Опасность новых рас заразихи для подсолнечника в России и меры предупреждения возможного ущерба

Автор: Дьяков А.Б., Васильева Т.А., Бойко Ю.Г.
Статья в выпуске: 1 (138), 2008 года.
Бесплатный доступ
Утверждается, что преодоление из-за гена Or3 подсолнечника резистентность заразихи могут рассматриваться как растения расы D только после проверки вирулентности потомства. Этот факт и теоретический анализ доказывают, что рекомендация о том, чтобы подавлять ядовитые расы в течение неограниченного времени, путем стабилизации отбора для повышения концентрации расы C в популяциях заразихи, ошибочна. Установленная ситуация требует оперативного реагирования на состояние расследований заражённых инфекций в регионах России и обеспечения селекции подсолнечника источниками сопротивления. В случае заразительной расы необходимо использовать не анатомические признаки агрессивности индивидов паразита, а масштабные различия между генотипом и дифференциаторами.
Короткий адрес: https://sciup.org/142171277
IDR: 142171277 | УДК: 633.854.78:582.952.6(470)
Danger of new broomrape races for sunflower in Russia and measures for possible damage prevention
It is stated that overcame due to gene Or3 sunflower resistance broomrape individuals may be considered as plants of race D only after verification of progenies virulence. This fact and theoretical analysis prove, that recommendation to repress virulent races during unrestricted long time by stabilizing selection for rising of race C concentrations in broomrape populations is erroneous. Established situation demands to take prompt actions race status of broomrape populations in Russia regions study and for providing sunflower breeding with resistance sources. For broomrape races identification not anatomical characters of parasite individuals aggressiveness must be used, but international genotype-differentiators scale.
Текст научной статьи Опасность новых рас заразихи для подсолнечника в России и меры предупреждения возможного ущерба
Академик В. С. Пустовойт (1967) считал результативную селекцию подсолнечника на устойчивость к заразихе ( Orobanche сumana Wallr.) важнейшим условием успешного возделывания этой культуры в нашей стране. Поэтому своевременно создавались сорта и линии подсолнечника, устойчивые к последовательно появляющимся расам заразихи А, В и С. Сообщения из разных стран свидетельствуют о том, что процесс образования новых рас этого паразита не только не завершился, но даже интенсифицировался.
A. V. Vrănceanu с сотрудниками (1980) уже в 1978 г. наряду со считавшейся еще новой расой С обнаружили в Румынии еще более вирулентные расы заразихи D и E. Позднее там же была выявлена также раса F (Шкорич, Йоцич, 2006). На юге Испании, в Андалузии, при обследовании посевов подсолнечника в 1986-1988 гг. было установлено, что от 9 до 89 % растений гибридов кондитерского типа были поражены со средней степенью от 0,2 до 35 заразих на растение, а масличного типа – 10- 30 % при средней степени менее 5 заразих на растение (Castejon et al., 1991). Однако уже при этих обследованиях на 19-ти полях масличного подсолнечника в зоне между El Coronil и Montellano было поражено 39 % растений при средней степени 5-6 заразих на растение.
При изучении популяций заразихи, собранных на посевах подсолнечника в Испании, была обнаружена необычная раса, которая поражала сорта-дифференциаторы с генами Or1, Or3 и Or4, но устойчивость к которой обеспечивали гены Or5 и Or2 (Melero-Vara et al., 1989). Дальнейшими исследованиями кроме рас C, D и E была обнаружена еще и раса F заразихи в южной и центральной Испании и показано, что в течение 10 лет в популяциях заразихи снижалась частота расы А и повышались на юге частоты рас E и F, а в центральной Испании – рас B и F (Melero-Vara et al., 2000). По мнению Т. С. Антоновой (1999а, с. 17), существует опасность появления и в России заразихи более вирулентной, чем раса С, так как в ее опытах изолят «Mitra» этого паразита, полученный из южной части Испании, поразил почти все лучшие по устойчивости линии подсолнечника ВНИИМК на 100 %. Полученный также из Испании изолят заразихи IN 23 с геном вирулентности F поразил на 100 % нашу линию ВК 653, устойчивую к расе С (Антонова, 2003). В центральной Испании выявлена еще и раса G заразихи; в Турции обнаружены популяции этого патогена, превосходящие по вирулентности расы А, В, С, D и Е; появились новые его расы в Сербии, Болгарии, Израиле и других странах (Шкорич, Йо-цич, 2006). Последнее время появляются сообщения о значительном поражении заразихой считающихся устойчивыми к ней гибридов в условиях производственных посевов на территории России.
Учитывая ускоренные темпы эволюции подсолнечной заразихи, специалистами ряда стран проводятся комплексные исследования, направленные на разработку мер предупреждения ущерба от возможных эпифитотий. С этой целью идентифицируются новые расы патогена, выявляются доноры устойчивости к этим расам.
В частности, было установлено, что устойчивые к расе В сорта подсолнечника селекции ВНИИМК начали поражаться новой для того времени заразихой с 1964-1966 гг. в Румынии (Vranceanu et al., 1981) и с 1968 г. в Болгарии (Buchvarova, Velkov, 1979). Исследованиями Р. Бъчваровой (1978, 1979) было выявлено 5 типов заразихи в Болгарии и разработана шкала дифференциаторов рас этого паразита, состоящая из трех сортов и одной линии подсолнечника. Проведенными в этой стране с 1975 г. испытаниями методом А. Я. Панченко устойчивости 2469 образцов подсолнечника к поражению этими популяциями было выявлено лишь 2,5 % устойчивых генотипов.
Более детальный анализ устойчивости генотипов подсолнечника к заразихе и вирулентности ее рас провели в Румынии A. V. Vranceanu c сотрудниками (1980, 1981). Они провели скрининг устойчивости среди 1027 сортов, гибридов, инбредных линий разного происхождения, межвидовых гибридов. В результате были выявлены различия генотипов подсолнечника по степени восприимчивости к поражению заразихой и выявили расы этого патогена, превосходящие по вирулентности не только расу В, но и расу С, идентифицированные как расы D и Е. Были найдены также генотипы подсолнечника, устойчивые не только к расам С и D, но и к поражению расой Е заразихи.
