Окислительная модификация белков сердечной и скелетной мускулатуры крыс под влиянием субстрата синтеза оксида азота

Автор: Арапова А.И., Фомина М.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Медико-биологические науки
Статья в выпуске: 1, 2016 года.
Бесплатный доступ
Целью исследования явилось изучение влияния L-аргинина на окислительную модификацию белков сердца и скелетной мышцы. Исследование было выполнено на крысах-самцах линии Wistar, содержавшихся в типовых условиях вивария и разделённых на контрольную и экспериментальную группы, по 12 особей в каждой. Экспериментальные выборки включали группы с введением L-аргинина в дозе 500 мг/кг. Показатели изучали в цитоплазматической фракции гомогената миокарда и скелетной мускулатуры крыс. Окислительную модификацию белков в цитоплазматической фракции оценивали по методу R.L. Levine в модификации Е.Е. Дубининой, после осаждения нуклеиновых кислот 10%-ным раствором стрептомицина сульфата. По полученным значениям экс-тинкций строили спектр окислительной модификации белков и подсчитывали площадь под кривой, выражая значения в условных единицах на грамм белка (у.е./г белка). Оценка резервно-адаптационного потенциала производилась путем подсчета отношения общей площади под кривой карбонильных производных белков при спонтанном окислении протеинов к индуцированному по реакции Фентона, принимая общее количество динитрофенилгидразонов за 100%. Установлено, что воздействие L-аргинина вызывает снижение окислительной модификации белков в скелетной мышце крыс без изменений резервно-адаптационного потенциала; при этом в миокарде отмечено нарастание содержания карбонильных производных белков и истощение резервно-адаптационного потенциала.
Миокард и скелетная мышца крыс, аргинин, маркеры окислительной модификации белков, резервно-адаптационный потенциал
Короткий адрес: https://sciup.org/147204757
IDR: 147204757 | УДК: 577.1:547.495.9
Oxidative modification of proteins in cardiac and skeletal muscles of rats under the influence of the substrate of nitric oxide synthesis
The purpose of this study is to examine the effect of L-arginine on oxidative modification of proteins of heart and skeletal muscle. The study was performed on rats male Wistar, contained in standard conditions of animal house and divided into control and experimental groups, 12 animals each. The experimental sample consisted of a group with administration of L-arginine at a dose of 500 mg/kg. The indicators studied in the cytosolic fraction of the homogenate of the myocardium and skeletal muscles of rats. Oxidative modification proteins in cytoplasmic fractions was evaluated by the method of R. L. Levine in modification of E. E. Dubininoj, after precipitation of nucleic acids 10 % solution of streptomycin sulfate. On the obtained values of extinction have built a range of oxidative modification of proteins, and calculated the area under the curve, expressing values in units per gram protein (u.e./g of protein). Estimation of reserve and adaptation capacity were produced by calculating the relationship of the total area under the curve of carbonyl derived proteins in spontaneous oxidation to proteins induced by Fenton's reaction, taking the total number of dinitrophenylhydrazones 100%. Found that exposure of L-arginine causes a decrease in oxidative modification of proteins in skeletal muscle of rats without changes of reserve and adaptive capacity; in the myocardium a significant increase in the content of carbonyl derivatives of proteins and depletion of the reserve of adaptive capacity.
Текст научной статьи Окислительная модификация белков сердечной и скелетной мускулатуры крыс под влиянием субстрата синтеза оксида азота
тельных повреждений, что связано с достаточно активным образованием карбонильных производных и их стабильностью [Дубинина, Пустыгина, 2008; Абаленихина, Фомина, Исаков, 2013].
Окислительная модификация белков (ОМБ) является одним из наиболее ранних проявлений поражения тканей при свободно-радикальной патологии [Губский и др., 2005]. Процесс старения заметно увеличивает рост чувствительности белков к окислению, в результате чего происходит накопление в тканях их окисленных форм. Достоверно известно, что количество окисляющихся белков в клетке обусловлено генетически и является ее постоянной фенотипической характеристикой [Еременко, Малоштан, 2014].
Накопление окисленных протеинов трактуется в качестве фактора регуляции синтеза и распада белков и активации протеолитических ферментов, которые избирательно разрушают окисленные формы. Этот механизм является проявлением антиоксидантной защиты организма, поэтому оценка степени ОМБ может считаться одним из наиболее надежных индикаторов свободно-радикального повреждения тканей, клеток и мембран [Еременко, Малоштан, 2014].
