Основные направления популяционной динамики в Северной Евразии от мезолита до эпохи ранней бронзы (по данным краниологии и генетики)
")
Автор: Козинцев А.Г.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология и палеогенетика
Статья в выпуске: 4 т.49, 2021 года.
Бесплатный доступ
Цель исследования - рассмотреть краниометрические данные о популяционной изменчивости на территории Северной Евразии под углом зрения генетических и отчасти лингвистических фактов. Измерения 66 серий мужских черепов разных эпох (от мезолита до раннего бронзового века) с этой территории обработаны статистическими методами, специально предназначенными для изучения пространственных закономерностей, в частности градиентов. С помощью неметрического многомерного шкалирования матрицы расстояний D2 (с поправкой на численность) получена двумерная проекция взаимоположения групп, построено минимальное остовное дерево, показывающее кратчайший путь между точками в многомерном пространстве. Обнаруживаемые генетическими и краниометрическими данными восточно-западные градиенты на территории Северной Евразии свидетельствуют, видимо, не столько о смешении, сколько о незавершенности процесса дифференциации бореального надрасового ствола. Западный компонент, представленный в Сибири и Центральной Азии носителями афанасьевской культуры, вероятно, мало повлиял на генетический облик местных популяций. Восточный компонент, проникший в неолите из Забайкалья в Прибайкалье, смешался там с автохтонным палеосибирским. Главный генетический маркер коренного населения - аутосомный компонент ANE - присутствовал в Сибири с верхнего палеолита. Автохтонными следует считать и обе евразийские формации - северную и южную. Статистический анализ позволил включить в их состав новые группы, причем границы первой расширились на восток до Кузнецкой котловины, а второй - на запад до Среднего Прииртышья. Участие восточно-европейских групп в генезисе северной евразийской формации и протоуральской общности обнаружить не удается.
Южная сибирь, западная сибирь, восточная европа, мезолит, неолит, ранний бронзовый век, краниометрическая дифференциация
Короткий адрес: https://sciup.org/145146494
IDR: 145146494 | УДК: 572 | DOI: 10.17746/1563-0102.2021.49.4.140-151
Текст научной статьи Основные направления популяционной динамики в Северной Евразии от мезолита до эпохи ранней бронзы (по данным краниологии и генетики)
Новейшие достижения палеогенетики, особенно в области широкогеномного анализа, все чаще помогают решить споры, которые антропологи вели на протяжении многих десятилетий. Яркий пример – дискуссия о том, было ли краниологическое своеобразие древних прибайкальцев вызвано европеоидной примесью [Дебец, 1948, с. 61] или же сохранением самостоятельного палеосибирского типа [Дебец, 1951, с. 95]. Краниологи так и не пришли к согласию: одни считали, что европеоидная примесь была минимальной [Мамонова, 1983], другие отводили ей решающую роль и выступали за упразднение термина «палеосибирский тип» [Алексеев, Гохман, Тумэн, 1987]. Работы генетиков поставили точку в этом споре. Они убедительно продемонстрировали, что своеобразие прибайкальцев вызвано не европеоидной примесью, по крайней мере не той, которая, как полагал Г.Ф. Дебец, проникала с запада по степям начиная с раннего бронзового века [Damgaard et al., 2018]. Такая примесь обнаружилась лишь у одного из 53 индивидов эпох неолита и ранней бронзы из Прибайкалья и Забайкалья (1,9 %) – судя по всему, у мужчины из глазковского могильника Хап-цагай на верхней Лене [Yu et al., 2020]*. Прочие же индивиды из погребений этого времени в Прибайкалье уклоняются от наиболее «чистых» монголоидов – носителей компонента NEA (Northeast Asian), особенно выраженного у неолитических обитателей Чертовых Ворот в Приморье и китойцев**, – в сторону не европеоидов бронзового века, а верхнепалеолитических жителей Сибири, носителей компонента ANE (Ancient North Eurasian), который наиболее выражен у мальчика со стоянки Мальта древностью ок. 24 тыс. лет, а также у муж- чины и девочки со стоянки Афонтова Гора древностью 15–17 тыс. лет [Raghavan et al., 2014; Fu et al., 2016]. В Прибайкалье доля ANE увеличивается с 14 % у ки-тойцев до 23 % у представителей более поздних стадий неолита и раннего бронзового века [Yu et al., 2020].
Итак, сдавать в архив термин «палеосибирский тип» едва ли целесообразно, поскольку речь, судя по всему, идет о морфологическом корреляте компонента ANE. Конечно, это не предполагает прямой зависимости строения черепа от аналитически выделенной части генома, обеспечивающей наиболее эффективную классификацию групп. Однако косвенная связь вполне вероятна, т.к. и краниологические типы были выделены для решения той же задачи.
Происхождение компонента ANE – огромная загадка. Хотя археологические параллели мальтинско-буретской культуре на территории Восточной и Центральной Европы общеизвестны, предположение о том, что она появилась в результате миграции оттуда в Cибирь, данными генетики не подтверждается [Fu et al., 2016]. Не находит опоры и противоположная гипотеза: хотя компонент ANE проник в Центральную и Западную Европу с во стока, произошло это не в верхнем палеолите, а гораздо позже, т.к. до экспансии носителей ямной культуры его там не было [Ibid.; Haak et al., 2015].