Этот комплекс проведенных в Румынии исследований позволил авторам создать шкалу генотипов подсолнечника – дифференциаторов рас заразихи (Vrănceanu et al., 1980, 1981). Генетическим анализом они выявили пять независимых доминантных генов Or1, Or2, Or3, Or4, Or5, определяющих устойчивость подсолнечника к расам А, В, С, D и Е заразихи. В шкалу дифференциаторов эти исследователи включили:
― линию АД-66, восприимчивую ко всем расам заразихи;
― сорт Круглик А-41 с геном Or 1 устойчивости к расе А;
― сорт Ждановский 8281 с геном Or2, устойчивый к расам А и В;
― сорт Рекорд с геном Or3, как дифференциатор расы С;
― линию S-1358 (а также О-7586) с геном Or 4 , устойчивую к расе D;
― линию Р-1380-2 с геном Or5, являющуюся дифференциатором самой вирулентной в то время расы Е. Позже, после появления в Румынии еще более вирулентной расы F, из сорта подсолнечника ВНИИМК была выделена устойчивая к ней линия LC-1093 с геном Or 6 , также включенная в шкалу дифференциаторов рас заразихи (Шкорич, Йоцич, 2006).
Этот набор сортов и линий подсолнечника – дифференциаторов рас заразихи признан основой международной шкалы, используемой в разных странах при изучении локальных популяций подсолнечной заразихи (Melero-Vara et al., 1989, 2000; Шкорич, Йоцич, 2006). С использованием этой шкалы, в частности, было обнаружено появление в Испании особо вирулентной расы G заразихи. Хотя в результате широких испытаний устойчивости родительских линий гибридов подсолнечника в разных странах в ряде случаев были выявлены неожиданные результаты, важность использования этой шкалы не подвергается сомнению. Требуется только ее совершенствование, повышение точности изучения расового состава популяций заразихи с применением новых методов, в том числе молекулярных маркеров (Melero-Vara et al., 2000; Шкорич, Йоцич, 2006).
На основе изучения генетического контроля расоспецифической устойчивости подсолнечника к заразихе А. V. Vrănceanu с сотрудниками (1981) пришли к выводу, что независимые доминантные гены устойчивости Or4 и Or5 линий подсолнечника S-1358, O-7586, P-1380-2 можно легко передавать возвратными скрещиваниями родительским линиям коммерческих гибридов. В результате селекционной работы в этом направлении в Румынии были созданы устойчивые к расе D гибриды Феликс, Се-лект, Ромсун 59, Флором и устойчивые к расе Е заразихи гибриды подсолнечника Фундуля 53 и Сорем 80 (Vranceanu et al., 1987).
На основе опыта своей работы румынские исследователи пришли к выводу о достаточно высокой частоте встречаемости в сортах и коллекциях подсолнечника генов устойчивости к возникающим новым расам заразихи (Vranceanu et al., 1981). Поиски источников таких генов широко ведутся также в популяциях, полученных в результате межвидовой гибридизации подсолнечника (Melero-Vara et al., 2000; Шкорич, Йоцич, 2006).
В 1963 г. в Молдавской ССР была обнаружена раса С подсолнечной заразихи. Она широко распространилась на юге России и в 70-е годы в составе популяций этого патогенна в этом регионе преобладала раса С. Такой их состав сохранялся стабильным в течение десятилетий. Однако, по мнению В. В. Толмачева (1984, 1990), единичные особи этих популяций заразихи, преодолевающие обусловленную геном Or 3 устойчивость подсолнечника, являлись 5 %-ной примесью биотипа наиболее вирулентной расы D в популяции расы С. Поэтому он логично заключил, что на посевах устойчивых к расе С сортов и гибридов подсолнечника происходит интенсивный естественный отбор в пользу расы D заразихи, содержание которой в популяциях этого паразита по предположению В. В. Толмачева (1990, 1991) должно было к 1990 г. достигать 50 %. И хотя единичные особи заразихи преодолевали обусловленную геном Or3 устойчивость подсолнечника уже более четверти века, эпифитотий расы D все это время не наблюдалось.
Для объяснения этого противоречия была предложена гипотеза о том, что численность особей расы D в популяции заразихи расы С ограничивается в силу действия закона стабилизирующего отбора в популяциях паразита. По этому закону менее вирулентная раса С должна подавлять расу D. Вследствие этого более вирулентная раса D при определенных условиях не будет иметь в России серьезного экономического значения. Чтобы обеспечить эти условия, надо способствовать воспроизводству семян расы С, чтобы она неограниченно долго господствовала в российских популяциях заразихи, тогда более вирулентная раса D будет в течение этого времени пребывать в подавленном состоянии. В свете таких представлений о стабильности расового состава заразихи казались преувеличенными сообщения о быстрой эволюции ее рас в европейских странах. Поэтому было высказано мнение, что в Испании комплекс рас заразихи В ошибочно называют расами С и D, расу С – расой Е, а расу D – расой F. Если бы это соответствовало действительности, то завезенной оттуда могла бы быть только раса D, которая уже имеется в России, но подавляется здесь стабилизирующим отбором.
Кроме двух описанных подходов к предотвращению ущерба от появления новых рас заразихи (селекция на устойчивость к ним и подавления стабилизирующим отбором) предложен и третий способ: подавление или уничтожение заразихи обработкой полей подсолнечника гербицидами. Раньше было предложено внесение трефлана под предпосевную культивацию, что обеспечивало уменьшение числа цветоносов заразихи в 3,5-5 раз
(Васильев, Баранова, 1974; Ačimovič, 1980). В настоящее время создаются гибриды подсолнечника, устойчивые к группе гербицидов имидазолинона, при обработке посевов которых такими гербицидами уничтожаются не только сорняки, но и популяции любых рас заразихи (Шкорич, Йоцич, 2006). Ниже мы рассмотрим только обоснованность представления о том, что в условиях России предотвратить потери от появления новых вирулентных рас заразихи можно не селекцией на устойчивость к ним, а поддержанием стабилизирующего отбора в пользу расы С заразихи.
