Определение модельных констант при вычислении температуры плавления ДНК

Автор: Курочкин В.Е., Белов Дмитрий Анатольевич, Белов Ю.В., Зубик А.Н.
Журнал: Научное приборостроение @nauchnoe-priborostroenie
Рубрика: Приборостроение физико-химической биологии
Статья в выпуске: 2 т.30, 2020 года.
Бесплатный доступ
В настоящей статье описаны факторы, влияющие на величину определяемой температуры плавления ДНК T m, оценена степень их влияния. Выполнен обзор существующих моделей для вычисления температуры плавления ДНК, выявлено значительное расхождение результатов применения моделей и экспериментальных данных. Предложено применение упрощенных моделей, учитывающих зависимость температуры T m от некоторых факторов при неизменных иных факторах. В качестве примера приведена упрощенная модель, учитывающая зависимость от длины фрагмента ДНК и содержания GC-пар нуклеотидов. Описана методика определения модельных констант для вычисления температуры плавления ДНК на основе анализа результатов экспериментов и данных из известных литературных источников. Отклонение температуры плавления T m при использовании модели, уточненной по экспериментальным данным, не превышает 0.29 °C, значение СКО составляет 0.13 °C.
Анализатор нуклеиновых кислот, пцр в реальном времени, температура плавления днк
Короткий адрес: https://sciup.org/142223740
IDR: 142223740 | УДК: 543.426.1, | DOI: 10.18358/np-30-2-i1016
Model constants determination in the DNA melting temperature calculating
The factors affecting the value of the DNA melting temperature T m are described and the degree of their influence is estimated in the article. A review of existing models for calculating the DNA melting temperature is carried out, a significant discrepancy between the results of model application and experimental data is revealed. It’s proposed to use simplified models that take into account the dependence of the temperature T m on some factors with other factors unchanged. A simplified model is given as an example, taking into account the dependence on the length of the DNA fragment and the content of GC-pairs of nucleotides. A method for determining model constants for calculating the DNA melting temperature based on the experimental results analysis and data from the known literature is described. The error in calculating the melting temperature T m when using the model adjusted according to experimental data does not exceed 0.29 °C; the standard deviation is 0.13 °C.
Текст научной статьи Определение модельных констант при вычислении температуры плавления ДНК
Известно, что генетическая информация закодирована в ДНК в виде двух полимерных цепей, соединенных водородными связями. Цепи образуют двойную спираль, содержащую азотистые основания-нуклеотиды: аденин (A), гуанин (G), тимин (T) и цитозин (C).
Согласно правилу комплементарности нуклеотиды A и G первой цепи связаны соответственно с нуклеотидами T и C второй цепи [1].
Физический процесс плавления молекулы ДНК возникает при повышении температуры и связан с переходом "спираль — клубок". Термодинамическая теория перехода ДНК "спираль — клубок" основана на учете разности свободных энергий спирального и клубкообразного состояний [2]. Процесс плавления сопровождается значительным изменением физических свойств исследуемого раствора ДНК, позволяющим проводить анализ: поглощение в ультрафиолетовой части спектра (260 нм) возрастает на 30–40 % [3], а вязкость раствора снижается в 12 раз [4].
Современный метод анализа кривых плавления молекулы ДНК реализуется на анализаторах нуклеиновых кислот после ПЦР в реальном времени с применением флуоресцентных красителей, интенсивность которых может изменяться более чем в 100 раз [5]. При этом получаются кривые плавления продуктов амплификации, представляющие собой зависимость измеренной величины флуоресцентного отклика от температуры образца. При вторичной обработке каждой кривой плавления определяется температура плавления ДНК Tm как точка перегиба этой кривой. Значение температу- ры Tm соответствует энергетическому состоянию, при котором половина молекул ДНК находится в спиральном, а другая половина — в клубкообразном состоянии [3].
Температура плавления T m зависит от состава фрагмента ДНК, это объясняется энергетическим состоянием ДНК: в паре нуклеотидов GC имеются три водородные связи, в паре АТ — только две. Поэтому чем больше пар GC в ДНК, тем более она "тугоплавка" [6]. В 1962 г. на основе анализа нескольких десятков образцов [7] была выявлена линейная зависимость между температурой T m и содержанием GC-пар нуклеотидов [G + C]:
T m = 69.3 +0.41[G + C], (1)
где [G + C] — содержание нуклеотидов G и С в продукте ПЦР (%). В этой и следующих моделях константы имеют единицы измерения, обеспечивающие единство размерности всех слагаемых в этих моделях.
