Оценка влияния постепенной рубки и азотного удобрения на жизнедеятельность подроста ели в северотаежных березняках

Автор: Зарубина Л.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 5, 2016 года.
Бесплатный доступ
Изучена функциональная активность про-цессов у подпологовой ели в березняках чер-ничных на северо-востоке России при ком-плексном действии постепенных рубок и азотного удобрения. Под пологом спелых бе-резняков интенсивность фотосинтеза хвои у подроста ели в 2-3-раза ниже, чем на выруб-ках. Постепенные рубки, увеличивая приток света под полог березняков, значительно ак-тивизируют физиологические процессы под-роста ели. После вырубки 52 % лиственного запаса в 59-летнем березняке черничном ин-тенсивность фотосинтеза хвои у подроста ели повысилась в 2,2-2,4 раза, усилился отток ассимилятов и улучшилось снабжение ими ростовых меристем (хвои, корней, камбия древесины ствола). Внесение в разреженные рубкой березняки азотного удобрения способ-ствовало дальнейшему усилению жизнедея-тельности у подроста ели. За пять лет дей-ствия комплексных уходов дополнительный прирост верхушечных побегов у подроста ели увеличился на 19,2-20,8 см, прирост по диа-метру увеличился в 2,8-3,4 раза. Для дости-жения положительного эффекта от ком-плексных уходов древостои северотаежных березняков черничных достаточно разредить с помощью рубки до полноты 0,5 и затем че-рез 2-3 года дополнительно внести 180 кг/га азота. Изреживание древесного яруса до пол-ноты древостоя 0,2 и внесение повышенной дозы азота не способствует усилению физио-логического состояния у подроста ели.
Березняки черничные, подрост ели, постепенные рубки, азотные удобрения, фотосинтез, пигменты, дыхание корней, рост вегетативных органов
Короткий адрес: https://sciup.org/14084695
IDR: 14084695 | УДК: 630*221
Assessment of shelterwood and nitric fertilizers on the life of spruce in north birch grove
The functional activity of processes in canopy spruce in bilberry birch groves in the north-east of Russia in the complex action of gradual cuttings and nitrogen fertilizer was studied. Under the cano-py of mature birch needles the intensity of photo-synthesis in spruce in 2-3 times lower than in clear-ings. Shelterwood, increasing the flow of light under the canopy of birch significantly activate physiologi-cal processes spruce. After cutting 52 % of decidu-ous stock in 59-year-old birch deciduous bilberry 52 % margin, the intensity of photosynthesis in needles spruce has increased in 2.2-2.4 times, increased outflow of assimilates and improved sup-ply of growth merisystem (needles, roots, wood cambium trunk). Adding to the sparse birch felling of nitrogen fertilizers contribute to the further strengthening of life in spruce. In the five years of the complex care of the additional growth of apical shoots from spruce increased by 19.2-20.8 cm in diameter growth increased by 2.8-3.4 times. In or-der to achieve a positive effect on the comprehen-sive care of birch stands of northern bilberry quite thin out using 0.5 to completeness, and then an additional 2-3 years to make 180 kg/ha of nitrogen. Thinning of the tree layer to the fullness of the stand 0.2 and the introduction of increased doses of nitrogen does not contribute to strengthening the physiological condition in spruce.
Текст научной статьи Оценка влияния постепенной рубки и азотного удобрения на жизнедеятельность подроста ели в северотаежных березняках
Введение. Лесные массивы Севера России являются крупной лесосырьевой базой, основным поставщиком древесины и ее продукции для страны и на мировой рынок. С целью получения деловой древесины с середины прошлого столетия наряду с хвойными в промышленную рубку стали широко вовлекаться спелые лиственные леса. В то же время, несмотря на относительно высокую плотность крон древостоев березняков, под их пологом имеется достаточно большое количество подроста ели [7]. Известно, что промышленные рубки в большинстве типов леса ухудшают условия минерального питания для растений. На вырубках в почвах уменьшаются запасы органического вещества, азота и других элементов, изменяются рН, зольность [10]. В целях повышения плодородия лесных почв в мировой практике лесного хозяйства в широких масштабах применяются минеральные удобрения [11]. Однако на cевере таежной зоны вопросы комплексного влияния рубок и удобрений на развитие елового подроста в мелколиственных лесах остаются слабо изученными [8].