Представление о возможности защиты подсолнечника от новых рас заразихи стабилизирующим отбором в ее популяциях возникло на основе факта длительной стабильности расового состава российских популяций заразихи и попытке объяснения этого факта действием закона стабилизирующего отбора в популяциях патогенна, ссылаясь при этом на книги Ван дер Планка (1972). Однако эта ссылка не совсем точна. Этот автор под стабилизирующим отбором понимает не отбор в пользу менее вирулентной расы, а то, что при отсутствии гена устойчивости «… вирулентность, необходимая для его подавления, оказывается ненужной и стабилизирующий отбор действует в пользу тех рас патогенна, которые не имеют этой ненужной вирулентности» (Ван дер Планк, 1972, с. 75). Сам автор признает, что это гипотеза, с которой не все согласны (Вандерпланк, 1981, с. 54, с. 143). Имеются факты, противоречащие этой гипотезе (Parlevliet, 1981; Расселл, 1982, с. 97).
В одной из своих книг Ван дер Планк (1972, с. 94) пишет: «Основной темой настоящей главы является сила, которую называли различно – генетический гомеостаз, стабилизирующий отбор или нормализирующий отбор». О том, что между этими понятиями нельзя ставить знак равенства, свидетельствует сравнение их определений. «Отбор, благоприятствующий сохранению особей, количественные признаки которых близки к среднему значению или равны ему, называется стабилизирующим отбором» (Меттлер, Грегг, 1972, с. 172). «Гомеостаз. 1. Явление, при котором частота гена в популяции, после ее нарушения в результате изменения условий среды или искусственного отбора, снова достигает прежнего уровня под влиянием естественного отбора. Лернер (1950) назвал этот тип гомеостаза генетическим гомеостазом или гомеостазом частоты гена» » (Ригер, Михаэлис, 1967, с. 118). Определение Я. Вандерпланка (1981, с. 142): «…стабилизирующий отбор означает, что частота аллеля авирулентности в соответствующем локусе патогена выше частоты аллеля вирулентности» ― явно подразумевает гомеостаз частоты гена, а не отбор особей по фенотипическим количественным признакам. Это подчеркивает и J. E. Parlevliet (1981).
О том, что Ван дер Планк под стабилизирующим отбором понимает только изменения частот генов вирулентности и авирулентности, свидетельствует использование им этой концепции для объяснения длительной вертикальной устойчивости к расам стеблевой ржавчины некоторых сортов яровой пшеницы в Канаде. В летний период на растениях таких сортов при адаптации популяций патогена к генам устойчивости сортов происходит направленный отбор на повышение частот генов вирулентности, концентрация которых затем снижается в течение осени, зимы и весны вследствие перезимовки патогена на юге США на неустойчивых сортах озимой пшеницы (Ван дер Планк, 1972, с. 75, 118, 125). Более подробное изучение показало, что важным или даже определяющим условием такой стабильной устойчивости является пренебрежимо малая вероятность комбинирования двух генов вирулентности стеблевой ржавчины, что защищало от поражения сорт Селкирк, обладавший соответствующими двумя генами устойчивости (Вандерпланк, 1981, с. 144-145). R. A. Robinson (1971) также считает, что стабилизирующий отбор может действовать лишь при наличии двух генов вертикальной устойчивости к облигатному паразиту. По Г. Э. Расселлу (1982, с. 96, с. 385), причиной долговременной устойчивости сорта Селкирк была комбинация главных генов в сочетании с генами-модификаторами, а не стабилизирующий отбор.
По аналогии с гипотезой Ван дер Планка была предложена рекомендация «– для подавления накопления расы D в российской популяции заразихи необходимо соблюдать правило наличия двух хозяев для расы С: устойчивого и восприимчивого к ней. В этом случае по закону стабилизирующего отбора раса С будет вытеснять из популяции расу D, и последняя не будет иметь экономического значения». Приемлемость такой аналогии в данном случае сомнительна по ряду причин. В частности, во взаимоотношениях подсолнечника и заразихи не обнаружено описанной Вандерпланком (1981, с. 144-145) устойчивости за счет пренебрежимо малой вероятности комбинирования двух рас патогена, нет и обязательной смены хозяев, расположенных друг от друга на расстоянии тысяч километров. Поскольку в отличие от спор ржавчины основная масса семян заразихи остается на том поле, где были поражены ею растения подсолнечника, для размножения расы С стабилизирующим отбором необходимо, чтобы до высева устойчивого гибрида или сорта на этом поле выращивался сильно поражаемый заразихой С сорт, что неприемлемо для производства. Более того, даже теоретически при выполнении такой рекомендации более вирулентная раса не может быть вытеснена расой С.
Посевы обладающих геном Or3 сортов и гибридов подсолнечника должны были служить селективным фоном для накопления в популяциях заразихи особей более вирулентной расы D. В. В. Толмачев (1990, с. 15) писал: «На посевах устойчивых к расе С сортов и гибридов подсолнечника идет интенсивный естественный отбор и накопление бо- лее вирулентной расы D, содержание которой в отдельных популяциях (Саратовская, Каневская) достигает 50 %». Информация о высокой вирулентности саратовской популяции заразихи оказалась ошибочной.
В особой жизнеспособности заразихи расы С, позволяющей ей угнетать более вирулентную расу D, обеспечивая тем самым стабильное равновесие между этими расами, заставляет усомниться история смен рас заразихи на посевах подсолнечника в нашей стране. Уже к 1934 году была создана серия сортов-популяций подсолнечника с высокой долей особей, устойчивых к расе В и названных «заразиховыносливыми» (Пустовойт, 1967). Выбраковка из таких сортов восприимчивых растений в процессе семеноводства позволила сделать эти сорта полностью устойчивыми к заразихе В (Пустовойт, 1957). Обусловленная геном Or 2 устойчивость к расе В не преодолевалась многие годы, хотя это никак нельзя объяснить стабилизирующим отбором по Ван дер Планку. Уже с 1938 г. биотипов расы А не было в популяциях заразихи.