Материалы и методы исследований
Работа была выполнена на 24 конвенциональных половозрелых крысах-самцах линии Wistar массой 280-320 г, разделенных на две группы: контрольную и экспериментальную. Содержание лабораторных животных и выведение из эксперимента осуществлялось согласно «Санитарным правилам по устройству, оборудованию и содержанию экспериментально-биологических клиник» от 06.04.1993; и в соответствии с правилами, изложенными Международным Советом Медицинских Научных Обществ (CIOMS) в «Международных рекомендациях по проведению медикобиологических исследований с использованием животных» (1985 г.) и приказе МЗРФ №267 от 19.06.2003 г. «Об утверждении правил лабораторной практики».
Животным экспериментальной группы внутри-желудочно вводился раствор L-аргинина («Sigma». США) на 0.9%-ном растворе NaCl в дозе 500 мг/кг [Покровский и др., 2008]. Препарат вводили 1 раз в сутки до утреннего кормления ежедневно в течение 10 дней.
Контрольная группа была сформирована из животных, сопоставимых по возрасту, массе и условиям содержания с экспериментальными особями. Животным контрольной группы вводили физиологический раствор по схеме, совпадающей с таковой для экспериментальной группы.
После обескровливания под эфирным рауш-наркозом при сохраненном дыхании и сердцебиении осуществляли извлечение сердца и скелетной мышцы передней поверхности бедра с помещением тканей в охлажденный 0.25 М раствор сахарозы. Гомогенизация участков ткани левого желудочка сердца и скелетной мышцы передней поверхности бедра осуществлялась с помощью аппарата «Potter S» (Sartorius, Германия) в охлаждённом 0.25 М растворе сахарозы в соотношении 1/10 в течение 60 сек. при скорости вращения тефлонового пестика 1500 об/мин.
Полученные гомогенаты подвергали дифференциальному центрифугированию для получения чистой цитоплазматической (неседиментируемой) фракции, в которой оценивали содержание ОМБ по методу R.L. Levine в модификации [Дубинина и др., 1995], после предварительного осаждения нуклеиновых кислот 10%-ным раствором стрептомицина сульфата.
По полученным результатам строили спектр поглощения продуктов ОМБ и подсчитывали площадь под полученной кривой [Пат., 2014]; значения выражались в условных единицах на грамм белка (у.е./г белка).
Оценка резервно-адаптационного потенциала (РАЛ) производилась путем подсчета отношения общей площади под кривой карбонильных производных белков при спонтанном окислении протеинов к индуцированному по реакции Фентона, принимая общее количество динитрофенилгидразонов за 100% [Никитина, Мухина, 2009].
Статистическая обработка анализа результатов исследования проведена согласно руководствам по медицинской статистике с использованием программы «Microsoft Office Excel 2010» и «Statistica 10.0». Проверку нормальности распределения данных осуществляли с помощью критерия Шапиро-Уилка (W-критерий). Поскольку отмечалось отсутствие согласия большинства данных с нормальным распределением, вычисляли характеристики: медиану, минимальное и максимальное значения, результаты представляли в формате Me [min; max], для оценки статистической значимости различий независимых выборок использовали ранговый критерий Манна-Уитни (U-тест).
Результаты и их обсуждение
В скелетной мышце под влиянием аргинина в дозе 500 мг/кг в экспериментальной группе животных наблюдается незначительное снижение содержания карбонильных производных нейтрального характера по сравнению с контрольными значениями (табл. 1, рис. 1). Статистически значимо уменьшается содержание альдегид-динитрофенилгидразонов (АДНФГ) нейтрального характера в ультрафиолетовой части спектра. Снижение суммарного содержания карбонильных производных белков (статистически значимое) в экспериментальной группе по отношению к контрольным показателям может свидетельствовать о запуске механизмов вторичной антиоксидантной защиты в тканях [Ду- тканях [Дубинина, Пустыгина, 2008; Абаленихина, дификации белков в ткани скелетной мышцы пе-Фомина, 2014], с подавлением окислительной мо- редней поверхности бедра.