Цель исследования: изучение влияния постепенной рубки и азотного удобрения на функционирование подроста ели в северотаежных березняках черничных.
Объекты и методы исследования. Исследования проведены в Архангельской области, которая расположена на северо-востоке европейской части Российской Федерации, в 2004– 2005 гг. В качестве опытного объекта служил подрост ели (Picea abies Karst. (Pinacea)) в березняке черничном (Betuletum myrtillosum) разного возраста. Опыты начинали проводиться через два года после проведения постепенной рубки. Для опытов было подготовлено два участка березняков. Насаждения характеризовались как однотипные, располагались в Леваш-ском участковом лесничестве (кварталы 23 и 96) Обозерского лесничества. До рубки на первом участке произрастал 53-летний двухярусный березово-еловый древостой IV класса боните- та, полнотой 0,92. Состав древостоя первого яруса – 7Б3Ос+С, второго – 10Е. Средняя высота березы – 15,2 м, диаметр – 12,1 см. Подрост еловый – 3,6 тыс. экз/га. На площади 1,2 га в осеннее-зимний период 2002 г. проведена опытная равномерно-постепенная рубка по среднепасечной технологии с трелевкой хлыстов за вершину на базе трелевочного трактора (ТДТ-55) и бензомоторной пилы «Урал». Сохранность подроста – 85–100 %. Для опытов в березняке в виде прямоугольной формы было оборудовано 4 секции с разной интенсивностью рубки деревьев господствующего полога по запасу: 35, 50, 70 % и контрольная. Площадь секций составляла 0,4 га. Сомкнутость лиственного полога в контроле составляла 0,9–1,0, после рубки она уменьшилась от 0,70 на секции с рубкой 35 % до 0,2 на секции с рубкой 70 % объема древесины.
Второй опытный участок был представлен 59-летним березняком черничным послепожар-ного происхождения. Участок занимает равнинную часть большого березового массива площадью 65 га. Средняя высота березы – 16,3 м, диаметр – 14,2 см. Древостой – III класс бонитета полнотой 0,9. Под пологом количество подроста ели – 4,3 тыс. экз/га. В осеннее-зимний период 1998–1999 гг. на площади 20 га промышленным способом проведена равномернопостепенная рубка по узкопасечной технологии с вырубкой 52 % запаса древостоя лиственных пород и вывозкой сортиментов на базе многооперационных машин Хорвестер (софит-х) + Форвардер (софит – 6ф). Ширина пасек – 24– 28 м, технологических полос – 20–23 м, волоков – 3,8–4,2 м. Состав древостоя до рубки – 7Б3Ос+С, на секции с вырубкой 52 % запаса – 8Б2Ос+С, сомкнутость полога до рубки – 0,9– 1,0, после рубки – 0,48. Для опыта на участке было оборудовано 2 площадки: опытная с промышленной рубкой 52 % запаса лиственных пород и контрольная. Исследования проводились через два года после рубки.
В рассматриваемых насаждениях дополнительно на контрольной и опытных секциях через два года после постепенной рубки были внесены азотные удобрения в виде гранулированной мочевины в дозах 180 и 270 кг на 1 га по действующему веществу (д.в). Выбор мочевины в качестве азотного удобрения диктовался теми соображениями, что в мочевине азот находится в аммиачной форме, которая, согласно [11], считается наиболее эффективным азотным удобрением для подкормки древесных пород. Удобрения внесены в почву 1 июня, поскольку на Севере уже в этот период начинается активный рост корней [4].
Измерение освещенности в березняках проводилось в июне-июле после полного облиств-ления березы и осины на высоте 1,5 м от поверхности почвы с помощью двух люксметров Ю-116М по 35–40 постоянным точкам в около-полуденные часы (13 ч.) одновременно в лесу и на открытом месте [2].