На значение температуры T m оказывают влияние и другие факторы: например, температура плавления фрагмента ДНК обратно пропорционально зависит от длины этого фрагмента [8, 9] и является линейной функцией логарифма концентрации ионов металлов в анализируемом растворе [10, 11]. Эти факторы учтены в следующей модели [12]:
T m = 81.5 + 16.6 lg ([K+]) +
+ 0.41 [G + C] – 675 / L p , (2) где [K+] — концентрация ионов калия (моль/л); L p — длина продукта (пар нуклеотидов, п. н).
Известна программа Tm_Tool [13] для расчета температуры плавления ДНК. В описании программы представлено следующее уравнение для последовательностей ДНК, длиннее чем 50 п. н., при pH раствора от 5 до 9:
T m = 81.5 + 41 (yG + zC) / (wA + xT + yG + zC) – – 500 / (wA + xT+ yG + zC) + +16.6 log 10 ([Na+]) – 0.62 F , (3)
где yG, zC, wA и xT — количество соответствующих нуклеотидов; [Na+] — концентрация в растворе ионов натрия (моль/л); F — процентная концентрация формамида (%). Видно, что увеличение концентрации формамида линейно снижает температуру T m [14].
Температура плавления зависит также от pH среды, значительно снижаясь при значениях pH ниже 5 и выше 9, что связано с эффектами протонирования и депротонирования азотистых оснований в кислой и щелочной средах [15]. Существенно на значение T m влияют тип и концентрация флуоресцентного красителя. Так, изменение температуры плавления может достигать (0.7 ± 0.1)°C при использовании красителя SYTO-64 и (10 ± ± 0.02)°C при использовании красителя SYBR Green I [16]. Зависимость интенсивности флуоресцентного отклика свободного красителя от температуры [17] также может влиять на значение температуры T m .
Зависимость температуры T m от некоторых из описанных выше факторов является линейной только при неизменных других факторах. Например, концентрация формамида может изменять температуру T m в диапазоне 2.4–2.9 ºC/моль в зависимости от содержания GC-пар нуклеотидов [G + C] [14]. Такая зависимость значительно усложняет модель для вычисления температуры плавления ДНК.
Существующие модели не учитывают всех факторов, влияющих на величину температуры плавления ДНК, погрешность вычисления темпе- ратуры Tm превышает 1 °C, что является неприемлемым для большого количества прикладных задач [18, 19]. Таким образом, существует необходимость создания новых моделей, позволяющих более точно определять значение температуры Tm при изменении некоторых факторов, например длины фрагментов ДНК и содержания GC-пар нуклеотидов, и неизменных иных факторах.
ПРИМЕР МОДЕЛИРОВАНИЯ ЗАВИСИМОСТИ ВЕЛИЧИНЫ T m ОТ ДЛИНЫ ФРАГМЕНТОВ ДНК И СОДЕРЖАНИЯ GC-ПАР НУКЛЕОТИДОВ
Формулы (2) и (3) можно представить в виде упрощенной модели, учитывающей изменение значений длины фрагмента ДНК L p и содержания нуклеотидов [G + C], справедливой для экспериментов с неизменными прочими параметрами:
T mc = P1+ 0.41 [G + C] – P2 / L p . (4)
Здесь исходное значение P1 при постоянном составе среды равно сумме 81.5 + 16.6 lg [K+] на основании формулы (2) или 81.5 + 16.6 lg([Na+]) – – 0.62 F , на основании формулы (3).
Значения модельных констант P1 и P2 вычисляются на основе данных, полученных в программе Tm_Tool, по методу наименьших квадратов (МНК) [20, 21]. Определяется сумма квадратов отклонений Q модели Tmc от экспериментальных данных Tm :
n - 1 \2
Q = ! ( T mc ( L p , [ G + C ] ,P1,P2 ) - T m ) , (5)
i = 0
где n — количество измерений.
Минимум суммы квадратов отклонений Q определяется из условий равенства нулю частных производных функции Q путем изменения коэффициентов L p , [G + C], P1 и P2 .