ANE составляет основную долю в компоненте EHG (Eastern Hunter-Gatherer), который особенно выражен в геномах мезолитических и ранненеолитических обитателей Северо-Восточной Европы (Южный Олений о стров, Попово, Сиделькино, Лебяжин-ка IV и др.) [Haak et al., 2015; Damgaard et al., 2018]. От них EHG унаследовали ямники и их потомки, в т.ч. афанасьевцы. С территории Северной России этот компонент еще в мезолите проник в Скандинавию, о чем свидетельствуют геномы людей из Муталы в Южной Швеции. Их предки мигрировали туда с востока вдоль норвежского побережья, поскольку южнее, в Восточной Прибалтике, у носителей более ранней кундской культуры доля EHG ниже [Haak et al., 2015;
Mittnik et al., 2018]. Как показывают разновременные материалы из могильника Звейниеки (Латвия), она увеличивается в среднем неолите вместе с археологическими свидетельствами влияния культуры ямочногребенчатой керамики [Jones et al., 2017].
Корни ANE в Сибири гораздо глубже, чем корни EHG в Европе, куда, судя по митохондриальным гаплогруппам, ANE проник с востока [Mittnik et al., 2018]. Предковым для по следнего оказался компонент ANS (Ancient North Siberian), представленный у мужчины с Янской стоянки древностью 31,6 тыс. лет [Sikora et al., 2019]. Этот компонент возник, видимо, в пределах ранней ветви западного ствола вскоре после его отделения от восточного ок. 43 тыс. л.н. Картина осложняется восточной примесью (ок. 22 %), полученной представителями западного ствола на ранних стадиях его существования [Ibid.].
Из Сибири ANE распространялся и в сторону Европы, и в сторону Америки. Предки американских индейцев принесли его в Новый Свет, и сейчас его доля там составляет 30–40 % [Flegontov et al., 2016; Sikora et al., 2019]. На территории же Старого Света к американским аборигенам в этом отношении ближе всего чукчи, коряки, кеты и селькупы, а из древних групп – окуневцы и ботайцы*, что позволяет считать последних прямыми наследниками носителей маль-тинско-буретской культуры. В обеих группах, как и у прибайкальцев, ANE смешан с «монголоидным» компонентом. Такая же смесь представлена у двух мужчин, генетически очень близких к американским аборигенам, – с верхнепалеолитической стоянки Усть-Кяхта (ок. 14 тыс. л.н.) в Западном Забайкалье [Yu et al., 2020] и с мезолитического местонахождения Дуванный Яр (ок. 10 тыс. л.н.) на р. Колыме [Sikora et al., 2019].
Совершенно неясны пути проникновения ANE (в значительной доле!) к верхнепалеолитическим людям, жившим на Западном Кавказе ок. 13 тыс. л.н. (Са-цурблия), и ранненеолитическим обитателям Северного Ирана (Гандж-Даре, VIII тыс. до н.э.) [Lazaridis et al., 2018]. Судя по громадному ареалу ANE, он является наследием ранних сапиенсов, представителей бореального надрасового ствола [Biasutti, 1941, p. 275; Козинцев, 2013, 2014], заселявших Северную Евразию и Америку, находившихся на разных стадиях дифференциации этого ствола и в разной степени затронутых контактами между его ветвями.
Геномными исследованиями выявлен ряд ранних градиентов, проходящих вдоль Северной Евразии в широтном направлении и связывающих Северо-Восточную Европу (ареал EHG) с Забайкальем и Приаму- рьем (ареалом NEA/AEA) [Damgaard et al., 2018; Jeong et al., 2019; Narasimhan et al., 2019; Kilin^ et al., 2021]. Их можно было бы счесть генетическими коррелятами европеоидно-монголоидного вектора, традиционно выявляемого антропологами, если бы не тот факт, что древнейший носитель компонента ANE, играющего ключевую роль в этих градиентах, обитал не в западной и не в восточной их точке, а в промежуточной. И если значительную (верхнепалеолитическую) древность компонента NEA/AEA в Приамурье можно считать доказанной, как и его роль в ранней популяционной дифференциации на территории Северной Евразии [Mao et al., 2021; Yu et al., 2020], то европеоидный компонент, отличающийся от EHG большой долей южных примесей (иранской, кавказской и анатолийской), участвовал в данном процессе лишь с эпохи ранней бронзы, конкретно со времени ямно-афа-насьевских миграций. До этого роль равноправного партнера NEA/AEA принадлежала компоненту ANE.
Отсюда следует, что насущной задачей палеоантропологических исследований является изучение протоморфных вариантов вроде палеосибирского и родственных ему. Была высказана аргументированная гипотеза об автохтонности одного из них, названного «южной евразийской антропологической формацией», на территории Алтае-Саянского нагорья и прилегающих горных систем [Чикишева, 2012, с. 57, 153, 169]. Эталоном данного варианта служат окунев-цы. Другой вариант, представленный в ряде групп Ба-рабинской лесостепи эпох неолита и ранней бронзы, Т.А. Чикишева, используя термин В.В. Бунака [1956], называет «северной евразийской антропологической формацией» [Чикишева, 2012, с. 6, 56, 59, 123–124, 179–180]. С этой формацией, отчасти соответствующей уральской расе в широком смысле, автор сближает ряд мезолитических и неолитических групп лесной полосы Северо-Восточной Европы – те самые, которые маркируются компонентом EHG. Как считает Т.А. Чикишева, Барабинская лесостепь заселялась из лесной полосы севера Русской равнины в раннем голоцене [Чикишева, Поздняков, 2021]. Обе формации (северная и южная), по ее мнению, представляют собой равноправные ветви бореального ствола наряду с европеоидной и монголоидной.
