластерный анализ изучаемых популяций свиней пород крупная белая и йоркшир по J.K. Pritchard с соав. (15) по 11 микросателлитным локусам: А — k = 2, Б — k = 5, В — k = 10, Г — k = 7.
относительно невысокие показатели, отражающие их членство в кластере (Q2/5 = 0,021-0,174), и близка лишь одной популяции канадского происхождения (№ 2) (Q2/5 = 0,424), что, по-видимому, стало следствием географической общности указанных популяций. Такую же тенденцию прослеживали при k = 10 (см. рис. 2, В) и k = 17 (см. рис. 2, Г). При k = 10 для популяции № 1 и № 2 коэффициент Q4/10 составил соответственно 0,780 и 0,237, в то время как для других популяций значения, характеризующие членство в этом кластере, варьировали от 0,011 до 0,087. При k = 17 коэффициент Q11/17 составил соответственно 0,373 и 0,167, а у дру- гих значения для вышеназванных кластеров были существенно ниже — 0,010-0,078. Генетически обособленной оказалась популяция свиней «Rhg Hirshman Hybrid GmbH» (№ 9), о чем свидетельствовали высокие значения коэффициента подобия Q в соответствующем кластере при k = 5 (Q4/5 = 0,900), k = 10 (Q9/10 = 0,834) и k = 17 (Q17/17 = 0,424), тогда как для других популяций эти показатели варьировали соответственно от 0,027 до 0,411, от 0,020 до 0,234 и от 0,017 до 0,122. Относительно высокими коэффициентами Q в своих кластерах характеризовались популяции свиней из Республики Чувашия (№ 3, Q1/17 = 0,630), селекции «Norsvin Lietuva» (№ 15, Q4/17 = 0,594) и датского исследовательского центра по свиноводству (№ 4, Q12/17 = 0,592), для других изученных популяций значения Q составили соответственно 0,015-0,110; 0,017-0,202 и 0,014-0,088. Отмеченный факт можно расценивать как указание на генетическую обособленность названных популяций.
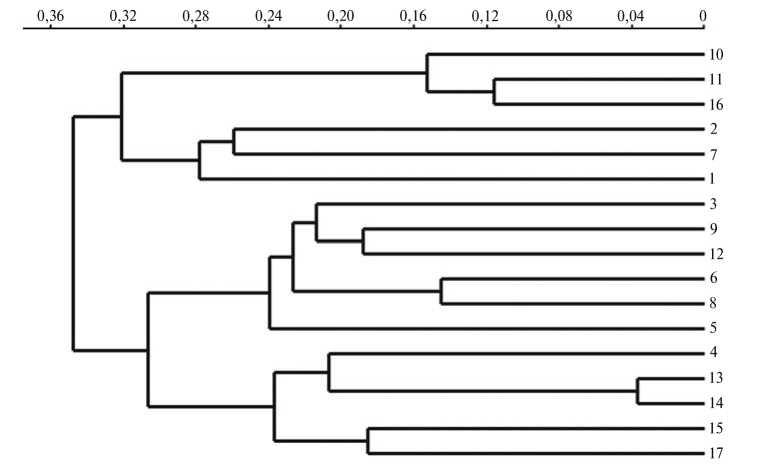
Рис. 3. Дендрограмма филогенетического родства изучаемых популяций свиней (№№ 1-17, см. табл.) , построенная по M. Nei (16) методом попарного внутригруппового невзвешенного среднего (UPGMA) на основании микросателлитного анализа по 11 локусам (ось X — генетические дистанции).
Формирующаяся структура генеалогического дерева (см. рис. 3) соответствовала описанным выше закономерностям и отражала происхождение изучаемых групп свиней. Так, популяции №№ 1, 2 и 7, образующие единую ветвь, объединяет их географическое происхождение (Канада). Популяции №№ 10, 11 и 16 представлены группой свиней французской селекции (№ 10), а также локальными популяциями, для улучшения мясных и откормочных качеств которых активно использовались свиньи французской селекции (№ 11 и № 16). По всей видимости, в единый подкластер эти популяции объединяются вследствие активного привлечения хряков породы йоркшир французского происхождения в программы разведения на канадских племенных фермах (17). Второй подкластер сформировали популяции отечественной селекции, для улучшения которых были использованы животные европейской селекции (№№ 3, 5, 6 и 8), а также европейские популяции — из Германии (№ 9) и Ирландии (№ 12). Третий подкластер объединял ветвь, представленную узкоспециализированными популяциями частных европейских компаний (№№ 4, 15 и 17), и ветвь, образованную двумя российскими популяциями свиней из Брянской области (№№ 13, 14), при разведении которых для улучшения мясных и откормочных качеств использовались хряки европейской селекции.
Следует также отметить, что популяции свиней пород крупная белая и йоркшир не расположены на генеалогическом дереве обособленно, что служит подтверждением их единого происхождение.
Таким образом, результаты наших исследований показывают, что вовлечение в исследование большого числа географически изолированных популяций позволяет дать наиболее полную характеристику аллелофонда пород. Микросателлитные профили могут быть использованы в качестве критерия оценки состояния и степени родства популяций внутри пород и отражают их происхождение.