В основу методики изучения потенциального фотосинтеза и оттока 14С-ассимилятов у подроста ели был положен радиометрический метод [6]. Удельная радиоактивность газовой смеси (СО 2 + 14СО 2 ) в камере при определении фотосинтеза ели составляла 0,2 МБк/л, при определении оттока ассимилятов – 8 МБк/л. Для определения оттока ассимилятов в ассимиляционную камеру, изготовленную из прочной прозрачной полиэтиленовой пленки, помещалось 3–5 верхних мутовок подроста ели. Камера герметически соединялась с газгольдером, которым служила полиэтиленовая канистра емкостью 5 л. Прокачка радиоактивной смеси через камеру продолжалась в течение 30 мин. В течение этого периода через каждые 5 мин над камерой замерялась освещенность. Повторность каждого варианта опыта – двухкратная. Образцы хвои, корней, древесины отбирались через 0,5 и 6 ч, 1 и 8 сут. Результаты выражались в имп./мин в расчете на 1 г сухой растительной пробы.
Интенсивность транспирации определяли путем двухкратного быстрого взвешивания образцов хвои [9], интенсивность дыхания корней и хвои – методом Бойсен-Йенсена [5]. При изучении содержания пластидных пигментов пользовались спектрофотометрическим методом [13].
Замеры прироста побегов в длину у подроста ели проводили с помощью мерной линейки, замеры диаметра ствола – штангенциркулем. Все работы проводились после окончания вегетационного периода. Обработка полевых материалов осуществлялась общепринятыми в лесоводстве и таксации методами.
Результаты исследования и их обсуждение
Освещенность. Среди экологических факторов свет под пологом леса является одним из ведущих в развитии фитоценозов [2]. В лиственных фитоценозах ежегодная смена ассимиляционного аппарата у лиственных растений вносит свои коррективы в общий режим светового довольствия леса. Результаты измерений освещенности показали, что в 53-летнем березняке черничном в наиболее длинные световые дни (18–23 июня) в околополуденные часы под полог проникает менее 8,0 тыс. лк, в 59-летнем березняке – не более 8,8 тыс. лк от приходящей к кронам солнечной радиации. На секциях с рубкой (50–52 %) освещенность составляла 34,3–35,5 тыс. лк [8], что, согласно [2], соответствовало значениям, считающимися оптимальными для роста елового подроста. Безусловно, усиление светового режима в подчиненных ярусах березняков черничных после разреживания верхнего полога должно положительно повлиять на рост и функциональную деятельность ассимиляционного аппарата подроста ели.
Дыхание корней . Корни являются не только органом для снабжения растений питательными веществами и водой, но и сами участвуют во многих обменных процессах. Нормальная жизнедеятельность корней возможна лишь при благоприятных условиях водного и минерального питания почвы, текущего фотосинтеза ассимилирующими органами, потока ассимилятов, поступающего из кроны в корни [1]. Как показали исследования, в лиственно-еловых фитоценозах у подроста ели внесенный в почву азот усиливает выделение СО 2 корнями. Более интенсивное выделение СО 2 корнями подроста ели в 53-летнем березняке черничном было отмечено сразу, как только удобрения поступили в почву (табл. 1).
Таблица 1
|
Дата |
Температура воздуха, 0С |
Интенсивность рубки, % |
|||||
|
0 |
70 |
||||||
|
Контроль |
(N 180 ) |
(N 270 ) |
Контроль |
(N 180 ) |
(N 270 ) |
||
|
24.06 |
14,7 |
0,41 |
0,56 |
0,44 |
0,68 |
0,87 |
0,60 |
|
12.07 |
17,2 |
0,59 |
1,00 |
0,61 |
0,74 |
0,94 |
0,64 |
|
28.07 |
19,1 |
0,66 |
1,22 |
0,72 |
0,88 |
1,16 |
0,90 |
|
20.08 |
17,8 |
0,56 |
0,96 |
0,59 |
0,73 |
0,94 |
0,90 |
|
Среднее |
0, 53 |
0,94 |
0,59 |
0,77 |
0,98 |
0,76 |
|
|
% |
100 |
177 |
111 |
100 |
127 |
99 |
|
Влияние постепенной рубки и азота на дыхание корней подроста ели в 53-летнем березняке черничном в 2004 г., мг СО 2 /(г·ч)