Причина долговременной устойчивости к расам как В, так и С иная. N. W. Simmonds (1985) показал, что быстрое преодоление новых генов вертикальной устойчивости характерно только для мобильных патогенов, споры которых переносятся по воздуху на большие расстояния, а от малоподвижных паразитов, инфекционное начало которых передается через почву, гены устойчивости значительно более надежно и длительно защищают сорта и гибриды. Более того, мобильные патогены размножаются в ходе эпифитотии в результате ряда следующих друг за другом поколений (Ван дер Планк, 1966, с.41). Это обеспечивает более высокую вертикальную мутабильность по сравнению с вероятностью образования новых вирулентных рас заразихи, которая размножается на конкретном поле не чаще, чем 1 раз в год, если не соблюдается севооборот. Относительно малая мобильность заразихи и намного меньшее число циклов ее размножения по сравнению с возбудителями большинства болезней являются достаточными аргументами для объяснения длительности периодов стабильной устойчивости подсолнечника к этому паразиту. Известно, что мутациями патогена вертикальная устойчивость хозяина преодолевается быстрее, если не происходит полная элиминация менее вирулентной расы (Robinson, 1971). Не исключено, что возникновение ряда рас заразихи в Испании было ускорено вследствие массового возделывания восприимчивых кондитерских гибридов подсолнечника рядом с посевами масличных (Castejon et al., 1991).
Через 10 лет после публикации В. В. Толмачевым (1984) вывода о наличии в каневской популяции заразихи расы С, 5%-ной примеси новой более вирулентной ее расы D и утверждения о накопле-ниидо 50 % этой расы в результате интенсивного естественного отбора (Толмачев, 1990) ни в Каневском районе, ни в Саратовской области не наблюда- лось соответствующего поражения сортов подсолнечника, устойчивых к расе С. Это вызвало сомнения в обоснованности идентификации расы D заразихи. Однако важность проблемы не позволяла игнорировать мнения о появлении такой расы в российских популяциях заразихи. Поэтому перед отделом генетики и физиологии растений ВНИИ масличных культур была поставлена задача найти и передать селекционерам источник гена устойчивости подсолнечника к этой расе. Проведенными исследованиями была установлена ошибочность вывода В. В. Толмачева (1984, 1987, 1990) о том, что единичные растения заразихи, преодолевающие обусловленную геном Or3 устойчивость подсолнечника, обладают наибольшей вирулентностью и являются особями новой расы D заразихи. Полученные экспериментальные данные были изложены в научном отчете отдела за 1995 год, но не были опубликованы. Поэтому и позднее появлялись публикации о наличии в российских популяциях заразихи 5 %-ной примеси расы D (Таволжанский, 2000, с. 312). Поскольку актуальность этой темы резко возросла в последние годы, мы посчитали необходимым опубликовать полученные ранее экспериментальные данные изучения вирулентности особей заразихи, преодолевающих устойчивость подсолнечника, обусловленную геном Or3.
Изучить предполагаемую новую расу заразихи можно было лишь в том случае, если исследовать не сложную популяцию с 5 %-ной примесью семян этой расы, а получить максимально чистые ее семена. Поэтому селективным фоном для размножения особей такой расы должны были служить растения генотипа подсолнечника, устойчивого к расам А, В и С заразихи, причем это должен был быть не сорт-популяция, а самоопыленная линия. С этой целью в пространственно изолированном посеве устойчивой к расе С инбредной линии подсолнечника ВК 639Б собрали семена с растений заразихи, преодолевших барьер устойчивости, обусловленный геном Or3. В качестве контрольных служили семена обычной для того времени популяции заразихи, собранные в посеве неустойчивой инбредной линии ВК 66. Для оценки вирулентности этих популяций заразихи использовали ту же устойчивую к расе С линию ВК 639Б и сорт ВНИИМК 8883, восприимчивый к этой расе паразита. Поражаемость растений этих линий определяли тепличным методом ранней диагностики по А. Я. Панченко (1975) по числу заразих на корневой системе одного растения подсолнечника на 25-й день после появления всходов. Полученные результаты представлены в таблице 1.
Оказалось, что воспроизводство семян заразихи на растениях устойчивой к расе С линии подсолнечника ВК 639Б не повысило степень вирулентности популяции паразита по отношению к этой линии как по проценту пораженных растений, так и по числу заразих в среднем на одно растение. Из этого следует, что обусловленная геном Or3 устойчивость растений подсолнечника преодолевается отдельными особями заразихи изученной ее популяции не вследствие появления в ней растений с новым, более вирулентным геном. На растениях устойчивой линии ВК 639Б размножались особи заразихи расы С вследствие преодоления не полной устойчивости к ней подсолнечника, обладающего геном Or3. В сельскохозяйственных патосистемах явление частичной устойчивости встречается не редко, ее действие может варьировать от незначительного проявления до полного подавления развития паразита (Попкова, 1979. с. 189; Вандерпланк, 1981, с. 163-164).