Таблица 1
Площадь под кривой спонтанной окислительной модификации белков (у.е./г белка); Me [min; max]
|
Показатель |
SАДНФГт |
S АД НФ Tvs |
SКДНФГт |
S КДНФГ\5 |
БОМБ |
|
|
Скелетная мышца |
Контроль |
23.17 [22.34; 24.78] |
6.30 [5.78; 6.81] |
4.26 [4.26; 6.31] |
1.06 [1.05; 1.28] |
35.01 [33.43; 38.96] |
|
Эксперимент |
17.36 [14.24; 20.21] * |
4.04 [2.71; 6.31] |
4.50 [2.69; 5.85] |
0.88 [0.34; 1.34] |
27.08 [21.80; 33.15] * |
|
|
Сердечная мышца |
Контроль |
21.28 [19.58; 22.80] |
3.25 [2.28; 5.21] |
3.28 [0.99; 4.77] |
0.44 [0.42; 0.71] |
28.23 [23.79; 32.99] |
|
Эксперимент |
18.97 [18.30; 19.10] * |
6.23 [5.41; 8.02] * |
5.24 [4.79; 7.79] * |
0.89 [0.83; 1.27] * |
31.33 [29.64; 36.17] |
|
*- статистически значимые отличия от группы контроля (р<0.05).
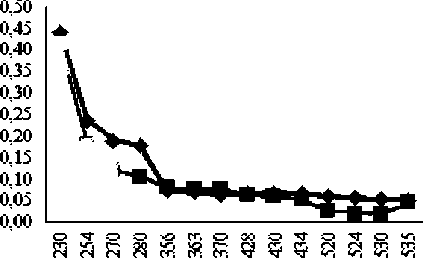
* контроль fl Аргинин
Рис. 1. Сравнительный анализ спектра поглощения продуктов спонтанной окислительной модификации белков и их компонентов в скелетной мышце контрольной и экспериментальной групп животных (у.е./г белка)
Изменения, касающиеся левого желудочка сердца крысы, характеризуются снижением АДНФГ нейтрального характера; нарастанием АДНФГ основного характера и обеих групп кетон-динитрофенилгидразонов (КДНФГ), результаты статистически значимо отличаются от контрольной группы крыс (табл. 1, рис. 2).
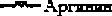
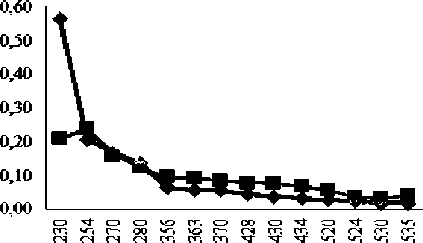
Рис. 2. Сравнительный анализ спектра поглощения продуктов спонтанной окислительной модификации белков и их компонентов в миокарде контрольной и экспериментальной групп животных (у.е./г белка)
В результате последних исследований принято альдегидные производные считать ранними маркерами ОМБ, характеризующими процесс фрагментации, а кетонные - поздними, отражающими агрегацию белков, результатом которой может оказаться нарушение их нативной конформации [Губский и др., 2005].
Образование альдегидов происходит в условиях стимуляции свободно-радикальных превращений в клетках, которые имеют место при оксидативном стрессе [Давыдов, Божков, 2014]. Ввиду уже доказанной причинно-следственной связи между карбонильным и оксидативным стрессом [Давыдов, Божков, 2014], можно предположить, что в результате воздействия субстрата синтеза оксида азота на сердце и скелетную мышцу крыс происходит статистически значимый рост показателей ранних маркеров карбонильного стресса относительно контрольной группы животных (табл. 2, рис. 3), вследствие активации образования альдегидов.
Контроль. Контроль.
15%
13%

85% 87%
Скелетная мышца.
Сердечная мышца.
19% 20%

доля АДНФГ (первичные маркеры) ■ доля КДНФГ (вторичные маркеры)
Рис. 3. Доля первичных и вторичных маркеров относительно общего содержания карбонильных производных белков
В изучаемых образцах мускулатуры крыс наблюдается уменьшение доли вторичных маркеров (КДНФГ), что может свидетельствовать о сниже- нии степени ОМБ или о повышении скорости утилизации измененного белка (вторичный антиоксидантный эффект) [Владимирский. Бородина. Аба шева. 2013]. Статистически значимое отличие от группы контроля наблюдается в ткани сердца крыс под влиянием аргинина.