В течение первого года (2004) наибольшее действие на дыхание корней оказала доза внесенного азота 180 кг/га. В неразреженной секции насаждения на площадках с указанной дозой удобрения 28 июля при температуре 19,1 0С выделяющееся при дыхании корней количество СО2 у подроста ели составляло 1,22±0,16 мг СО2 /(г·ч) и превышало контроль на 77 %. В секции с интенсивностью рубки 70 % выделенное корнями количество СО2 составляло 1,16 мг СО2 /(г·ч), что было выше на 29 %, чем в контроле. Расчеты t-критерия Стьюдента показали, что для контрольной секции критический уровень достоверности, равный 2,8, превышен лишь для варианта N180 (tфакт.= 4,7). В варианте с максималь- ной дозой азота (N270) дыхание корней в течение всего летнего периода практически оставалось на уровне контроля (tфакт.= 0,3). На секции с рубкой дыхание корней в контроле и опытных вариантах (N180, N270) в этот срок составляло 0,88±0,07, 1,16±0,13, 0,90±0,17 мг СО2 /(г·ч) при tфакт. с контролем, равным 3,2 и 0,2 соответственно. В начале вегетации количество выделяемого корнями СО2 у подроста в секции N270 было на 11–13 % меньше, чем у подроста в контроле. Как показало исследование, корни подроста ели на площадках с повышенной дозой азота не росли, светлые корневые окончания у них отсутствовали, часть их приобрела бурую окраску и в последствии отмерла. Низкий уро- вень дыхания корней у подроста ели на площадках с повышенной дозой азота в наших опытах [8] наблюдался в результате слабого обеспечения корней 14С-ассимилятами из кроны.
Аналогичные результаты влияния выборочной рубки и азота на дыхание корней елового подроста были получены нами в 59-летнем березняке черничном с промышленной рубкой березы (участок № 2). Самое высокое дыхание корней у подроста ели отмечалось на площадках с азотом на секции без рубки березы, где превышало контроль на 39–69 % (при tфакт. = = 2,9–3,7). На секции с рубкой 52 % объема древесины березы азот повысил дыхание корней подроста ели лишь на 19–36 %. Комплексное действие рубки и азота повысило дыхание корней ели в 1,9–2,3 раза.
Содержание пигментов. Как показали наши исследования, постепенная рубка в березняках черничных, увеличив поступление света под полог, привела к снижению концентрации зеленых пигментов в хвое у подроста ели. В 53летнем березняке в течение трех лет на секциях с рубкой хвоя подроста ели содержала хлорофилла на 16–29 % меньше, чем в контроле (табл. 2).
Таблица 2
Динамика накопления пигментов в хвое подроста ели в 53-летнем березняке черничном при разных объемах рубки и дозах внесенных азотных удобрений (2004–2005 гг.)
|
Дата |
Интенсивность рубки, % |
|||||||||
|
Контроль |
35 |
50 |
70 |
|||||||
|
Доза азота удобрений, кг/га д.в. |
||||||||||
|
0 |
180 |
270 |
0 |
180 |
0 |
180 |
0 |
180 |
270 |
|
|
Сумма хлорофиллов, мкг/г свежей массы хвои |
||||||||||
|
5.06.2004 |
70 |
74 |
84 |
70 |
74 |
77 |
121 |
58 |
94 |
65 |
|
20.06.2004 |
212 |
217 |
223 |
205 |
195 |
190 |
206 |
190 |
203 |
215 |
|
31.07.2004 |
650 |
743 |
826 |
565 |
594 |
555 |
712 |
539 |
728 |
828 |
|
27.08.2004 |
614 |
827 |
833 |
487 |
538 |
547 |
802 |
600 |
938 |
1030 |
|
10.10.2004 |
790 |
849 |
851 |
564 |
607 |
557 |
898 |
624 |
1017 |
1129 |
|
20.02.2005 |
743 |
890 |
943 |
700 |
565 |
672 |
811 |
548 |
985 |
1093 |
|
10.06.2005 |
762 |
814 |
843 |
570 |
657 |
644 |
767 |
658 |
884 |
812 |
|
13.08.2005 |
932 |
1093 |
1173 |
649 |
686 |
648 |
990 |
744 |
998 |
1069 |
|
Сумма каротиноидных пигментов, мкг/г свежей массы хвои |
||||||||||
|
5.06.2004 |
26 |
26 |
26 |
28 |
26 |
33 |
41 |
25 |
38 |
28 |
|
20.06.2004 |
53 |
53 |
60 |
45 |
57 |
56 |
69 |
68 |
69 |
73 |
|
31.07.2004 |
85 |
92 |
96 |
73 |
74 |
77 |
106 |
90 |
95 |
122 |
|