А.Н. Багашёв [1998], оспаривая предложенную В.В. Бунаком [1956] концепцию уральской расы в широком смысле, отрицает генеалогическую связь между современными популяциями Западной Сибири, включаемыми им в западно-сибирскую расу, и восточно-европейскими, которые он считает европеоидными*. Результаты же краниоскопических
*У окуневцев к тому же есть европеоидная примесь ям-но-афанасьевского происхождения [Damgaard et al., 2018; Jeong et al., 2019; Yu et al., 2020].
исследований не только демонстрируют эту связь с полной убедительностью, но и противопоставляют современные уралоязычные группы всем прочим [Козинцев, 1988, с. 137–140; Kozintsev, 1992; Моисеев, Козинцев, 1998]. Интеграция данных по четырем независимым системам признаков – краниометрии, краниоскопии, одонтологии и дерматоглифике – делает данное противопоставление еще отчетливее [Моисеев, 2001]. Если, таким образом, отрицание общности происхождения уралоязычных народов по обе стороны Урала стало анахронизмом и если, как резонно утверждает В.В. Напольских [1997, с. 177–178], перед нами свидетельства существования «уральского пранарода во плоти и крови», возникает вопрос: как глубоко в прошлое прослеживается уральская общность? Как она соотносится с северной евразийской формацией?
Задача настоящей работы – переосмыслить краниологическую классификацию в свете новейших генетических и отчасти лингвистических данных с особым вниманием к соотношению метисно сти и протоморфности. Именно на это нацелены применяемые в работе методы.
Материал и методика
Использованы данные о мужских сериях черепов из погребений, относящихся к следующим культурам, эпохам и территориям*:
-
1. Бойсманская культура Приморья [Чикишева, 2012, с. 38–39];
-
2. Неолит Якутии [Там же];
-
3. Неолит и бронзовый век Забайкалья [Гохман, 1980];
-
4. Китойская стадия, Восточное Забайкалье [Мамонова, 1983];
-
5. То же, Западное Забайкалье [Там же];
-
6. То же, Фофановский могильник [Герасимова и др., 2010];
-
7. То же, р. Лена [Мамонова, 1983];
-
8. То же, р. Ангара [Там же];
-
9. То же, Шаманка (неопубликованные данные Д.В. Пежемского);
-
10. Исаковская стадия, р. Ангара [Мамонова, 1983];
-
11. Серовская стадия, р. Лена [Там же];
-
12. То же, р. Ангара [Там же];
-
13. Глазковская стадия, Западное Забайкалье [Там же];
-
14. То же, Фофановский могильник [Герасимова и др., 2010];
-
15. То же, о-в Ольхон [Мамонова, 1983];
-
16. То же, р. Лена [Там же];
-
17. То же, р. Ангара [Там же];
-
18. Неолит красноярско-канской лесостепи [Солодовников, Багашёв, Савенкова, 2020];
-
19. Неолит нижней Ангары [Там же];
-
20. Большемысская культура Бийского Приобья (Ит-куль, Костёнкова Избушка) [Чикишева, 2012, с. 36–37];
-
21. Неолит Барнаульского Приобья (Фирсово XI) [Солодовников, Тур, 2017];
-
22. Кузнецко-алтайская культура Верхнего Приобья (Солонцы-5, Усть-Иша) [Чикишева, 2012, с. 36–37];
-
23. Неолит и энеолит Среднего Прииртышья [Солодовников и др., 2019];
-
24. Неолит лесостепного Зауралья и Северного Казахстана (ботайская культура, Гладунино) [Хохлов, Китов, 2015];
-
25. Неолит Кузнецкой котловины [Солодовников, Тур, 2017];
-
26. Неолит Новосибирско-Каменского Приобья [Там же];
-
27. Неолит Барабинской лесостепи [Чикишева, 2012, с. 36–37; Чикишева, Поздняков, Зубова, 2015];
-
28. Усть-тартасская культура, Сопка-2/3 [Чикише-ва, 2012, с. 69–72];
-
29. То же, Сопка-2/3A [Там же];
-
30. Одиновская культура, Сопка-2/4A [Там же, с. 98–101];
-
31. То же, Тартас-1 [Чикишева, Поздняков, 2019];
-
32. То же, Преображенка-6 [Там же];
-
33. Кротовская культура, Сопка-2/4Б, В [Чикише-ва, 2012, с. 98–101];
-
34. Окуневская культура, Уйбат [Громов, 1997];
-
35. То же, Верх-Аскиз I [Там же];
-
36. То же, Черновая IV, VI, VIII [Там же];
-
37. То же, Тас-Хазаа [Там же];
-
38. Каракольская культура Алтая [Тур, Солодовников, 2005];
-
39. Чаахольская культура Тувы (Аймырлыг) [Гох-ман, 1980];
-
40. Елунинская культура Верхнего Приобья [Солодовников, Тур, 2003];
-
41. Самусьская культура Верхнего Приобья [Солодовников, 2005];
-
42. Чемурчекская культура Западной Монголии [Солодовников, Тумен, Эрдэнэ, 2019];
-
43. Афанасьевская культура, Западная Монголия [Гохман, 1980];
-
44. То же, Юго-Восточный Алтай [Солодовников, 2009];
-
45. То же, Северо-Западный Алтай [Там же];
-
46. То же, средняя Катунь [Там же];
-
47. То же, Усть-Куюм [Там же];
-
48. То же, Курота [Там же];
-
49. То же, р. Урсул [Там же];
-
50. То же, Сальдяр [Там же];
-
51. То же, Минусинская котловина [Там же];
-
52. То же, Карасук III [Алексеев, 1981];
-
53. То же, Афанасьева Гора [Там же];
-
54. Неолит и ранний энеолит Волго-Уралья [Хохлов, 2017, с. 219–223];
-
55. Мезолит севера Русской равнины, Южный Олений остров [Алексеев, Гохман, 1984];
-
56. То же, Попово [Гохман, 1984];
-
57. Мезолит Восточной Прибалтики, Звейниеки [Денисова, 1975, с. 187–188];
-
58. Ранний неолит Восточной Прибалтики, Звей-ниеки [Там же, с. 193–194];
-
59. Средний и поздний неолит Восточной Прибалтики, Звейниеки [Там же, с. 202–203];
-
60. Культура ямочно-гребенчатой керамики севера России и Волго-Окского междуречья [Чикишева, 2012, с. 38–39];
-
61. Волосовская культура Волго-Окского междуречья [Там же];
-
62. Хвалынская культура, Хвалынские могильники [Хохлов, 2017, с. 226–230];
-
63. То же, Хлопков Бугор [Там же, с. 230–231];
-
64. Мезолит Украины [Кондукторова, 1973, с. 13–14];
-
65. Неолит Украины, днепро-донецкая культура [Потехина, 1999, с. 190–192];
-
66. Ранний энеолит Украины, Средний Стог II [Там же, с. 204–208].