Таблица 1 – Оценка вирулентности популяций заразихи, собранных в посевах линий подсолнечника, устойчивой (ВК 639Б) и восприимчивой (ВК66) к расе С паразита
|
Заразиха из посева линии |
Инфицированный генотип подсолнечника |
Число растений подсолнечника |
Процент пораженных растений |
Заразих на 1 растение подсолнечника |
||
|
линия, сорт |
ген устой- чивости |
всего |
в т. ч. пораженных |
|||
|
ВК 639Б |
ВК 639Б |
Or 3 |
90 |
7 |
7,8 |
1,0 |
|
ВК 639Б |
ВНИИМК 8883 |
Or 2 |
12 |
12 |
100 |
Более 10 |
|
ВК 66 |
ВК 639Б |
Or 3 |
35 |
4 |
11,4 |
1,0 |
|
ВК 66 |
ВНИИМК 8883 |
Or 2 |
10 |
10 |
100 |
Более 10 |
В связи с полученным результатом представляет интерес анализ эксперимента, на основании которого В. В. Толмачев (1984) сделал вывод не только о наличии расы D, но и о том, что доля ее в каневской популяции составляет 5 %. Этот опыт проводился не для анализа популяции заразихи, а с целью определения соотношения частот устойчивых и восприимчивых растений к расе С заразихи в потомствах от самоопыления гетерозиготных по признаку заразихоустойчивости растений линии подсолнечника ВК 373Б. Оценка проводилась по методу А. Я. Панченко (1975) при одинаковой инфекционной нагрузке для всех семей сибов 0,2 г семян заразихи на 1 кг почвенного субстрата. К классу «устойчивые» относились особи подсолнечника не только полностью непораженные, но и с единичными растениями заразихи на корнях. На основании этого опыта не только был сделан вывод о моногенном контроле признака устойчивости, но еще и по данным шести семей потомков от самоопыления гетерозиготных родителей были проведены корреляционный и регрессионный анализы. При этом выявилась положительная зависимость между средним числом заразих на корнях восприимчивых растений и на корнях устойчивых растений той же семьи полных сибов при r=0,88 и коэффициенте регрессии b=0,047. При интерпретации этих данных автор принял без доказательств два сомнительных тезиса:
-
1. Особи заразихи, преодолевающие устойчивость, обусловленную геном Or 3 , относятся к более
-
2. Модификационная изменчивость степени поражения вызвана неодинаковой инфекционной нагрузкой для разных семей полных сибов.
вирулентной расе.
В. В. Толмачев должен был знать, что опытные селекционеры, изучавшие заразихоустойчивость подсолнечника, неоднократно писали о поражении отдельными особями паразита устойчивых растений подсолнечника со средней степенью от 1 до 3 и более заразих на одну корневую систему, однако не делали на этом основании вывода о появлении более вирулентной расы (Пустовойт, 1963; Жданов, 1964; Г. В. Пустовойт, Крохин, 1978). M. Acimovic
(1980) относил к числу устойчивых растения подсолнечника с единичными особями заразихи на корнях. Однако В. В. Толмачев (1984) без проверки вирулентности объявил представителями новой расы заразихи единичные ее особи, преодолевшие устойчивость к расе С. В других же его опытах обусловленную геном Or3 устойчивость заразиха расы С преодолевала со средней степенью до 13,4 особей на 1 растение подсолнечника, однако интерпретировалось это как неполная экспрессивность гена Or3, то есть не полная устойчивость к расе С, а не как появление новой расы (Толмачев, 1987, 1990). А поскольку большие различия по степени поражения устойчивых растений в этих опытах были обусловлены введением гена Or3 в различные генотипические среды, а не разным составом популяций заразихи, этими результатами В. В. Толмачев сам доказал ошибочность своего вывода о том, что уже к 1984 году в российских популяциях заразихи расы С имелось 5 % особей более вирулентной расы D.
При обсуждении результатов опыта, на основании которого В. В. Толмачев (1984) сделал вывод о появлении в популяциях заразихи расы С примеси особей расы D, он также бездоказательно утверждал, что различия между семьями потомств само-опыленных растений подсолнечника по степени поражения заразихой были обусловлены неодинаковой инфекционной нагрузкой. Однако из описания условий проведения этого опыта следует, что для всех семей создавалась одинаковая нагрузка 0,2 г семян заразихи на 1 кг почвенного субстрата. Поэтому возможная очень небольшая неравномерность распределения семян заразихи при перемешивании ее с почвенно-песчаной смесью по методике А. Я. Панченко (1975) не может быть причиной различий между семьями по степени поражения от 40 до 140 заразих на одно растение подсолнечника. Более вероятной причиной таких различий может быть неодинаковая степень инбредной депрессии растений разных семей вследствие самоопыления их матерей, так как В. В. Толмачев (1990, с. 7) продемонстрировал необходимость вносить поправки на «… генотипически обусловленные различия в степени развития корневой системы, которые определяют фенотипические различия в степени поражения». Однако в данном опыте автор не произвел такую коррекцию, поэтому в итоге произошла экстраполяция на все российские популяции заразихи сомнительного вывода о содержании в них пяти процентов расы D.
Для того чтобы действительно обеспечить разные уровни инфекционной нагрузки и определить их влияние на степень поражения заразихой растений конкретного генотипа подсолнечника также в 1995 г. был проведен опыт с целью сравнительного изучения реакций на увеличенный диапазон таких уровней устойчивой к расе С инбредной линии ВК 464А и восприимчивого к ней сорта ВНИИМК 8883. Результаты этого опыта, представленные в таблице 2, также были изложены в научном отчете отдела генетики и физиологии ВНИИМК за 1995 г., но не были опубликованы.
личение инфекционной нагрузки устойчивая к расе С линия ВК 464А (см. табл. 2). При дозе 0,2 г семян заразихи на 1 кг почвы, при которой из-за неравномерности распределения их в почве имеет значение вероятность встречи корней с патогеном (Дьяков, Антонова, 1978), процент пораженных растений линии ВК 464А оказался ниже, чем при 0,4 и 0,8 г/кг. Эти результаты прямого опыта не подтвердили основанное на косвенных данных предположение В. В. Толмачева (1984) о высокой корреляции между уровнем инфекционной нагрузки и степенью поражения заразихой растений устойчивого к расе С генотипа. Среднее число заразих на одно пораженное растение линии ВК 464А варьировало от 1,38 до 2,62 независимо от доз семян патогена. Вследствие разного характера изменения этого показателя у растений сорта ВНИИМК 8883 и линии ВК 464А на градиенте инфекционных нагрузок закономерно в 4,8 раза изменяется величина степени поражения устойчивого генотипа в процентах к
Таблица 2 – Влияние количества семян заразихи в почвенном субстрате на поражение растений устойчивой к расе С линии подсолнечника ВК 464А и восприимчивого к ней сорта ВНИИМК 8883
|
Семян заразихи, г/кг почвы |
Поражено растений, % |
Среднее число заразих на 1 пораженное растение |
|||
|
сорта ВНИИМК 8883 |
линии ВК 464А |
сорта ВНИИМК 8883 |
Линии ВК 464А |
||
|
заразих на 1 растение |
в % к сорту ВНИИМК 8883 |
||||
|
0,0 |
8,3 |
0 |
1,0 |
0 |
- |
|
0,2 |
100 |
8,9 |
29,0 |
1,66 |
5,7 |
|
0,4 |
100 |
35,4 |
57,9 |
1,46 |
2,5 |
|
0,8 |
100 |
32,0 |
54,8 |
1,64 |
3,0 |
|
1,6 |
92,3 |
20,2 |
22,1 |
2,62 |
11,9 |
|
3,2 |
93,3 |
14,0 |
12,1 |
1,38 |
11,4 |
этому показателю восприимчивого. А именно сохранение стабильности этой величины на уровне около 5 % при широком диапазоне модификационной изменчивости степени поражения В. В. Толмачев (1984) считал критерием наличия новой расы заразихи и ее доли в популяции расы С. О неправомочности таких выводов свидетельствуют как значительная вариабельность этого показателя, так и относительно небольшая изменчивость среднего числа заразих на корнях устойчивых растений на градиенте доз семян заразихи в почве.
