Оксид азота и сульфид водорода как р38-зависимые регуляторы экспрессии генов XIAP и AVEN в опухолевых клетках линии Jurkat

Автор: Таширева Л.А., Старикова Е.Г., Новицкий В.В., Васильева О.А., Якушина В.Д., Степовая Е.А., Бельдягина Е.В., Зима А.П., Прохоренко Т.С., Осихов И.А., Saprina T.V.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 5 (53), 2012 года.
Бесплатный доступ
Проведено исследование р38-зависимых механизмов влияния газовых трансмиттеров оксида азота и сульфида водорода на экспрессию генов XIAP и AVEN в клетках линии Jurkat. Методом ПЦР в реальном времени показано, что экспрессия генов XIAP и AVEN активна в опухолевых клетках линии Jurkat. Доноры газов оксида азота и сульфида водорода вызывали подавление экспрессии генов XIAP и AVEN в клетках Т-лимфобластной лейкемии. Изменение экспрессии указанных генов под действием газов NO и H 2S происходило с вовлечением р38 МАР киназы.
Апоптоз, оксид азота, сульфид водорода, р38 мар киназа
Короткий адрес: https://sciup.org/14056261
IDR: 14056261 | УДК: 616-091.818:546.172.6:546.221.1:576.3
Nitric oxide and hydrogen sulphide as p38-dependent regulators of XIAP and AVEN genes expression in Jurkat cells
In this paper we investigated p38-dependent mechanisms of gaseous transmitters nitric oxide and hydrogen sulfide influence on XIAP and AVEN genes expression in Jurkat cells. Using the method of real-time PCR, XIAP and AVEN genes expression in cancer Jurkat cells was shown. Donors of gases of nitric oxide and hydrogen sulfide were negative regulators of the XIAP and AVEN gene expression in the cells of T-lymphoblast leukemia. Changes in the expression of above mentioned genes under NO and H2S influence occurred with the involvement of p38 MAP kinase
Текст научной статьи Оксид азота и сульфид водорода как р38-зависимые регуляторы экспрессии генов XIAP и AVEN в опухолевых клетках линии Jurkat
Апоптоз является программированной формой клеточной гибели, посредством которой в организме происходит удаление, в том числе и определенных клонов дифференцирующихся клеток. Нарушение этого механизма регуляции клеточного гомеостаза является одним из условий опухолевого перерождения клеток. Регуляция апоптоза представляет собой сложную многокомпонентную систему баланса про- и антиапоптогенных факторов. В последние годы к числу модуляторов програм- мированной клеточной гибели относят семейство газовых трансмиттеров, состоящее из 3 газов – NO, CO и H2S [15].
Известно, что NO и H2S могут изменять реализацию программированной гибели клеток за счет влияния на функционирование основных эффекторных молекул апоптоза – каспаз, в частности, показано, что оксид азота способен ингибировать активность каспазы-3 посредством ее S-нитрозилирования [3]. До недавнего времени запуск проапоптотических реакций, следующих после активации каспаз, считался необратимым. Однако существуют протеины, способные ингибировать функции каспаз даже после их энзиматической активации. К указанным белкам относят протеины группы xIAР и белок Aven. Aven нарушает процесс самосборки комплекса Apaf1/каспаза-9 [5]. Протеины группы xIAР непосредственно ингибируют активность каспаз-3, -7 и -9 [13]. Регуляция функции белков xIAР и Aven в условиях изменения концентрации внутриклеточных газовых трансмиттеров не изучена.
Показано, что внутриклеточные газовые трансмиттеры участвуют в регуляции различных внутриклеточных процессов, в том числе за счет изменения экспрессии генов. Так, оксид азота активирует транскрипционные факторы NF-kB и АР-1, что приводит к увеличению экспрессии генов гемоксигеназы-1, белка теплового шока 70, IL-8 , ДНК-связывающего фактора c-Fos и нейроспе-цифичного белка Zif/268. Известно, что факторы транскрипции HIF1 и CREB также являются мишенями действия NO [8, 12]. Рядом авторов продемонстрировано вовлечение р38 МАРК в опосредуемую газами модуляцию клеточных функций. S. Adhkari, M. Bhatia [2] показали, что изменение содержания белков Bax и Bcl-2 при действии сульфида водорода на клетки поджелудочной железы зависит от р38 МАР киназы. В данном исследовании предпринята попытка оценить влияние стресс-активируемой р38 МАРК на экспрессию генов белков-ингибиторов каспаз xIAР и Aven в условиях изменения внутриклеточной концентрации газов оксида азота и сульфида водорода.