27.08.2004 |
79 |
114 |
115 |
94 |
101 |
94 |
130 |
79 |
144 |
120 |
|
10.10.2004 |
138 |
148 |
131 |
129 |
124 |
115 |
181 |
123 |
180 |
188 |
|
20.02.2005 |
216 |
219 |
224 |
163 |
157 |
181 |
225 |
172 |
246 |
272 |
|
10.06.2005 |
146 |
147 |
151 |
140 |
154 |
141 |
174 |
154 |
195 |
177 |
|
13.08.2005 |
126 |
144 |
145 |
87 |
99 |
93 |
120 |
103 |
150 |
164 |
Внесение в 53-летний березняк черничный азотных удобрений у подроста ели значительно усилило синтез хлорофилла и каротиноидных пигментов и привело к их накоплению в хвое. В хвое текущего года у подроста на участках, где было внесено азотное удобрение, на контрольной секции и в секциях с рубкой в первый год (2004) пигментов содержалось на 30–41 % больше, чем в аналогичной хвое контрольных растений. Замечено, что наиболее активно в хвое пигменты накапливались на максимально разреженной секции. В августе в хвое текущего года у подроста ели на площадках N270 секции с вырубкой 70 % запаса древесины березы хлорофилла содержалось 1030 мкг/г, у растений, подкормленных такой же дозой азота в секции без рубки – 833 мкг/г, или в 1,2 раза меньше. В целом азотное удобрение способствовало по- вышению содержания хлорофилла у подроста ели на секциях с 70 и 50 %-й рубкой объема древесины на 41 и 30 %, на секции без рубки – на 26 %.
Азотное удобрение увеличило концентрацию пигментов также в хвое подроста ели, возникшей еще за год до его внесения и через год после его внесения в березняк. Внесение азота в указанных дозах в 59-летний березняк у подроста ели также положительно сказалось на синтезе пигментов. Максимальное количество пигментов в хвое подроста здесь накапливалось на секции с рубкой 52 % запаса древесины и в августе превышало контроль на 22–35 % [8].
Фотосинтез хвои ели. Известно, что путем внесения в почву лесных сообществ азотных, фосфорных и калийных удобрений можно влиять на скорость и направленность углеродного питания у растений. Наши опыты показали, что после внесения в 53-летний березняк черничный азотных удобрений в дозах 180 и 270 кг/га интенсивность фотосинтеза хвои у подроста ели на секции без рубки составляла 10,5– 16,5 мг СО2 на 1 г сухой массы хвои в час или по отношению к контролю увеличилась на 3–12 %. В это же время на секциях с рубкой она за этот период выросла в 1,2–1,4 раза (табл. 3). Низкий уровень фотосинтеза у ели на удобренных площадках в не тронутом рубкой древостое, на наш взгляд, объясняется слабым использованием азота удобрения растениями в условиях недостатка света.
Таблица 3
Фотосинтез хвои, мг СО 2 /(г·ч), подроста ели в 53-летнем березняке черничном при разных объемах рубки и внесения азотных удобрений
|
Категория высоты подроста |
Контроль |
Доза азота, кг/га д.в. |
|||||
|
0 |
N 180 |
||||||
|
Интенсивность рубки, % |
|||||||
|
35 |
50 |
70 |
35 |
50 |
70 |
||
|
Крупный |
9,5±0,6 |
15,6±0,9 |
20,6±1,2 |
21,4±0,9 |
16,4±0,7 |
27,2±2,0 |
29,2±2,2 |
|
Средний |
9,8±0,8 |
15,2±0,8 |
20,8±1,3 |
24,8±1,8 |
17,2±1,3 |
25,9±1,1 |
28,9±2,2 |
|
Мелкий |
9,4±0,6 |
16,6±1,3 |
21,1±1,5 |
23,5±1,7 |
18,9±1,0 |
25,0±1,6 |
25,9±2,3 |
Проведение верховой постепенной рубки в целом положительно сказалось на эффективности действия внесенного удобрения. На удобренных площадках интенсивность фотосинтеза хвои у подроста ели возрастала по мере увеличения изреженности древесного полога. На второй год (2005) максимальная скорость фотосинтеза хвои ели в 53-летнем березняке отмечалась на удобренных площадках секции с 70 %-й интенсивностью рубки объема древесины. На данной секции интенсивность фотосинтеза хвои у ели почти в два раза превышала показатели в контроле. В этот период у формирующейся хвои (в расчете на 1 г сухой массы) фотосинтез составлял 26,2–30,2 мг СО 2 в час.