Программа включает 14 основных признаков (приводятся их стандартные обозначения, см.: [Алексеев, Дебец, 1964]): продольный (1), поперечный (8), высотный (17) диаметры, наименьшая ширина лба (9), скуловой диаметр (45), верхняя высота лица (48), высота носа (55), ширина носа (54), ширина орбиты (51), высота орбиты (52), назомалярный угол (77), зигомаксиллярный угол, симотический указатель (SS : SC), угол выступания носа (75 (1)). Данные обработаны с помощью канонического анализа и подсчета расстояний D2 c поправкой на численность. Матрица расстояний подвергнута неметрическому многомерному шкалированию. Построено минимальное остовное дерево (MST – minimum spanning tree), показывающее кратчайший путь между точками в многомерном пространстве и потому оптимальное для выявления градиентов. Использовались программа CANON Б.А. Козинцева и пакет PAST Э. Хаммера (версия 4.05) [Hammer, 2012]*.
-
*Бо лее ранние версии использовать не следует, т.к. они подсчитывают кратчайший путь не в исходном многомерном пространстве, а на плоскости.
Метод MST выделяет группировки менее формально, чем кластерный анализ. Но такие группировки легче согласовать с географическими данными, чем кластеры*. Кластерный анализ не годится для изучения клинальной изменчивости, а потому используется здесь лишь в качестве вспомогательного метода в двух разновидностях – невзвешенного парно-группового (UPGMA) и метода ближайшего соседа (NJ), реализованных в соответствующих модулях пакета PAST. Дендрограммы не приводятся ввиду их значительного размера, но высылаются электронной почтой по запросу.
Результаты
Взаимоположение и связи групп отражены на графике (рис. 1). Рассмотрим их в направлении главного градиента краниометрической изменчивости – от восточного (монголоидного) полюса к западному (европеоидному). Этот градиент в значительной мере совпадает с географическим.
Группы восточного полюса и палеосибирские . Серии с крайней выраженностью монголоидных особенностей (в порядке их убывания, т.е. в порядке возрастания значений 1-й координаты неметрического шкалирования) – китойская из Восточного Забайкалья (№ 4), бойсманская из Приморья (№ 1), китой-ская из Западного Забайкалья (№ 5) и неолитическая из Якутии (№ 2).
Действительно ли китойцы в среднем монголо-иднее представителей более поздних этапов неолита и бронзового века Байкальского региона, как следует из работ генетиков? Возможно, использование индивидуальных данных позволило бы подтвердить это, но на уровне средних величин результат получается неопределенным. Среднее значение 1-й координаты (рис. 1) в шести китойских сериях (№ 4–9) –0,147 ± ± 0,032, в восьми прочих из Забайкалья и Прибайкалья (№ 10–17) оно составляет –0,098 ± 0,007, разность, согласно критерию Манна–Уитни, вполне может быть случайной ( U = 16, p = 0,33). При учете только забайкальских и прибайкальских серий (рис. 2) отличие китойских от остальных остается недостоверным, не смотря на зашкаливающую монголоидность восточно-забайкальских китойцев**.
Географический фактор проявляется отчетливее хронологического. На рис. 1 видно, что забайкальская группировка значительно монголоиднее прибайкальской, в которую помимо всех девяти серий из При-
Рис. 1. Расположение мужских краниологических серий в пространстве двух координат неметрического шкалирования матрицы расстояний Махаланобиса.
Прямые линии – ребра минимального остовного дерева, показывающие кратчайший путь между точками в многомерном пространстве. I–VIII – группировки: I – палеосибирская, Забайкалье, II – палеосибирская, Прибайкалье, III – северная евразийская, IV – южная евразийская, V – окуневская, VI – ямочно-гребенчатая, VII – чаахольско-елунинская, VIII – афанасьевская. Штрихпунктиром обведены группы западного тяготения.
а – группы Восточной Сибири и Дальнего Востока; б – группы Южной и Западной Сибири, Западной Монголии, кроме афанасьевских; в – афанасьевские; г – европейские. Нумерация групп соответствует списку в тексте.