Табл. 1. Оценка отклонения вычисленного значения T m
|
№ фрагмента |
L p , п. н. |
T °C m , , по Tm_Tool |
T m , °C, по ф-ле (6) ( T m(6) ) |
Δ T m |
|
1 |
100 |
85.69 |
85.60 |
–0.09 |
|
2 |
200 |
87.59 |
87.80 |
0.21 |
|
3 |
400 |
88.91 |
88.89 |
–0.02 |
|
4 |
600 |
89.28 |
89.26 |
–0.02 |
|
5 |
800 |
89.48 |
89.44 |
–0.04 |
|
6 |
1000 |
89.59 |
89.55 |
–0.04 |
В столбце " T m , °C, по Tm_Tool" табл. 1 представлены значения температуры плавления T m , рассчитанные с помощью известной программы Tm_Tool при содержании нуклеотидов [G + C] = = 50 % в зависимости от длины фрагментов L p .
Формула (4), уточненная в программной среде MatLab с помощью функции regress по значениям T m , L p и [G + C], имеет вид:
T m = 69.5 +0.41 [G + C] – 438.4 / L p . (6)
Для оценки отклонения вычисленной температуры плавления T m по формуле (6) в столбце T m(6) табл. 1 находятся значения, рассчитанные по этой формуле в зависимости от длины фрагментов L p . Значение Δ T m соответствует разности значений " T m по Tm_Tool" и T m(6) .
Максимальное отклонение вычисленной температуры плавления T m по формуле (6) составляет 0.21 °C, значение СКО составляет 0.05 °C. Для оценки адекватности модели, полученной при использовании программы Tm_Tool, необходимо сравнить ее результаты с результатами эксперимента.
ЭКСПЕРИМЕНТАЛЬНОЕ ОПРЕДЕЛЕНИЕ ЗНАЧЕНИЙ ТЕМПЕРАТУРЫ ПЛАВЛЕНИЯ
ФРАГМЕНТОВ ДНК T M
Исходные (сырые) данные получены методом анализа кривых плавления на анализаторе нуклеиновых кислот АНК-32, который серийно выпускается в Институте аналитического приборостроения РАН (ИАП РАН).
Выполнен трехкратный анализ 3 образцов ампликонов цитокератина (CK-19) в четырехкратном повторении по методике плавления в диапазоне температур образца 70–95 °C с шагом 0.5 °C и длительностью выдержки каждого значения температуры 30 с.
С помощью программы ANK_Melting [22] определены 12 величин T m каждого образца. Вычислены средние значения T m каждого образца: 84.30, 90.42 и 90.55 °C. Погрешность определения средней температуры плавления T m для этих образцов оценена с помощью значения СКО, которое составляет 0.12 °C.
В верхней части рисунка показаны кривые плавления, полученные при анализе трех образцов с разной температурой плавления. В нижней части рисунка показаны производные от этих кривых по температуре.
Средние значения T m для образцов 1, 2 и 3 приведены в столбце T mА табл. 2. Образцы 1, 2 и 3 имеют соответственно содержание нуклеотидов [G + C], равное 53, 59 и 58 % при длине фрагментов L p , равной 103, 209 и 309 п. н. соответственно.
Параметры среды в эксперименте соответствуют параметрам, принятым при расчете значений T m с помощью программы Tm_Tool. Это позволяет оценить соответствие формулы (6) экспериментальным данным. Отклонение вычисленного значения T m образцов 1, 2 и 3 оказалось значительным и составляет 2.60, 1.45 и 1.09 °C соответственно. Для уточнения предложенной модели предлагается определить модельные константы в соответствии с экспериментальными данными.