Дозы азотного удобрения (N 180 и N 270 ) оказали неоднозначное действие на фотосинтез хвои ели. В первый год (2004) повышенная доза азота на секциях с рубкой оказалась неэффективной. Наибольшее влияние на фотосинтез хвои подроста ели удобрения оказали на секции с
70 %-й интенсивностью рубки объема древесины. Однако по сравнению с секцией, в которой было вырублено 50 % лиственного запаса, различия оказались небольшими (11 %) и недостоверными. Согласно Ю.Л. Цельникер [12], причиной слабого повышения фотосинтеза хвои у подроста при повышенной освещенности могла явиться инактивация сильным светом ключевого фермента фотосинтеза РБФ-карбоксилазы. Удобрения в комплексе с постепенной рубкой по отношению к общему контролю (без рубки и удобрений) способствовали увеличению интенсивности фотосинтеза хвои у елового подроста в 2,2–3,2 раза (достоверность различий средних значений по критерию Стьюдента tфакт.= 13,6– 22,7). Результаты исследований показали, что в условиях Севера наибольшее влияние на фотосинтез хвои у подроста ели в березняках черничных оказывает интенсивность рубки (58,0– 67,8 %). Влияние азотного удобрения в этих условиях действует опосредовано, видимо, через другие физиологические процессы, и является все же менее значимым фактором для развития подроста ели по сравнению с освещенностью, несмотря даже на особую роль азота в метаболических процессах [3].
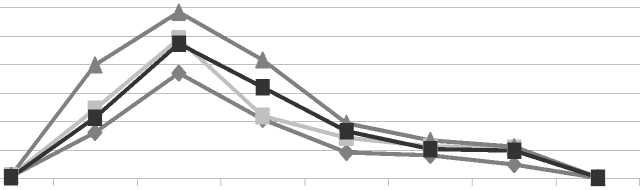
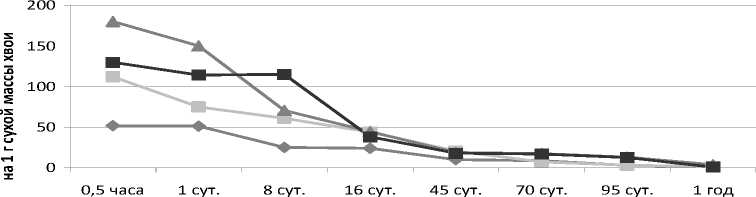
Отток углерода-14. Внесение в разреженный рубкой 59-летний березняк черничный азотного удобрения значительно активизировало у подроста ели процесс фотосинтеза и способствовало накоплению радиоактивных метаболитов (рис.). Результаты радиохимического анализа
А показали, что за время экспозиции (0,5 ч) в атмосфере 14СО2 + СО2 в процессе фотосинтеза 1-3летней хвоей ели в расчете на 1 г сухой массы хвои на секции без рубки в контроле и на опытных площадках (N180, N270) было ассимилировано 103·103; 215,1·103; 162,4·103 имп/мин. На секции с рубкой в хвое этих возрастов за указанный срок в контроле и на опытных площадках (N180, N270) было накоплено соответственно 213,3·103; 349,8·103; 257,7·103 имп/мин.
О га
га S
о
га
0,5 часа 1 сут. 8 сут.