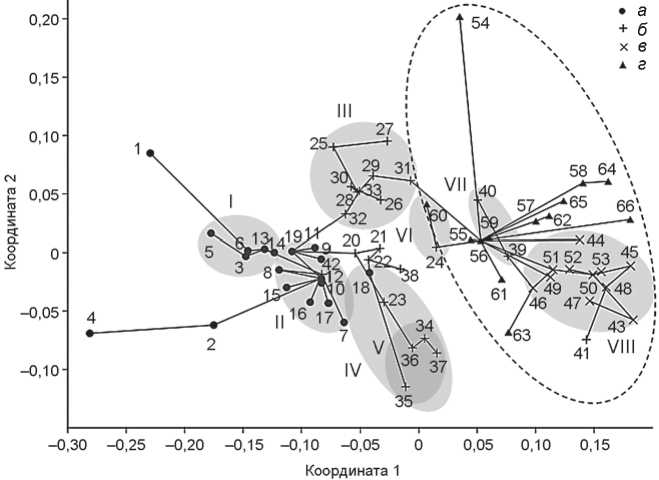
Рис. 2. Расположение мужских краниологических серий из Прибайкалья и Забайкалья в пространстве двух координат неметрического шкалирования матрицы расстояний Махаланобиса.
а – китойские; б – исаковские и серовские; в – глазковские; г – стадия не указана. Нумерация групп соответствует списку в тексте. Штрихпунктирная линия отделяет прибайкальские серии от забайкальских. Прямые линии – см. рис. 1.
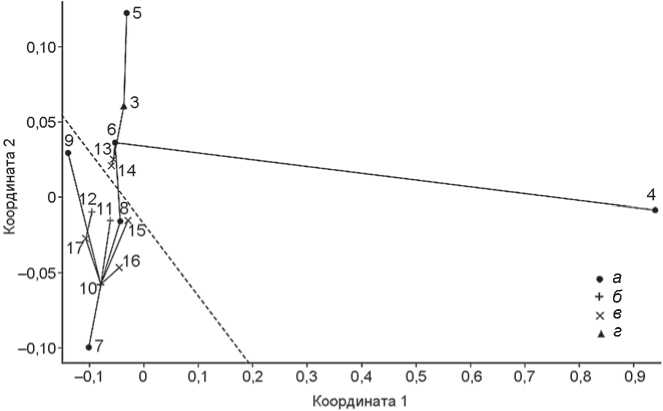
байкалья вошли два неолитических черепа с нижней Ангары (№ 19) и два чемурчекских черепа из Западной Монголии (№ 42)*. Среднее значение 1-й координаты для пяти забайкальских групп (суммарная забайкальская исключена) равно –0,172 ± 0,029, для девяти прибайкальских составляет –0,088 ± 0,006; вероятность случайности разницы, по критерию Манна-Уитни, очень мала (U = 0,5, p = 0,004). Различие заметно и на рис. 2, но лишь в виде тенденции, которая не достигает уровня значимости ни по одной из двух координат.
Евразийские формации – северная и южная. После работ Т.А. Чикишевой казалось, что ареал северной евразийской формации не простирается восточнее Оби. Помимо изученных ею семи групп эпох неолита и ранней бронзы из Барабинской лесостепи (см. рис. 1, № 27–33) в данную группировку попадает неолитическая серия из Новосибирско-Каменского Приобья (№ 26). Впрочем, кластерный анализ подтверждает это лишь в варианте NJ, но не UPGMA. Зато еще одна неолитическая серия, из гораздо более восточного района – Кузнецкой котловины (№ 25), объединяется с барабинскими по всем трем критериям (MST и оба варианта кластерного анализа).
Похоже, что ареал северной формации придется существенно расширить на восток, быть может, даже до Енисея. Кстати, из двух евразийских формаций именно северная, а не южная, имеет непосредственную связь с восточно-сибирской группировкой: ребро MST coединяет серии из Преображенки-6 (№ 32) и с нижней Ангары (№ 19).
Напротив, южная формация, как можно было думать до недавнего времени, распространялась от Енисея на запад, ведь лучшими претендентами на роль предков окуневцев (№ 34–37) казались неолитические обитатели красноярско-канской лесостепи (№ 18). Однако недавно выяснилось, что с не меньшим правом претендовать на эту роль могут люди, жившие в неолите и энеолите гораздо западнее – на среднем Иртыше (№ 23). Авторы публикации, правда, делят сборную группу по географическому принципу, усматривая у индивидов из лесной и лесостепной зоны черты северной формации, а у степных – южной [Солодовников и др., 2019]. Но серия очень невелика и по средним характеристикам входит в южную группировку, что подтверждается и результатами кластерного анализа. Четыре серии из Верхнего Приобья – большемысская из Иткуля и Костёнковой Избушки (№ 20), неолитическая из Фирсова XI (№ 21), кузнецко-алтайской культуры из Усть-Иши и Солонцов-5 (№ 22), а также каракольская (№ 38) – занимают промежуточное положение между двумя формациями, не попадая, согласно результатам кластерного анализа, ни в одну из них. Неподалеку от этих групп, в промежутке между ними и европеоидной группировкой располагается серия из лесостепного Зауралья и Северного Казахстана (№ 24), куда входят малочисленные ботайские черепа. Будучи самой европеоидной из неолитических групп Северной Азии, она образует обособленную пару с серией культуры ямочногребенчатой керамики (№ 60) – самой монголоидной из европейских.