Таблица 2
Суммарное содержание первичных и вторичных маркеров окислительного стресса экспериментальной и контрольной групп животных (у.е./г белка); Me [min; max]
|
Показатель/ группа |
S АДНФГиу + S АДНФГта |
S КДНФГ„ + S КДНФГ,, |
|
|
Скелетная мышца |
Контроль |
29.48 [28.12; 31.59] |
5.33 [5.31; 7.37] |
|
Эксперимент |
21.39 [17.46; 26.06] * |
5.38 [3.26; 7.09] |
|
|
Сердечная мышца |
Контроль |
24.53 [22.38; 27.50] |
3.72 [1.42; 5.48] |
|
Эксперимент |
25.21 [24.02; 27.11] |
6.13 [5.62; 9.06] * |
|
*- статистически значимые отличия от группы контроля (р<0.05).
Оценка содержания динитрофенилгидразонов при спонтанном окислении белков позволяет охарактеризовать степень окислительной деструкции белковой молекулы, а при стимулированном (ме-талл-катализируемом) окислении - описать изменения резервно-адаптационных возможностей организма [Владимирский. Бородина. Абашева. 2013]. Металл-катализируемое окисление в настоящее время рассматривают как посттранскрипционную окислительную модификацию белков [Дубинина. 2006].
Исходя из данных литературы [Давыдов. Божков. 2014]. можно сделать следующее предположение: аргинин, являясь донатором синтеза оксида азота. увеличивает степень окислительно-го/карбонильного стресса [Капелько и др.. 2008; Давыдов. Божков. 2014]. тем самым уменьшая степень РАП относительно групп контроля (рис. 4. 5).
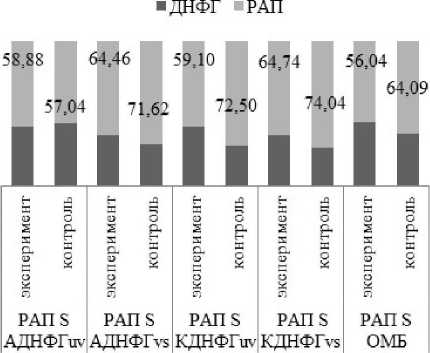
Рис. 4. Оценка резервно-адаптационного потенциала ткани скелетной мышцы аргинин/контроль
Однако в группе воздействия на сердечную мышцу аргинином (рис. 4). РАП S АДНФГ,,, незначительно статистически значимо увеличился по сравнению с контролем. Возможно, причиной та ковых изменений является дефицит нитроксида. при котором динитрозильные комплексы негемового железа (ДНКЖ). содержащие цистеин или глутатион [Шумаев. Космачевская. Топунов, 2008]. распадаются, освобождая NO из собственных депо [Капелько и др., 2008; Абаленихина. Фомина. 2014].
■ ДНФГ -РАП
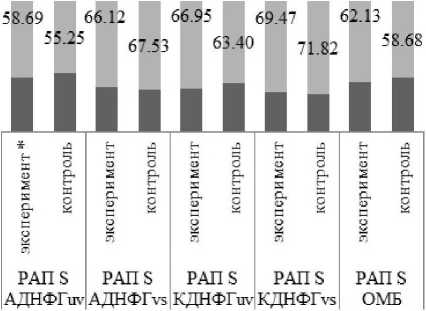
Рис. 5. Оценка резервно-адаптационного потенциала ткани сердечной мышцы аргинин/контроль
* - статистически значимые отличия от группы контроля (р<0.05)
Динитрозильные комплексы негемового железа обнаружены во многих клетках и тканях, продуцирующих оксид азота. ДНКЖ являются стабильными депо NO. в свою очередь, низкомолекулярные ДНКЖ выполняют функцию его переносчика [Граник. Григорьев. 2004]. Таким образом, динитрозильные комплексы негемового железа могут образовываться в клетках при умеренном окислительном стрессе и тем самым формировать собственный внутриклеточный пул NO как форму запасания оксида азота. Вместе с тем при снижении уровня NO окислительный стресс активирует его выход из комплексов с последующим демпфиро- ванием повреждающего действия активных форм кислорода [Капелько и др., 2008].
Заключение
Таким образом, результаты исследования демонстрируют, что применение аргинина в дозе 500 мг/кг у экспериментальных крыс вызывает снижение окислительной модификации белков в скелетной мышце за счет уменьшения образования аль-дегид-динитрофенилгидразонов, как первичных маркеров оксидативного стресса без существенных изменений резервно-адаптационного потенциала. Миокард в тех же условиях демонстрирует тенденцию к нарастанию карбонильных производных белков, в большей степени кетон-динитрофенилгидразонов, и истощению резервноадаптационного потенциала.