Материал и методы
В работе использованы клетки линии Jurkat (T-лимфобластная лейкемия), полученные из Российской коллекции клеточных культур позвоночных Института цитологии РАН (г. Санкт-Петербург). Клетки культивировали суспензионным методом в чашках Петри («Биомедикал», Россия ) в атмосфере 5 % CO2 при температуре 37ºС (СО2-инкубатор Sanyo, Япония) в полной питательной среде (90 % RPMI-1640 («Вектор-Бест», Россия), 10 % эмбриональной телячьей сыворотки («Invitrogen», США), инактивированной при 56ºС в течение 30 мин, 0,3 мг/мл L-глутамина («Вектор-Бест», Россия) и 100 мкг/мл гентамицина («INS», США)). Изменение внутриклеточной концентрации газовых трансмиттеров осуществляли с использованием донора сульфида водорода (натрий гидросульфид гидрат (NaHS) («Sigma», США)) и донора оксида азота (натрия нитропруссид (SNP) («Sigma», США)) в подобранной ранее проапоптотической концентрации 10 и 100 мМ, соответственно [1].
Для ингибирования р38 МАР-киназы к клеткам линии Jurkat добавляли селективный ингибитор р38 МАР-киназы SB 203580 в концентрации 0,2 мкМ и инкубировали в течение 20 мин. Указанные условия культивирования вызывают ингибирование активности р38 МАР-киназы на 80–95 % [10].
Для количественного определения уровня мРНК генов XIAP и AVEN использовали метод ПЦР в режиме «реального времени». Выделение тотальной РНК из клеток осуществляли сорбентноколоночным методом (QIAmp RNA Blood mini Kit, Qiagen, Германия) согласно прилагаемой инструкции. Качество всех тестируемых образцов тотальной РНК оценивалось по наличию в агарозном геле-электрофорезе двух бандов, соответствующих 18S и 28S субъединицам рРНК.
Следующим шагом синтезировали кДНК на матрице мРНК при участии обратной транскриптазы. Для этого готовили реакционную смесь: 0,5–2 мкг РНК, 100 нг статистического праймера (N9), RT-буфер (20 мМ Tris-HCl (pH=8,3), 5 мМ MgCl2, 10 мМ DTT, 100 мМ KCl, 0,4 мМ dNTP), 200 ед. акт. ДНК-зависимой РНК полимеразы (MMuLV-RT) («Promega», США).
Полученный фрагмент кДНК амплифициро-вали методом ПЦР в режиме реального времени с использованием SYBR Green I («Мolecular Probe», США) на амплификаторе Mini Opticon («Bio-Rad», США). Реакционная смесь содержала K25 буфер (×1), 0,2mM dNTP, 300 nМ праймеров, 0,5 ед. акт. SmartTaq ДНК-полимеразы блокированной антителами («Диалат», Россия), SYBRGreen I [1:25000] и 0,1–5 нг кДНК.
Праймеры, позволяющие специфично амплифи-цировать фрагменты кДНК генов, представлены в табл. 1. Для нормализации начального количества мРНК в образце и эффективности обратной транскрипции измеряли количество кДНК гена β-актин (гена «домашнего хозяйства», в относительно равной степени экспрессирующегося во всех клетках [4]). Для определения относительного количества кДНК в образце использовался метод ddCt. Результаты выражали в условных единицах (отношение относительного количества кДНК исследуемого гена к относительному количеству кДНК гена «домашнего хозяйства»).
Апоптотически измененные клетки регистрировали с помощью проточной цитофлуориметрии (FACSCantoII («Beckman Dickenson», США)) с использованием ФИТЦ-меченного аннексина V, обладающего сродством к мембранно-связанному фосфатидилсерину.
Статистический анализ проводили с использованием программного обеспечения SPSS 11.0 (SPSS, Chicago, USA). Для каждой выборки вычисляли средневыборочные характеристики. Проверку нормальности распределения количественных показателей проводили с использованием критерия Колмогорова–Смирнова. Для оценки достоверности различий независимых выборок использовали ранговый критерий Манна–Уитни. Различия считались достоверными при уровне значимости р<0,05.