Вывод В. В. Толмачева (1984) о
Характер зависимости показателей поражения растений восприимчивого сорта подсолнечника ВНИИМК 8883 от содержания семян заразихи в почвенной смеси оказался типичным для оценок по методу А. Я. Панченко (1975) при учетах по клубенькам паразита на корнях подсолнечника (Дьяков, Антонова, 1978). Внесения 0,2 г всхожих семян заразихи на 1 кг почвенной смеси достаточно для обеспечения поражения всех восприимчивых растений, но число клубеньков заразихи на их корнях еще возрастает прямо пропорционально увеличению инфекционной нагрузки до 0,4 г/кг; в диапазоне от 0,4 г/кг до 0,6 г/кг достигается насыщение корневых систем паразитом вследствие ограниченности ресурсов хозяина. Как и в ранее неоднократно проведенных опытах, при внесении на 1 кг почвы более 1 г семян заразихи уменьшается процент пораженных растений и, особенно, число заразих на их корневых системах, что, по данным А. Я. Панченко, объясняется массовым поражением фузарио-зом проростков заразихи при большом их количестве в единице объема почвы.
Несколько иначе реагировала на такое же уве- том, что в широком диапазоне инфекционных нагрузок преодолевают моногенно обусловленную устойчивость 5 % особей каневской популяции заразихи был сделан на основе уравнения y=0,4+0,047x, где у и х – показатели степени поражения соответственно устойчивого и восприимчивого аналогов линии подсолнечника ВК 373Б. Однако рассмотрение этого уравнения свидетельствует об ошибочности этого вывода. Соответствующая этой формуле линия регрессии проходит выше начала координат, поэтому оценки доли преодолевших устойчивость особей заразихи не остаются постоянными, а возрастают по мере уменьшения степени поражения восприимчивых растений. Вычисления по этому уравнению показывают, что 5 % особей заразихи поражают устойчивые растения только при степени поражения восприимчивых 140 заразих на одно растение. Если же восприимчивые поражены со степенью 10 заразих на растение, то процент особей паразита, преодолевающих устойчивость, достигает 100(0,4+0,047∙10) 10=8,7 %, а при степени 7 заразих – 10,4 %.
При достаточных инфекционных нагрузках час- тоты преодоления устойчивости растений подсолнечника отдельными особями авирулентной для них расы патогена должны подчиняться распределению случайных событий. Для того чтобы проверить, в какой мере фактическая изменчивость степени поражения заразихой расы С растений устойчивой к ней линии ВК 464А соответствует вероятностному распределению, были изучены частоты растений этой линии с разным числом заразих. Для такого анализа использовали данные о растениях линии ВК 464А вариантов 0,4 и 0,8 г семян заразихи на 1 кг почвенной смеси опыта 1995 г., средние данные которого представлены в таблице 2. На графике (рисунок) эмпирический вариационный ряд частот растений с разным числом преодолевших устойчивость подсолнечника линии ВК 464А заразих представлен резко асимметричной односкатной кривой. Такая кривая характерна для распределения редких событий, называемого распределением Пуассона. Вычисление и нанесение на график соответствующих теоретических частот показало хорошую согласованность с ними фактических данных, что свидетельствует о подчинении эмпирического ряда закону редких событий Пуассона. А это означает, что при высоких инфекционных нагрузках с небольшой вероятностью возникают сочетания обстоятельств, позволяющие отдельным особям заразихи расы С преодолевать устойчивость подсолнечника, обусловленную геном Or3.
Ошибочность сделанного в 1984 г. В. В. Толмачевым вывода о принадлежности к расе D отдельных особей популяции молдавской расы заразихи, поражающих устойчивые к расе С растения подсолнечника, ставит под сомнение и справедливость гипотезы о стабилизирующем отборе в пользу расы С в популяциях заразихи. О том, почему возникла такая гипотеза, можно судить по таким цитатам: «Еще в середине семидесятых мы наблюдали на корнях форм подсолнечника, проявлявших защитную реакцию в сосудах против биотипа С, иногда рядом с обилием погибших проростков заразихи соседствовал нормально развившийся клубенек …
Очевидно, что это были представители более вирулентного биотипа, чем биотип С. Поэтому, по нашим наблюдениям, представители биотипа D присутствуют в российских популяциях заразихи по крайней мере на протяжении двух последних десятилетий. За этот период не возникло эпифито-тий».