Как пишет Т.А. Чикишева, сибирские группы, относящиеся к северной евразийской формации, в отличие от тех, которые относятся к южной, тяготеют к мезолитическим и неолитическим группам лесной полосы Северо-Восточной Европы. Действительно, ребро MST напрямую соединяет одиновскую серию из Тартаса-1 (№ 31) с мезолитической из Попова (№ 56). Но среднее расстояние D2 от восьми мезолитических и неолитических серий из лесной зоны Северо-Восточной Европы (№ 54–61) до девяти северно-евразийских групп Сибири (№ 25–33) не намного меньше, чем до шести южно-евразийских (№ 18, 23, 34–37) – 13,90 и 15,60 соответственно. Согласно критерию Уилкоксона для сопряженных наблюдений, различие не достигает порога достоверности (z = 1,82, p = 0,069). Поэтому говорить о единстве северной евразийской формации от Восточной Прибалтики до Кузнецкой котловины на основании имеющихся данных едва ли возможно. Пока можно лишь констатировать, что обе евразийские формации Сибири, вместе взятые, и группы, близкие к ним, заполняют промежуток между палеосибирскими, в частно сти прибайкальскими, и европейскими с ослабленной выраженностью «западных» черт.
Группы западного тяготения. Помимо уже упомянутых групп из лесной зоны, ослабление ев-ропеоидности наблюдается в хвалынской серии из Хлопкова Бугра (№ 63), которая по европеоидному масштабу заметно отличается от хвалынской из эпонимных могильников (№ 62), и в двух сибирских группах явно западного происхождения – чаа-хольской (№ 39) и елунинской (№ 40), образующих отдельную пару. Довольно сильна временнáя динамика в популяции из Звейниеки: умеренно европеоидная комбинация признаков в мезолите (№ 57) сменяется резко европеоидной в раннем неолите (№ 58), по сле чего европеоидность вновь существенно ослабляется и серия эпохи среднего и позднего неолита (№ 59) оказывается заметно сдвинутой в сторону ямочно-гребенчатой группировки. Противоположное, крайне западное сочетание признаков демонстрируют три группы из Украины (№ 64–66), а также большинство афанасьевских, к которым близка самусьская серия (№ 41).
Обсуждение
Подтвердить отличие китойцев в целом от представителей позднейших стадий байкальского неолита и бронзового века на краниологическом материале не удалось. Однако, несомненно, восточный компонент распространялся из Забайкалья, о чем свидетельствуют и крайняя выраженность монголоидных черт на двух китойских черепах из Восточного Забайкалья, опережающих в этом отношении даже бойсманскую серию, и прямая связь по следней с забайкальской группировкой, и достоверное отличие забайкальских серий от прибайкальских.
Поскольку европеоидная примесь в то время, как мы теперь знаем, Прибайкалья практически не достигала, расположение групп вдоль восточно-западной оси указывает на соотношение восточного компонента с палеосибирским (в генетических терминах NEA/AEA и ANE соответственно). То же относится и к обеим евразийским формациям. В эпоху, когда процесс распада бореального надрасового ствола еще не завершился, генетические и краниометрические градиенты, направленные от Восточной Сибири к Восточной Европе и, судя по всему, далее на запад, свидетельствуют, видимо, не столько о смешении, сколько об эффекте изоляции расстоянием. При этом на обширных территориях Евразии сохранялись группы, демонстрирующие все стадии перехода между двумя противоположными полюсами формообразования – восточным и западным.
Ближайшими к западному полюсу в нашей выборке являются три группы с территории Украины, в частности среднестоговская (№ 66), а также афанасьевцы. На то, что последние могли быть потомками средне стоговцев, указывали и археологи [Николаева, 2019а, б], и антропологи [Солодовников, 2009]. График (см. рис. 1) демонстрирует это достаточно наглядно. Временнáя динамика в группе из Звейниеки, в частности «восточный сдвиг» серии из погребений среднего и позднего неолита, заставляет вспомнить археологические свидетельства усиления связей с культурой ямочно-гребенчатой керамики и генетические данные о возрастании доли аутосомного компонента EHG на этом этапе [Jones et al., 2017].
Генетические факты говорят о том, что роль ранних мигрантов из западных степей как источника европеоидной примеси в Сибири не следует преувеличивать. По словам генетиков, афанасьевцы по пути на восток «перепрыгивали» через автохтонные популяции, не смешиваясь с ними [Narasimhan et al., 2019, Suppl. materials, p. 235]. Итак, Т.А. Чикишева была совершенно права, когда писала, что на фоне явной протоморфности южной евразийской формации «влияние миграционных импульсов на формирование антропологического состава алтае-саянского населения было несколько преувеличено» [2012, c. 180] (новую сводку фактов, касающихся окуневцев, см.: [Козинцев, 2020]).
То же самое, судя по всему, относится и к северной формации. Особых оснований считать, что ее носители пришли в Западную Сибирь из лесной зоны Северо-Восточной Европы, я не вижу. Вопрос о соотношении северной формации с уральской расой в ее традиционном широком понимании на рассмотренном материале не может быть решен. Во-первых, специфическое сходство обитателей лесов Северо-Восточной Европы с какой-либо из двух евразийских формаций Сибири установить не удается. Во-вторых, если уче сть высокую степень биологического своеобразия уралоязычных групп по обе стороны Урала (см. выше), то поиски истоков уральского единства на столь широкой территории в мезолите или неолите обречены на неудачу. Нет сомнений, что уральская прародина располагалась в таежной зоне Западной Сибири, частично захватывая Северное Приуралье [Напольских, 1997, с. 132, 140; Janhunen, 2009]. Ранние краниологические материалы оттуда очень скудны. В более западные районы лесной зоны, в частно сти в Поволжье и Прибалтику, уралоязычные (финно-угорские) группы продвинулись не раньше II тыс. до н.э. [Напольских, 1997, с. 125, 197–198; Janhunen, 2009].