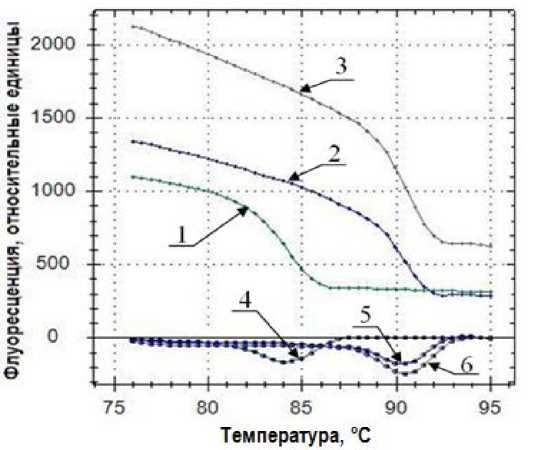
Графики плавления (линии 1–3), полученные при анализе трех образцов, и их производные по температуре (линии 4–6)
Табл. 2. Определение модельных констант P1 и P2
|
№ образца |
[G + C], % |
L p , п. н. |
T m(7) , °C |
T mА , °C |
Δ T m |
|
1 |
53 |
103 |
84.37 |
84.30 |
0.07 |
|
2 |
59 |
209 |
90.14 |
90.42 |
–0.29 |
|
3 |
58 |
309 |
90.77 |
90.55 |
0.22 |
ОПРЕДЕЛЕНИЯ МОДЕЛЬНЫХ КОНСТАНТ ПРИ ВЫЧИСЛЕНИИ ТЕМПЕРАТУРЫ
ПЛАВЛЕНИЯ ФРАГМЕНТОВ ДНК ПО РЕЗУЛЬТАТАМ АНАЛИЗОВ
Значения модельных констант P1 и P2 формулы (6) вычисляются на основе данных T mА по методу наименьших квадратов.
Формула, модельные константы которой определены по экспериментальным данным в программе Matlab, имеет вид:
T m = 69.2 + 0.41 [G + C] – 671.8/ L p . (7)
В столбце T m(7) приведены значения температуры плавления T m , рассчитанные по формуле (7) в зависимости от величин [G + C] и длины фрагментов L p . Значение Δ T m в табл. 2 соответствует разности T m(7) – T mA .
Максимальное отклонение вычисленной температуры плавления T m по формуле (7) соответствует значению 0.29 °C, значение СКО равно 0.13°C.
Модель обеспечивает возможность сравнивать исследуемые образцы по экспериментальным результатам определения величин T m . Например, изменение длины фрагмента L p от 100 до 110 п. н. вызывает изменение T m на (671.8 / 100) – – (671.8 / 110) = 0.6 °C, а изменение состава [G + C] на 1 % вызывает изменение T m на 0.41 °C.
ЗАКЛЮЧЕНИЕ
Предложено применение упрощенных моделей для вычисления температуры плавления ДНК T m , учитывающих зависимость от некоторых факторов и справедливых для экспериментов с неизменными прочими факторами. В качестве примера приведена упрощенная модель, учитывающая зависимость от длины фрагмента ДНК и содержания GC-пар нуклеотидов.
Описана методика определения модельных констант для вычисления температуры плавления фрагментов ДНК на основе экспериментальных данных и данных из известных литературных источников.
На анализаторе нуклеиновых кислот АНК-32 проведен трехкратный анализ методом плавления трех известных образцов ДНК с четырехкратным повторением. В результате выполнена оценка погрешности определения средней температуры плавления Tm: значения СКО для этих образцов во всех пробирках — не более 0.12 °C.
Отклонение вычисленной температуры плавления T m при использовании уточненной по экспериментальным данным модели не превышает 0.29 °C, значение СКО составляет 0.13 °C.
Работа выполнена в ИАП РАН в рамках государственного задания № 075-01073-20-00 Министерства науки и высшего образования РФ.
Список литературы Определение модельных констант при вычислении температуры плавления ДНК
- Chargaff E., Lipshitz R., Green C., Hodes M.E. The composition of the desoxyribonucleic acid of salmon sperm // J. Biol. Chem. 1951. Vol. 192. P. 223-230. PMID ISBN: 14917668
- Веденов А.А., Дыхне А.М., Франк-Каменецкий М.Д. Переход спираль-клубок в ДНК // Успехи физических наук. 1971. Т. 105, № 11. С. 479-519. DOI: 10.3367/UFNr.0105.197111d.0479
- Wartell R.M., Benight A.S. Thermal denaturation of DNA molecules: A comparison of theory with experiment // Physics Reports. 1985. Vol. 126. P. 67-107. DOI: 10.1016/0370-1573(85)90060-2
- Rice S.A., Doty P. The Thermal Denaturation of Deoxyribose Nucleic Acid // Journal of American Chemical Society. 1957. Vol. 79. P. 3937-3947. DOI: 10.1021/ja01572a001
- Wittwer C.T., Herrmann M.G., Moss A.A., Rasmussen R.P. Continuous fluorescence monitoring of rapid cycle DNA amplification // Biotechniques. 1997. Vol. 22. P. 134-138. DOI: 10.2144/97221bi01
- Marmur J., Doty P. Heterogeneity in Deoxyribonucleic Acids: I. Dependence on Composition of the Configurational Stability of Deoxyribonucleic Acids // Nature. 1959. Vol. 183, no. 4673. P. 1427-1429.