Отметим еще раз, что преодоление устойчивости отдельными особями патогена нельзя считать очевидным и достаточным доказательством отнесения этой особи к более вирулентной расе. Важно, что автор видит противоречие между представлением о появлении такой расы и отсутствием в течение десятилетий эпифитотий. Для объяснения этого противоречия была выдвинута ни чем другим не обоснованная (как было показано выше) гипотеза: «… в популяциях паразитов имеет место стабили-
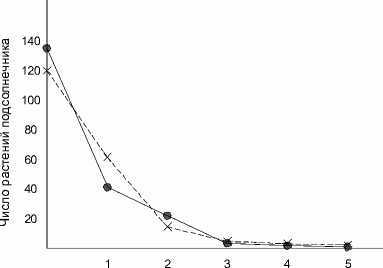
Число заразих на растении подсолнечника
Рисунок - Фактические (х) и теоретические (•) частоты распределения Пуассона преодоления устойчивости растений линии подсолнечника ВК 464А заразихой расы С зирующий отбор менее вирулентных биотипов, как более жизнеспособных и сохраняющихся на растениях-хозяевах, не имеющих генов устойчивости … Сложившийся баланс между биотипами D и C с преобладанием последнего в российских популяциях заразихи соответствует теории стабилизирующего отбора». А далее из этого делается практический вывод: «По закону стабилизирующего отбора внутри популяции патогенна будет преобладать менее вирулентная раса, если в популяции хозяина одновременно присутствуют устойчивый и неустойчивый к этой расе сорта (Ван дер Планк, 1972). … Мы полагаем, что при сложившихся условиях в настоящее время раса D в России не имеет серьезного экономического значения». (Заметим, что присутствие разных по устойчивости сортов в одной популяции несовместимо с гипотезой Ван дер Планка).
Вывод о малом ущербе от заразихи расы D усугубляется представлением о путанице в Западной Европе в обозначении рас заразихи, о том, что расами С и D там обозначают комплекс рас В, раса Е это раса С, а раса F соответствует нашей расе D. В сочетании же с ранее приведенными цитатами это означает, что нам в России нечего опасаться: ничего более вирулентного, чем раса D в Европе нет, а расу D мы можем подавить стабилизирующим отбором настолько, что она неограниченно долго будет пребывать в подавленном состоянии, не представляя для нас серьезного экономического значения.
Описанным выше анализом показана несостоятельность гипотез, на которых основана эта практическая рекомендация. Следовательно, появление настоящих более вирулентных рас заразихи нельзя будет нейтрализовать предложенным способом, даже если бы была практически реальной возможность на 50 % всех посевов подсолнечника в стране выращивать его сорта, сильно поражающиеся расой
С паразита. Более того, уверенность в том, что при выполнении этой рекомендации можно будет неограниченно долго подавлять вирулентные расы паразита, уже негативно сказалась на текущих исследованиях в этой области: ВНИИМК даже не имеет набора генотипов-дифференциаторов рас заразихи, не контролирует ее расовый состав в регионах страны, не ведет поиски генотипов, устойчивых к новым расам.
Возникает опасение, что и в случае проведения таких исследований будут допускаться те же ошибки, которые привели к утверждению о путанице в определении рас заразихи в европейских странах. Поэтому важно сравнить методические подходы к дифференциации рас заразихи.
Для выявления физиологических рас биотроф-ных патогенов во всем мире используется единый принцип. Ю. Т. Дьяков (1971) описывает его так: «Определение рас фитопатогенных организмов проводится на сортах-дифференциаторах. Чтобы иметь сопоставимые данные по распространению физиологических рас в различных районах выращивания восприимчивой культуры, определение рас проводится на международных наборах сортов-дифференциаторов». В. С. Пустовойт (1967) предложил для распознания рас заразихи А и В сорта подсолнечника, которые он назвал индикаторами. Один из этих сортов Круглик А-41 A.V. Vranceanu с соавторами (1981) включил в набор генотипов подсолнечника – дифференциаторов рас заразихи. Этот набор, включавший вначале дифференциаторы с доминантными генами устойчивости Or1, Or2, Or3, Or4 и Or5, был принят в качестве международного и дополнялся по мере появления новых рас заразихи и выявления соответствующих линий – источников комплементарных генов устойчивости (Melero-Vara et al., 1989, 2000; Шкорич, Йоцич, 2006).
Вскоре после появления в Краснодарском крае молдавской заразихи (расы С) предварительный анализ местной популяции этого патогенна показал, что число особей заразихи, преодолевающих обусловленную геном Or 2 устойчивость сорта подсолнечника Передовик, составляло от 44 до 50 % от числа заразих на корнях сорта Круглик А-41, имеющего ген устойчивости Or1 (Дьяков, Антонова, 1978). Это даже больше, чем 39,6 % таких особей, обнаруженных в 1978 г. в популяции из Mircea-Voda в Румынии на дифференциаторах Круглик А-41 и Ждановский 8281 (Vranceanu et al., 1981).
При изучении особенностей реакций растений сортов подсолнечника Круглик А-41 и Передовик на градиенты доз семян заразихи было обнаружено, что различия между этими сортами можно установить не только по числу развившихся особей патогена, но и по числу некротических участков на корнях, образовавшихся в местах гибели проростков заразихи (Дьяков, Антонова, 1978). Число таких некрозов было больше на корнях тех растений, ко- торые в меньшей мере поражались заразихой. Проводившимися в тот же период Т. С. Антоновой детальными исследованиями были выявлены различные типы анатомических картин защитных реакций растения-хозяина и гибели паразита. Однако при интерпретации этих данных нами (Дьяков, Антонова, 1978) была допущена ошибка. Мы решили, что до того времени, когда будет в полном объеме изучена генетика взаимоотношений заразихи и подсолнечника, судить о вирулентности рас паразита можно по анатомическому типу защитных реакций хозяина подобно тому, как для этого оказалось эффективным использовать некрозы на корнях подсолнечника.