На юг, в лесостепную зону Западной Сибири, уральцы проникли тоже поздно, о чем свидетельствуют краниоскопические данные, особенно чувствительные к присутствию уральского компонента [Громов, Моисеев, 2004; Моисеев, 2006]. Ни оку-невцы, ни люди, захороненные в разновременных погребениях Сопки, ни европеоиды Южной Сибири (афанасьевцы, андроновцы, карасукцы, ирмен-цы, тагарцы) по сочетанию краниометрических и краниоскопических характеристик не обнаруживают уральских черт [Козинцев, 2004]. Такие черты впервые проявляются в серии из «андроноид-ных» погребений могильника Еловка II в Томском Приобье [Там же], и это наиболее раннее свидетельство проникновения уральского компонента из тайги в подтаежную зону Западной Сибири в финале бронзового века.
Происхождение людей, связанных с чемурчек-ской культурой, остается загадочным. Генетические исследования выявляют у них сложную смесь западных, восточных и южных компонентов, причем один из главных, а именно ANE, мог быть получен от носителей ботайской культуры [Jeong et al., 2020; Wang et al., 2021]. На графике (см. рис. 1) два чемурчекских черепа (№ 42) попадают в прибайкальскую группировку, но ввиду метисности чемурчекцев этому не стоит придавать значения.
Серия, в которую включены ботайские черепа (№ 24), располагается «западнее» и образует пару с группой из погребений культуры ямочно-гребенчатой керамики Европейской России, что подтверждается археологическими данными о родстве этих культур [Мосин, 2003, с. 97–98]. Однако предположение о их связи с финно-уграми [Там же] противоречит многочисленным фактам, свидетельствующим о более позднем появлении финно-угорских племен к западу от Приуралья (см. выше).
Выводы
-
1. Краниометрические данные подтверждают вывод генетиков о раннем (в эпоху неолита) проникновении восточного компонента из Восточного Забайкалья в Прибайкалье, где он смешался с автохтонным (палеосибирским).
-
2. Северная и южная евразийские формации, а также группы, близкие к ним, занимают промежуточное положение между прибайкальской (палеосибирской) группировкой и европейскими группами с ослабленной выраженностью западных черт. Роль северной формации в генезисе уральской общности неясна.
-
3. Восточно-западный градиент генетиче ской и краниологической изменчивости на территории Северной Евразии свидетельствует, судя по всему, не о смешении, масштабы которого в эпохи неолита и ранней бронзы были еще невелики (во всяком случае, в центральной и западной частях рассмотренного региона), а о незавершенности процесса дифференциации бореального ствола.
-
4. Группы, которые можно считать метисными (чаахольская и елунинская), составляют незначительное меньшинство и противостоят как автохтонным, относящимся к палеосибирскому типу и обеим евразийским формациям, так и мигрантным, в частности афанасьевским.
Я признателен Д.В. Пежемскому за предоставление неопубликованных данных о серии из могильника Шаманка и Т.А. Чикишевой за ценные соображения.
Список литературы Основные направления популяционной динамики в Северной Евразии от мезолита до эпохи ранней бронзы (по данным краниологии и генетики)
- Алексеев В.П. О происхождении древнейшего европеоидного населения Минусинской котловины // Вопросы этнографии Хакасии. – Абакан: ХакНИИЯЛИ, 1981. – С. 4–10.
- Алексеев В.П., Гохман И.И. Результаты экспертизы надежности краниометрических показателей антропологических материалов из могильника на Южном Оленьем острове Онежского озера // Проблемы древнего и современного населения севера Евразии. – Л.: Наука, 1984. – С. 155–188.
- Алексеев В.П., Гохман И.И., Тумэн Д. Краткий очерк палеоантропологии Центральной Азии (каменный век – эпоха раннего железа) // Археология, этнография и антропология Монголии. – Новосибирск: Наука, 1987. – С. 208–241.
- Алексеев В.П., Дебец Г.Ф. Краниометрия: Методика антропологических исследований. – М.: Наука, 1964. – 128 с.
- Багашёв А.Н. Антропологические общности, их систематика и особенности расообразовательных процессов // Очерки культурогенеза народов Западной Сибири. – Томск: Изд-во Том. гос. ун-та, 1998. – Т. 4: Расогенез коренного населения. – С. 303–327.
- Бунак В.В. Человеческие расы и пути их образования // СЭ. – 1956. – № 1. – С. 86–105.
- Герасимова М.М., Васильев С.В., Фризен С.Ю., Боруцкая С.Б., Жамбалтарова Е.Д. Палеоантропологическое исследование Фофоновского могильника (Забайкалье) эпохи неолита – энеолита // Вестн. антропологии. – 2010. – № 18. – С. 113–127.
- Гохман И.И. Происхождение центральноазиатской расы в свете новых палеоантропологических материалов // Исследования по палеоантропологии и краниологии СССР. – Л.: Наука, 1980. – С. 5–34. – (Сб. МАЭ; т. 36).
- Гохман И.И. Новые палеоантропологические материалы эпохи мезолита в Каргаполье // Проблемы древнего и современного населения севера Евразии. – Л.: Наука, 1984. – С. 6–27.
- Громов А.В. Происхождение и связи населения окуневской культуры // Окуневский сборник. – СПб.: Петро-РИФ, 1997. – С. 301–358.
- Громов А.В., Моисеев В.Г. Краниоскопия населения Западной и Южной Сибири: география и хронология // Расы и народы. – М.; Л.: Наука, 2004. – Т. 30. – С. 216–248.
- Дебец Г.Ф. Палеоантропология СССР. – М.; Л.: Изд-во АН СССР, 1948. – 391 с. – (ТИЭ; т. 4).