Результаты и обсуждение
Молекулярные пути программированной гибели клеток контролируются генами, которые промоти-руют или ингибируют активацию каскада каспаз. Нарушение баланса между про- и антиапопто-тическими регуляторами может вносить существенный вклад в устойчивость к апоптозу. xIAP (X-связанный белок – ингибитор апоптоза) – это член семейства IAP, являющийся внутриклеточным антиапоптотическим белком. Данный протеин непосредственно ингибирует каспазу-3, -7 и -9 [13]. Кроме того, xIAP обладает E3 лигазной активностью и способен убиквитинировать каспазу-3, вызывая ее деградацию. Таким образом, xIAP действует как мощный ингибитор суицидальной программы клетки, эффективно предотвращая апоптоз, вызванный разнообразием внеклеточных стимулов, включая противоопухолевые препараты и ионизирующее излучение [9]. Воздействие доно- ров газов NO и H2S приводило к статистически значимому увеличению апоптотически-измененных клеток Т-лимфобластной лейкемии по сравнению с интактной культурой (контроль) (р<0,05) (рис. 1). При этом инкубация с 100 мМ SNP значимо уменьшала уровень мРНК гена XIAP по сравнению с контролем (р<0,05). Аналогичная тенденция была выявлена при изменении внутриклеточной концентрации сульфида водорода с использованием NaHS. Добавление указанного донора в концентрации 10 мМ к культуре клеток линии Jurkat снижало уровень мРНК гена XIAP в 25 раз по сравнению с интактными клетками Т-лимфобластной лейкемии (р<0,05) (табл. 2). Возможно, оксид азота и сульфид водорода подавляют экспрессию гена XIAP посредством S-нитрозилирования и S-сульфгидрирования транскрипционных факторов соответственно [8, 15]. Таким образом, снижение экспрессии гена XIAP, являющегося ингибитором каспаз, опосредует проапоптотический эффект газовых трансмиттеров.
Показано, что одним из элементов системы индукции апоптоза является МАР (mitogen activated protein)-киназный каскад. MAPK являются серин/ треониновыми протеинкиназами, которые могут фосфорилировать цитоплазматические и ядерные мишени [11]. Семейство МАР-киназ включает 4 небольших подсемейства протеинкиназ – p38, ERK (extracellular signal-regulated kinases), JNK/SAPK (c-jun N-terminal or stress-activated protein kinases) и BMK1 (ERK/big MAP kinase 1) [16]. Ингибирование р38-зависимых путей на фоне воздействия SNP и NaHS, как показали результаты настоящего исследования, приводило к снижению количества клеток с признаками апоптоза в культуре клеток Т-лимфобластной лейкемии по сравнению с контролем (р<0,05) (рис. 1). Предварительное инкубирование клеток линии Jurkat с селективным
Структура специфических праймеров
Таблица 1
|
Ген |
Праймеры |
Размер продукта, п.о. |
|
β-actin |
F: 5′- CTGGCACCCAGCACAATG -3′ |
72 |
|
R: 5′- AGCGAGGCCAGGATGGA -3′ |
||
|
AVEN |
F: 5′-GATTTCAGTGTCCTCCTTAG-3′ |
253 |
|
R: 5′-CCTTGCCATCATCAGTTCTC-3′ |
||
|
XIAP |
F: 5′-ATAGTGCCACGCAGTCTACAA-3′ |
101 |
|
R: 5′-AGATGGCCTGTCTAAGGCAAA-3′ |
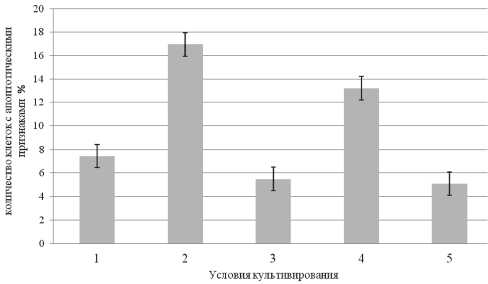
Рис. 1. Число апоптотически измененных клеток при различных условиях культивирования. Примечание: 1 – интактные клетки линии Jurkat; 2 – клетки линии Jurkat при добавлении 100 мМ SNP;
3 – клетки линии Jurkat при добавлении 100 мМ SNP и 0,2 мкМ SB203580; 4 – клетки линии Jurkat при добавлении 10 мМ NaHS;
5 – клетки линии Jurkat при добавлении 10 мМ NaHS и 0,2 мкМ SB203580
ингибитором р38 МАР киназы частично отменяло снижение экспрессии гена XIAP, вызванное обработкой клеток донором оксида азота (р<0,05), однако оставалась достоверно ниже контрольных значений (р<0,05). Совместная инкубация клеток с проапоптотической концентрацией донора сульфида водорода и 0,2 мкм SB203580 не приводила к достоверному изменению экспрессии гена XIAP по сравнению с изолированным действием газа (р>0,05) (табл. 2). Полученные данные свидетельствуют о том, что трансдукция проапоптотического сигнала в условиях изменения внутриклеточной концентрации оксида азота частично осуществляется с участием р38 МАР-киназы. Сульфид водорода является p38-независимым негативным регулятором экспрессии гена XIAP.