16 сут. 45 сут. 70 сут. 95 сут. 1 год
Б
—•— контрольная секция без рубки ..... ■ — контрольная секция с рубкой
—А— N 180
■ N 270
Влияние постепенной рубки и азотного удобрения на скорость оттока 14С-ассимилятов у подроста ели в 59-летнем березняке: А – хвоя текущего года; Б – однолетняя хвоя
Как показали наши исследования, внесение азотного удобрения значительно ускорило транспорт ассимилятов по растению. Уже через сутки после подкормки на площадках с N180 радиоуглерод был обнаружен в коре, лубе, древесине ствола и дополнительно в развивающейся хвое ели. Содержание его в данной хвое было в 1,3–1,6 раза больше, чем у ели контрольных участков. В корни этих растений за сутки из кроны поступило 112 имп/мин. На площадках с N270 в корни подроста притекло лишь 5 имп/мин. В корнях контрольных растений радиоуглерод еще отсутствовал. Максимальное количество радиоуглерода в корни растений поступило спустя неделю после подкормки. В этот период у растений с N180 удельная радиоактивность корней была на 24 % выше, чем в контроле и на площадках с N270. Наиболее активно радиоуглерод накапливался в мелких корнях. Радиоактивность проводящих корней оставалась на 12– 24 % ниже радиоактивности мелких корневых окончаний.
Рост подроста ели . Обусловленность жизненного состояния от световых условий и азотного питания у подроста ели проявилась и в скорости роста в высоту и толщину. Как видно из данных таблицы 4, до внесения удобрений (1988–2000 гг.) среднепериодический прирост верхушечного побега у опытных растений в древостое и на секции с рубкой на уровне 0,95 (t = 0,2–0,4) не имел статистически значимых различий. После внесения азота (2001 г.) прирост их в высоту значительно активизировался.
Однако в первый год он был небольшим. Различия с контролем у опытных растений на обеих секциях в конце периода роста на уровне 0,95 оказались недостоверными. Корреляционная связь между сезонным ростом ели в высоту и оптимальной дозой азота (N 180 ) в этот год составила r = 0,12±0,009. Максимальный прирост в высоту у подроста под влиянием азота удобрений произошел на третий и четвертый годы (2003–2004) и превышал контроль в 2,2–2,4 раза. Наиболее активным он был на секции с рубкой.
Таблица 4
Влияние интенсивности рубки и азота на динамику роста, см, подроста ели в высоту в 59-летнем березняке черничном
|
Годы |
Интенсивность рубки, % |
|||||
|
0 |
52 |
|||||
|
Доза вносимого азота, кг/га д.в. |
||||||
|
0 |
N 180 |
N 270 |
0 |
N 180 |
N 270 |
|
|
1998-2000 |
3,7±0,4 |
3,9±0,8 |
3,4±0,8 |
3,8±0,8 |
3,5±0,6 |
3,6±0,3 |
|
2001 |
3,8±0,7 |
4,6±1,3 |
4,0±0,6 |
4,1±0,7 |
6,2±0,4 |
4,1±0,5 |
|
2002 |
4,1±0,8 |
8,1±1,2 |
7,9±0,8 |
6,8±0,5 |
11,2±2,0 |
10,4±0,8 |
|
2003 |
4,6±0,6 |
10,0±1,6 |
10,5±1,9 |
7,9±0,9 |
13,9±1,9 |
14,3±1,9 |
|
2004 |
4,4±0,4 |
9,9±2,0 |
10,7±2,2 |
8,3±1,1 |
14,3±2,1 |
14,6±2,2 |
|
2005 |
4,5±0,6 |
8,0±1,0 |
9,1±1,4 |
8,9±1,2 |
12,5±1,9 |
13,5±2,1 |
|
Всего, см |
21,4 |
40,6 |
42,2 |
36,0 |
58,1 |
56,9 |
Примечание: 1998 – год проведения выборочной рубки; 2001 – год внесения в древостой удобрений.
Общий суммарный прирост верхушечного побега за 5 лет действия азота у опытных растений (N 180 , N 270 ) на секции без рубки составил 19,2 и 20,8 см соответственно. На секции с постепенной рубкой дополнительный прирост главного побега у опытных растений (N 180 , N 270 ) по отношению к контролю (без рубки и удобрения) составил 36,6 и 35,5 см, дополнительный прирост по диаметру –1,73 и 1,35 мм. Дополнительный прирост верхушечного побега у растений на опытных участках (N 180 , N 270 ) за счет одной рубки составил 40,0 и 41,2 %, за счет удобрений – 60,0 и 58,8 %.
