Осевой градиент содержания олеиновой кислоты в частях семени подсолнечника

Автор: Демурин Я.Н., Борисенко О.М., Чебанова Ю.В.
Рубрика: Селекция и семеноводство
Статья в выпуске: 1 (157-158), 2014 года.
Бесплатный доступ
Представлены результаты исследования изменчивости содержания олеиновой кислоты в различных частях семени у инбредных линий подсолнечника генетической коллекции ВНИИМК. У среднеолеиновой линии ЛГ27 наблюдается увеличение содержания олеиновой кислоты от геммулы к дистальному концу семядолей зародыша на 13,1 %, у повышенноолеиновой ВК678 -на 12,8 % и у низкоолеиновой ЛГ28 - на 5,5 %. Высокоолеиновая линия ЛГ26 не показала достоверных различий в содержании изучаемого признака между частями семени.
Олеиновая кислота, семена, градиент содержания
Короткий адрес: https://sciup.org/142151178
IDR: 142151178 | УДК: 633.854.78:575
The longitudinal gradient of oleic acid content in different parts of sunflower seeds
The results of the research on variability of oleic acid content in different parts of a seed of inbred lines of sunflower genetic collection at VNIIMK are presented. The mid oleic line LG27 had the highest increase of oleic acid content from gemmule to the distal end of cotyledon, about 13.1 %, the increased oleic line VK678 had 12.8 % and the low oleic line LG28 had 5.5 %. The high oleic line LG26 did not show any differences in investigated character between the seed parts.
Текст научной статьи Осевой градиент содержания олеиновой кислоты в частях семени подсолнечника
Введение. Использование в селекции подсолнечника на качество масла метода прижизненного анализа отдельных семян, основанного на биохимической оценке части семени с последующим отбором и выращиванием оставшегося зародыша, вызвало необходимость проведения исследований фенотипической однородности жирнокислотного состава у различных частей семени.
Первое изучение подобного осевого (продольного) градиента с нарезанием семени на три части показало наличие достоверного увеличения содержания олеиновой кислоты от «кончика» семени к «верхушке» семядолей у высокоолеиновой линии с 87,5 до 91,3 %, т.е. на 3,8 %, а у обычной линии – с 44,5 до 56,9 %, т.е. на 12,5 %, при выращивании растений в условиях высокой температуры воздуха [1]. Дальнейшее исследование испанских учёных с разрезанием отдельного семени на четыре равные части также позволило описать повышение содержания олеиновой кислоты к «верхушке» семядолей на 13,2 % для обычной линии CAS-6 [2].
Плод подсолнечника – семянка – состоит из семени и околоплодника (перикарпа). Семя включает в себя семенную оболочку, остаток эндосперма и собственно зародыш, который в свою очередь подразделяется на две семядоли и геммулу, состоящую из зародышевых корешка, гипокотиля и почечки
(плюмулы). При этом все части зародыша содержат клетки с высоким содержанием депонируемого жира с масличностью семени около 65 % [3; 4].
В лаборатории генетики ВНИИМК ранее было определено увеличение в семядолях среднеолеиновой линии ЛГ27 содержания олеиновой кислоты на 12,7 % по отношению к геммуле, тогда как у высокоолеиновых и нормальных линий статистически достоверных отличий не установлено [5].
В данной статье представлены результаты изучения гетерогенности семян по содержанию олеиновой кислоты у гомозиготных линий генетической коллекции ВНИИМК по признаку жирно-кислотного состава масла.
Материалы и методы. В исследовании были использованы четыре инбредные линии: низкоолеиновая ЛГ28, повышенноолеи-новая ВК678, среднеолеиновая ЛГ27 и высокоолеиновая ЛГ26.
Выращивание и самоопыление растений проводили в полевых условиях на ЦЭБ ВНИИМК в 2007, 2012 и 2013 гг. В случае изучения градиента содержания олеиновой кислоты отдельные семянки были очищены от лузги, а зародыш семени разделен на три части: геммулу, среднюю и дистальную части семядолей, которые анализировались отдельно (рис. 1).

Рисунок 1 – Поперечный разрез семени на примере линии ЛГ27
(1 – геммула, 2 – средняя часть семядолей, 3 – дистальная часть семядолей)
Для анализа жирно-кислотного состава использовали средние пробы трех вышеописанных частей семени (по 25 семян) при навеске 20 г. Анализ выполняли в трех повторностях на основе газо-жидкостной хроматографии метиловых эфиров на хроматографе Хром-5.