Известно, что другой белок Aven также способен ингибировать апоптоз, оказывая влияние на активность каспаз. Показано, что Aven ингибирует самосборку Apaf-1 и таким образом уменьшает Apaf-1-опосредованную активацию каспаз. Aven ингибирует активность каспаз в беcклеточных экстрактах и подавляет апоптоз, индуцированный комплексом Apaf-1/каспаза-9 [5]. Кроме того, экспрессия гена AVEN отдельно или в комбинации с Bcl-xL увеличивает выживаемость клеток линии CHO после воздействия апоптотических стимулов, включая истощение или полное лишение питательной среды [6]. Показано, что Aven связывается с Bcl-xL, усиливая антиапоптотические свойства последнего в ответ на каспаза-1-индуцированный апоптоз [5].
Как показали результаты настоящего исследования, при инкубировании клеток линии Jurkat с 10 мМ NaHS и 100 мМ SNP экспрессия гена АVEN значимо снижалась – в 6,5 и в 7,5 раза соответственно (р<0,05). При этом экспозиция указанных доз доноров газовых трансмиттеров приводила к усилению апоптоза клеток линии Jurkat, что подтверждает антиапоптотические свойства Аven. Выключение р38-зависимых сигнальных путей в клетках линии Jurkat с помощью селективного ингибитора приводило к статистически значимому
Таблица 2
Уровень мРНК генов XIAP и AVEN в условиях воздействия доноров газов NO и H2S и ингибитора р38 МАРК
|
Условия эксперимента |
Экспрессия гена XIAP , усл.ед. |
Экспрессия гена AVEN , усл.ед. |
|
Интактные клетки линии Jurkat |
4,97 (4,88–5,4) |
1,99 (1,63–2,0) |
|
Клетки линии Jurkat при добавлении 100 мМ SNP |
0,24 (0,21–0,27) p1<0,05 |
0,31 (0,3–0,32) p1<0,05 |
|
Клетки линии Jurkat при добавлении 100 мМ SNP и 0,2 мкМ SB203580 |
0,92 (0,73–1,35) p1<0,05 р2<0,05 |
0,45 (0,37–0,46) p1<0,05 р2<0,05 |
|
Клетки линии Jurkat при добавлении 10 мМ NaHS |
0,2 (0,19–0,21) p1<0,05 |
0,26 (0,22–0,28) p1<0,05 |
|
Клетки линии Jurkat при добавлении 10 мМ NaHS и 0,2 мкМ SB203580 |
0,17 (0,17–0,2) p1<0,05 р3>0,05 |
0,4 (0,36–0,53) p1<0,05 р3<0,05 |
Примечание: р1 – различия, статистически значимые по сравнению с интактной культурой линии Jurkat; р2 – различия, статистически значимые по сравнению с интактной культурой линии Jurkat и при воздействии 100 мМ SNP; р3 – различия, статистически значимые по сравнению с интактной культурой линии Jurkat и при воздействии 10 мМ NaHS.
СИБИРСКИЙ ОНКОЛОГИЧЕСКИЙ ЖУРНАЛ. 2012. № 5 (53)
увеличению уровня мРНК гена АVEN по сравнению с интактной культурой Jurkat-клеток в ответ на добавление в культуральную среду как донора оксида азота, так и донора сульфида водорода (р<0,05), однако по сравнению с интактной культурой клеток Т-лимфобластного лейкоза экспрессия гена AVEN оставалась сниженной (р<0,05) (табл. 2).
Таким образом, полученные данные свидетельствуют, что газы оксид азота и сульфид водорода модулируют апоптотическую программу клеток линии Jurkat частично за счет вовлечения р38-зависимых механизмов изменения экспрессии генов XIAР и АVEN . Поскольку повышение экспрессии ингибиторов апоптоза обнаружено во многих опухолях ассоциированно с прогрессией, неблагоприятным прогнозом и устойчивостью к химиотерапии [7, 14], перспективны дальнейшие исследования газовых трансмиттеров в плане возможного использования в качестве агентов для негативного регулирования экспрессии генов XIAР и АVEN в опухолевых клетках.
Работа выполнена при поддержке Министерства образования и науки России в рамках Федеральной целевой программы «Научные и научно-педагогические кадры инновационной России на 2009–2013 годы» (Государственное соглашение № 8302, ГК № П1311).