- DOI: 10.1038/1831427a0
- Marmur J., Doty P. Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature // Journal of Molecular Biology. 1962. Vol. 5, no. 1. P. 109-118.
- DOI: 10.1016/s0022-2836(62)80066-7
- Porschke D. Cooperative non-enzymic base recognition II. Thermodynamics of the helix-coil transition of oligoadenylic + oligouridylic acids // Biopolymers. 1971. Vol. 10. P. 1989-2013.
- DOI: 10.1002/bip.360101016
- Blake R.D. Cooperative lengths of DNA during melting // Biopolymers. 1987. Vol. 26. P. 1063-1074.
- DOI: 10.1002/bip.360260706
- Breslauer K.J., Frank R., Blocker H., Marky L.A. Predicting DNA duplex stability from the base sequence // Proc Natl Acad Sci USA. 1986. Vol. 83. P. 3746-3750.
- DOI: 10.1073/pnas.83.11.3746
- Owczarzy R., You Y., Moreira B.G., Manthey J.A., Huang L., et al. Effects of sodium ions on DNA duplex oligomers: improved predictions of melting temperatures // Biochemistry. 2004. Vol. 43. P. 3537-3554.
- DOI: 10.1021/bi034621r
- Календарь Р.Н., Сиволап Ю.М. Полимеразная цепная реакция с произвольными праймерами // Биополимеры и клетка. 1995. Т. 11, № 3-4. С. 55-65. URL: http://www.biopolymers.org.ua/pdf/ru/11/3/055/biopolym.cell-1995-11-3-055-ru.pdf
- DNA.UTAH.EDU Software Methods and Equations. URL: https://www.DNA.UTAH.edu/tm/tool.html
- Blake R.D., Delcourt S.G. Thermodynamic effects of formamide on DNA stability // Nucleic Acids Res. 1996. Vol. 24, no. 11. P. 2095-2103.
- DOI: 10.1093/nar/24.11.2095
- Privalov P.L., Ptitsyn O.B., Birshtein T.M. Determination of stability of the DNA double helix in an aqueous medium // Biopolymers. 1969. Vol. 8, no.5. P. 559-571.
- DOI: 10.1002/bip.1969.360080502
- Gudnason H., Dufva M., Bang D.D., Wolff A. Comparison of multiple DNA dyes for real-time PCR: effects of dye concentration and sequence composition on DNA amplification and melting temperature // Nucleic Acids Res. 2007. Vol. 35, no. 19. e127.
- DOI: 10.1093/nar/gkm671
- Сочивко Д.Г., Варламов Д.А., Федоров А.А., Курочкин В.Е. Метод бесконтактного измерения температуры в реакторах полимеразной цепной реакции // Письма в ЖТФ. 2016. Т. 42, вып. 7. С. 53-58. URL: http://journals.ioffe.ru/articles/viewPDF/43031
- Ломзов А.А., Пышный Д.В. Расчет температуры плавления нативных и модифицированных комплексов ДНК при различных концентрациях катионов металлов с помощью расширенной модели конденсации противоионов // Вестник НГУ. Серия: Физика. 2008. Т. 3, № 2. С. 61-75. URL: https://lib.nsu.ru/xmlui/handle/nsu/3282
- Vologodskii A., Frank-Kamenetskii M.D. DNA melting and energetics of the double helix // Physics of Life Reviews. 2018. Vol. 25. P. 1-21.
- DOI: 10.1016/j.plrev.2017.11.012
- Мудров А.Е. Численные методы для ПЭВМ на языках Бейсик, Фортран и Паскаль. Томск: МП "РАСКО", 1991. 272 с.
- Лизунова Н.А., Шкроба С.П. Матрицы и системы линейных уравнений. М.: ФИЗМАТЛИТ, 2007. 171 с.
- Белов Д.А., Белов Ю.В., Широкорад А.Л. Разработка экспериментальной версии программного обеспечения на основе новой методики определения температуры плавления ДНК // Научное приборостроение. 2018. Т. 28, № 2. С. 11-19. URL: http://iairas.ru/mag/2018/abst2.php#abst2