Результаты и обсуждение. Все четыре изученные линии характеризовались дис- кретными фенотипическими классами по содержанию олеиновой кислоты (табл. 1). Низ-коолеиновая ЛГ28 (30,6 %) и повышенноолеиновая ВК678 (47,3 %) показали крайние значения нормального фенотипа. Среднеолеиновая ЛГ27 (67,4 %), или mid-oleic, приближается по фенотипу к высоко-олеиновой ЛГ26 (88,1 %), или high oleic. При этом ЛГ27 не обладает доминантной мутацией высокоолеиновости Ol, присутствующей у ЛГ26.
Таблица 1
Фенотипы линий генетической коллекции подсолнечника по жирно-кислотному составу масла в семенах у линии ЛГ28 – на 5,5 %. Линия ЛГ26 не показала достоверных отличий (рис. 2).
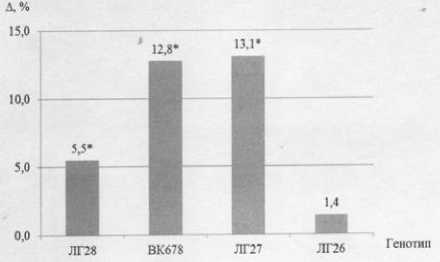
Рисунок 2 – Разница (∆) между семядолями и геммулой по содержанию олеиновой кислоты в семенах линий подсолнечника, * – р < 0,05
ВНИИМК, Краснодар, 2007, 2012 и 2013 гг.
|
Линия |
Фенотипический класс |
Жирно-кислотный состав, % |
|||
|
С16:0 |
С18:0 |
С18:1 |
С18:2 |
||
|
ЛГ28 |
Низко-олеиновый |
7,9 |
1,7 |
30,6 |
59,7 |
|
ВК678 |
Повышенно-олеиновый |
5,7 |
5,1 |
47,3 |
41,9 |
|
ЛГ27 |
Средне-олеиновый |
4,4 |
3,7 |
67,4 |
24,5 |
|
ЛГ26 |
Высоко-олеиновый |
5,2 |
4,5 |
88,1 |
2,1 |
У линий ЛГ28, ВК678 и ЛГ27 наблюдалось достоверное увеличение содержания олеиновой кислоты от геммулы к средней части семядолей (табл. 2). При этом между их средней и дистальной частями различия были не достоверными. С другой стороны, семена высокооолеиновой ЛГ26 были однородными по содержанию олеиновой кислоты.
Таблица 2
Содержание олеиновой кислоты в различных частях зародыша семени у инбредных линий подсолнечника, %
ВНИИМК, Краснодар, 2013 г.
|
Линия |
Гемму-ла |
Средняя часть семядолей |
Дистальная часть семядолей |
HCP 05 |
|
ЛГ28 |
28,7 |
34,2 |
30,6 |
3,7 |
|
ВК678 |
37,6 |
48,0 |
50,4 |
2,9 |
|
ЛГ27 |
52,9 |
65,9 |
65,9 |
4,6 |
|
ЛГ26 |
87,9 |
86,7 |
86,5 |
1,6 |
Различия в изучаемом градиенте прямо пропорциональны увеличению общего содержания олеиновой кислоты в семенах. Так, у линии ЛГ27 наблюдался рост содержания олеиновой кислоты к дистальному концу зародыша на 13,1 %, у линии ВК678 – на 12,8, а
Феномен положительного осевого градиента в содержании олеиновой кислоты в семенах подсолнечника от геммулы к семядолям, вероятно, объясняется отрицательным градиентом концентрации кислорода. Последний может быть связан с локально направленным проникновением кислорода в ткани семени через место его прикрепления к цветоложу корзинки при условии газовой непроницаемости семенных покровов. Известно, что кислород является необходимым фактором в процессе десатурации олеиновой кислоты в линолевую. В частности, молекулярный кислород, поступающий в цитоплазму клеток семян посредством диффузии, является акцептором электронов при десатурации [6]. С другой стороны, отсутствие осевого градиента у высокоолеиновой линии ЛГ26 связано, очевидно, с отсутствием самой реакции десатурации из-за генетического блока превращения олеиновой кислоты в линолевую под действием мутации Ol [7].
Заключение. Явление осевого градиента по содержанию олеиновой кислоты в семенах должно учитываться при изучении наследования признака жирно-кислотного состава и практической селекции на качество масла с использованием метода половинок семян (half-seed-technique).