В результате большого объема анатомических исследований с использованием гистохимических методов Т. С. Антонова получила чрезвычайно важные приоритетные результаты, позволившие детально описать механизмы защиты подсолнечника от поражения заразихой. В частности, она установила, что соседствующие на одном и том же корне проростки различных особей одной популяции заразихи могут погибать как в разных слоях коровой паренхимы, так и по достижению ксилемы корня. Эти различия по агрессивности особей заразихи были интерпретированы как их принадлежность к разным по вирулентности расам. Этот вывод в принципе невозможно проверить путем испытания потомств в связи с гибелью проростков, поэтому такие умозаключения остаются гипотетическими. Однако известно, что признаки, определяющие характер развития поражения, являются показателями не вирулентности, а агрессивности (Попкова, 1979, с. 137). А как показал Я. Ван дер Планк (1972, с. 30), различия по агрессивности особей патогена не могут свидетельствовать об их разной вирулентности; корреляция между этими свойствами обычно даже отрицательна. В мировой литературе по этой проблеме общепризнано, что единственный признак, объединяющий разные по агрессивности и другим свойствам генотипы био-трофного паразита в одну физиологическую расу, ― это ее специфическая вирулентность, то есть унаследованная способность поражать конкретные генотипы растения-хозяина (Расселл, 1982, с. 30). Поэтому единственным и общепринятым способом идентификации физиологических рас патогенов, то есть установления их вирулентности, является определение специфических реакций, которые они вызывают при заражении растений сортов-дифференциаторов (Ю. Т. Дьяков, 1967; Попкова, 1979, с. 106-107; Вандерпланк, 1981, с. 201).
Поскольку растение должно быть устойчивым к множеству биотрофных патогенов с учетом большого числа рас каждого из них, реакции отторжения паразитов не могут быть расоспецифичными. Поэтому функцией гена такой устойчивости является «узнавание» проникшего в ткани хозяина паразита соответствующей расы и инициирование в результате этого процессов неспецифичной защит- ной реакции растения-хозяина (Ю. Т. Дьяков, 1971; Щербаков, 1976; Вандерпланк, 1981, с. 64). Это «узнавание» обеспечивается комплементарным взаимодействием специфических белков – первичных продуктов активности гена вирулентности паразита и гена устойчивости хозяина (Ю. Т. Дьяков, 1971; Щербаков, 1976; Вандерпланк, 1981, с. 202). С этими представлениями совершенно несовместим вывод о том, что ген устойчивости подсолнечника к расе С заразихи контролирует биосинтез конифери-лового спирта, а соответствующий ген вирулентности расы С заразихи – пероксидазу. Такого типа реакции должны контролироваться полигенно, если биосинтез хотя бы одного из участников реакции является многоступенчатым, а полигенная устойчивость не может быть расоспецифической (Щербаков, 1976; Вандерпланк, 1981, с. 202).
В качестве примера нелогичных, противоречащих фактам и фундаментальным теоретическим принципам рассуждений приведем цитату о расе В заразихи: «Против биотипа В у подсолнечника действует неспецифическая раневая реакция в коровых клетках корня. Она состоит из цепи биохимических превращений фенольных соединений в поврежденных клетках. Образующиеся при этом многочисленные биохимические соединения, каждое само по себе, могут быть токсичны для заразихи. Потеря чувствительности к ним создает у заразихи целый спектр мало значимых рас. Это и объясняет разнородность биотипа В, который в свое время был назван комплексом рас В. И, как известно, устойчивость к этому биотипу имеет полигенный контроль». Не говоря уже о недопустимости суждений о расовом составе популяции паразита по анатомическим особенностям патологического процесса и о принципиальной недоказуемости принадлежности к расе В погибших в коровой паренхиме проростков заразихи, цитированные рассуждения не выдерживают критики и по ряду других моментов. Совершенно нелогично устойчивость к конкретной расе считать расонеспецифической. Полностью гипотетическим, не доказанным является представление о том, что в пределах комплекса рас В заразихи сформировался спектр рас, соответствующих многочисленным токсичным биохимическим соединениям. Без ссылки на источник непонятно, откуда взяты сведения о полигенном контроле устойчивости подсолнечника к расе С, поскольку не только доказано, что обусловленная геном Or2 устойчивость подсолнечника к этой расе контролируется одним независимым доминантным геном, но и обладающий им сорт Ждановский 8281 включен в международную шкалу дифференциаторов (Г. В. Пустовойт, Крохин, 1978; Vranceanu et al., 1980, 1981; Шкорич, Йоцич, 2006). Устойчивость к отдельной расе в принципе не может быть полиген-ной, так как такой генетический контроль обусловливает только горизонтальную, расонеспецифическую устойчивость (Щербаков, 1976; Ван-дерпланк, 1981, с. 180, 202).
Такая же оригинальность взглядов на генетику взаимоотношений паразита и хозяина обнаруживается и при знакомстве с изложением результатов ее анатомических исследований течения инфекционных процессов, вызываемых патогенными для подсолнечника грибами. Например, об этом свидетельствует такой вывод: «Разнообразие расового состава плазмопары формировалось под воздействием токсичных продуктов раневой реакции клеток корня и гипокотилей проростков. Поэтому различия между расами состоят в адаптационных механизмах ферментативного комплекса выделений паразита к токсичным продуктам раневой реакции».
От надежности результатов фитопатологического и генетического изучения устойчивости селектируемого растения к вредоносному для него паразиту зависит своевременность обнаружения новых вирулентных рас патогена, объективность прогноза распространения этих рас, научная обоснованность рекомендуемых мер предупреждения возможного ущерба от эпифитотий. По всем этим крайне важным вопросам описанные представления совершенно не согласуются с результатами исследований зарубежных специалистов и с принимаемыми ими практическими мерами по созданию заразихоустойчивых гибридов подсолнечника. В основе этих противоречий лежит использование разных принципов идентификации рас заразихи.
Для решения о том, чьим выводам о расах заразихи можно доверять ясно и без описанного выше анализа используемых методов идентификации рас. В случаях таких разногласий правда оказывается на стороне сообщества ученых, пользующихся обоснованной, общепринятой методикой, а не одного исследователя, использующего сомнительную методику.
Таким образом, никаким стабилизирующим отбором невозможно предотвратить распространение вирулентных рас заразихи и вызываемые ими эпи-фитотии, если такие расы появятся или уже появились в России. Поэтому необходимо принять срочные меры для изучения общепринятым методом расового состава заразихи на пораженных ею посевах подсолнечника в разных регионах страны и обеспечить селекцию на заразихоустойчивость выявлением соответствующих источников устойчивости.