Список литературы Окислительная модификация белков сердечной и скелетной мускулатуры крыс под влиянием субстрата синтеза оксида азота
- Абаленихина Ю.В., Фомина М.А., Исаков С.А. Окислительная модификация белков и изменение активности катепсина L селезенки крыс в условиях моделирования дефицита синтеза оксида азота//Российский медико-биологический вестник им. академика И.П. Павлова. 2013. № 1. С. 45-49
- Абаленихина Ю.В., Фомина М.А. Влияние модуляторов синтеза оксида азота на активность и аутопро-цессинг катепсина в иммунокомпетентных органов крыс в условиях in vitro//Наука молодых (Eruditio Juvenium). 2014. № 1. С. 53-59
- Владимирский Е.В., Бородина Е.Н., Абашева Н.М. Клиническая эффективность и безопасность включения в рацион питания пациентов с кардиоваскулярной патологией диетического профилактического продукта «масло «Гармоникум с живицей»//Современные проблемы науки и образования. 2013. № 5. URL: http://www.scienceeducation.ru/ru/article/view?id=10489 (дата обращения: 15.12.2015)
- Граник В.Г., Григорьев Н.Б. Оксид азота (NO). Новый путь к поиску лекарств: монография. М.: Вузовская книга, 2004. 360 с
- Губский Ю.И. и др. Токсикологические последствия окислительной модификации белков при различных патологических состояниях (обзор литературы)//Современные проблемы токсикологии. 2005. Т. 8, № 3. С. 20-27
- Давыдов В.В., Божков А.И. Карбонильный стресс как неспецифический фактор патогенеза (обзор литературы и собственных исследований)//Журнал НАМН Украши. 2014. Т. 20, № 1. С. 25-34
- Дубинина Е.Е. и др. Окислительная модификация белков сыворотки крови человека, метод её определения//Вопросы мед. химии. 1995. Т. 41, № 1. С. 24-26
- Дубинина Е.Е. Продукты метаболизма кислорода в функциональной активности клеток (жизнь и смерть, созидание и разрушение). Физиологические и клинико-биохимические аспекты. Санкт-Петербург: Медицинская пресса, 2006. С. 276282
- Дубинина Е.Е., Пустыгина А.В. Окислительная мо дификация протеинов, ее роль при патологических состояниях//Укр. бiохiм. журн. 2008. Т. 80, № 6. С. 5-18
- Еременко Р.Ф., Малоштан Л.Н. Оценка влияния корректора белкового обмена экстракта из травы люцерны посевной на процессы окисления белков//Медицина и образование в Сибири. 2014. № 2. URL: http://ngmu.ru/cozo/mos/article/pdf.php?id=1351 (дата обращения: 15.12.2015)
- Капелько В.И. и др. Динитрозильные комплексы железа -естественные доноры оксида азота в организме//Патолопя. 2008. Т. 5, № 3. С. 76. URL: http://nbuv.gov.ua/j-pdf/pathology_2008_5_3_58.pdf (дата обращения: 15.12.2015)
- Никитина Ю.В., Мухина И.В. Изменения окислительных процессов в ткани головного мозга и крови крыс в раннем онтогенезе//Вестник Нижегородского университета им. Н.И. Лобачевского. 2009. № 6(1). C. 124-131
- Пат. 2524667 РФ, МПК G01N33/52. Способ комплексной оценки содержания продуктов окислительной модификации белков в тканях и биологических жидкостях/Фомина М.А. Абаленихина Ю.В., Фомина Н.В., Терентьев А.А. 2014. Бюл. № 21. 8 с
- Покровский М.В. и др. Эндотелиопротекторные эффекты L-аргинина при моделированиии дефицита окиси азота//Экспериментальная и клиническая фармакология. 2008. Т. 71, № 2. С. 29-31
- Шумаев К.Б., Космачевская О.В., Топунов А.Ф. Оксид азота -с гемоглобином и не только//Химия и жизнь. 2008. № 4. С. 22-25. URL: http://files.school-collection.edu.ru/dlrstore/2ef88c65-1ae6-4eb0-d808-7b39a08283b1/22-25_04_2008.pdf (дата обращения: 15.12.2015)