Выводы. Результаты исследования свидетельствуют, что недостаток света, острая корневая конкуренция за элементы питания в березняках черничных подзоны северной тайги отрицательно сказываются на углекислотном газообмене хвои и росте подпологовой ели. Постепенные рубки в спелых березняках, улучшая световые условия и сохраняя при этом лесную среду, у подроста ели значительно активизируют фотосинтетическую деятельность, ускоряют процессы оттока ассимилятами, обеспеченность активных меристем ими, что вызывает улучшение их ростовых процессов. Дополнительное внесение в разреженные рубкой березняки азотных удобрений способствует дальнейшему улучшению жизнедеятельности под-пологовой ели. Под влиянием постепенной рубки и внесения азотных удобрений интенсивность фотосинтеза хвои у подроста ели увеличивается в 2,2–3,2 раза, дыхание корней – в 1,9–2,3 раза, концентрация пигментов в хвое возросла на 22–35 %, дополнительный прирост верхушечного побега в длину за пять лет – на 36–37 см. Для достижения необходимого положительного эффекта в развитии подпологовой ели от комплексных уходов в северотаежных березняках черничных достаточно разредить древостой рубками до полноты 0,5 и затем через 2–3 года дополнительно внести 180 кг/га азота. Дальнейшее разреживание лиственного полога до полноты 0,2 и внесение повышенной дозы азота практически не вызывает усиления физиологической и ростовой активности у подроста ели.
Список литературы Оценка влияния постепенной рубки и азотного удобрения на жизнедеятельность подроста ели в северотаежных березняках
- Абражко М.А. Влияние азотных удобрений на изменение массы и фракционный состав корней ели//Лесоведение. -1986. -№ 6. -С. 75-80.
- Алексеев В.А. Световой режим леса. -М.: Наука, 1975. -280 с.
- Андреева Т.Ф. Метаболические аспекты ус-воения азота и углерода растениями при фо-тосинтезе//Фотосинтез и продукционный процесс. -М.: Наука, 1988. -С. 86.
- Бобкова К.С., Патов А.И. Сезонная динами-ка роста побегов и корней//Эколого-биологические основы повышения продук-тивности таежных лесов Европейского Севе-ра. -Л.: Наука, 1981. -С. 93-103.
- Вальтер О.А., Пиневич Л.М., Варасова Н.Н. Практикум по физиологии растений с основа-ми биохимии. -М.; Л.: Сельхозиздат, 1957. -341 с.
- Вознесенский Л.В., Заленский О.В., Семиха-това О.А. Методы исследования фотосинтеза и дыхания растений. -М.; Л.: Наука, 1965. -305 с.
- Грязькин А.В. Возобновительный потенциал таежных лесов (на примере ельников севе-ро-запада России). -СПб.: Изд-во СПбЛТА, 2001. -188 с.
- Зарубина Л.В, Коновалов В.Н. Эколого-физиологические особенности ели в берез-няках черничных. -Архангельск: Изд-во САФУ, 2014. -376 с.
- Иванов Л.А., Силина А.А, Цельникер Ю.Л. О методе быстрого взвешивания для опре-деления транспирации в естественных усло-виях//Ботан. журн. -1950. -Т. 35, вып. 2. -С. 171-185.
- Кошельков С.П., Терентьева Е.Н. Группы азота, фосфора и гумусовых соединений в почвах сплошных вырубок березняков южной тайги//Лесоведение. -1985. -№ 2. -С. 10-15.
- Победов В.С. Исследование и обоснование применения минеральных удобрений в ин-тенсивном лесном хозяйстве: автореф. дис. … д-ра с.-х. наук. -М.: Изд-во МЛТИ, 1981. -38 с.
- Цельникер Ю.Л. Физиологические основы теневыносливости древесных растений. -М.: Наука, 1978. -215 с.
- Шлык А.А. Определение хлорофиллов и ка-ротиноидов в экстрактах зеленых листьев//Биохимические методы в физиологии расте-ний. -М.: Наука, 1971. -С. 154-169.