- Дебец Г.Ф. Антропологические исследования в Камчатской области. – М.; Л.: Изд-во АН СССР, 1951. – 263 с. – (ТИЭ; т. 17).
- Денисова Р.Я. Антропология древних балтов. – Рига: Зинатне, 1975. – 402 с.
- Козинцев А.Г. Этническая краниоскопия: Расовая изменчивость швов черепа современного человека. – Л.: Наука, 1988. – 168 с.
- Козинцев А.Г. Кеты, уральцы, «американоиды»: интеграция краниометрических и краниоскопических данных // Палеоантропология, этническая антропология, этногенез: К 75-летию Ильи Иосифовича Гохмана. – СПб.: МАЭ РАН, 2004. – С. 172–185.
- Козинцев А.Г. Расовая классификация в свете новых генетических данных // Радловский сборник: научные исследования и музейные проекты МАЭ РАН в 2012 г. – СПб.: МАЭ РАН, 2013. – С. 64–71.
- Козинцев А.Г. Европеоиды, монголоиды, австралоиды: стадиальность или метисация? // Радловский сборник: научные исследования и музейные проекты МАЭ РАН в 2013 г. – СПб.: МАЭ РАН, 2014. – С. 25–33.
- Козинцев А.Г. О некоторых аспектах статистического анализа в краниометрии // Радловский сборник: научные исследования и музейные проекты МАЭ РАН в 2015 г. – СПб.: МАЭ РАН, 2016. – С. 381–390.
- Козинцев А.Г. Происхождение окуневского населения Южной Сибири по данным физической антропологии и генетики // Археология, этнография и антропология Евразии. – 2020. – Т. 48, № 4. – С. 135–145.
- Кондукторова Т.С. Антропология населения Украины мезолита, неолита и эпохи бронзы. – М.: Наука, 1973. – 127 с.
- Мамонова Н.Н. К вопросу о межгрупповых различиях в неолите Прибайкалья // Вопр. антропологии. – 1983. – № 71. – С. 88–103.
- Моисеев В.Г. Северная Евразия: языковая дифференциация и данные физической антропологии // Археология, этнография и антропология Евразии. – 2001. – № 4. – С. 154–159.
- Моисеев В.Г. Краниоскопическая характеристика населения Западной и Южной Сибири скифского времени // Археология, этнография и антропология Евразии. – 2006. – № 1. – С. 145–152.
- Моисеев В.Г., Козинцев А.Г. Антропологическое своеобразие уралоязычных народов // СЭ. – 1998. – № 2. – С. 140–150.
- Мосин В.С. Энеолитическая керамика Урало-Иртышского междуречья. – Челябинск: Изд-во Юж.-Урал. гос. ун-та, 2003. – 220 с.
- Напольских В.В. Введение в историческую уралистику. – Ижевск: Удм. ин-т истории, языка и лит-ры, 1997. – 268 с.
- Николаева Н.А. Древнейшая история Предкавказья в свете концепции индоевропейских миграций (часть 1) // Oriental Studies. – 2019а. – Т. 12, № 3. – С. 355–366.
- Николаева Н.А. Древнейшая история Предкавказья в свете концепции индоевропейских миграций (часть 2) // Oriental Studies. – 2019б. – Т. 12, № 4. – С. 570–579.
- Потехина И.Н. Население Украины в эпохи неолита и раннего энеолита по антропологическим данным. – Киев: Ин-т археологии НАН Украины, 1999. – 217 с.
- Солодовников К.Н. Краниологические материалы из могильника андроновской культуры Фирсово XIV в свете проблем формирования населения Верхнего Приобья в эпоху бронзы // Изучение историко-культурного наследия народов Южной Сибири. – Горно-Алтайск: АКИН, 2005. – С. 47–75.
- Солодовников К.Н. Антропологические материалы афанасьевской культуры: к проблеме происхождения // Вестн. антропологии. – 2009. – № 17. – С. 117–135.
- Солодовников К.Н., Багашёв А.Н., Савенкова Т.М. Ареалы антропологических общностей населения неолита юга Западной и Средней Сибири // Вестн. Том. гос. ун-та. Сер.: История. – 2020. – № 68. – С. 158–167.
- Солодовников А.Н., Багашёв А.Н., Тур С.С., Громов А.В., Нечвалода А.И., Кравченко Г.Г. Источники по палеоантропологии неолита – энеолита Среднего Прииртышья // Вестн. археологии, антропологии и этнографии. – 2019. – № 3. – С. 116–136.
- Солодовников К.Н., Тумен Д., Эрдэнэ М. Краниология чемурчекской культуры Западной Монголии // Древности Восточной Европы, Центральной Азии и Южной Сибири в контексте связей и взаимодействий в евразийском культурном пространстве (новые данные и концепции). – СПб: ИИМК РАН, 2019. – Т. 2. – С. 79–81.
- Солодовников К.Н., Тур С.С. Краниологические материалы елунинской культуры эпохи ранней бронзы Верхнего Приобья // Кирюшин Ю.Ф., Грушин С.П., Тишкин А.А. Погребальный обряд населения эпохи ранней бронзы Верхнего Приобья (по материалам грунтового могильника Телеутский Взвоз I). – Барнаул: Изд-во Алт. гос. ун-та, 2003. – С. 142–176.
- Солодовников К.Н., Тур С.С. К антропологии неолитического населения Барнаульского Приобья (по материалам могильника Фирсово XI) // Вестн. археологии, антропологии и этнографии. – 2017. – № 3. – С. 60–72.
- Тур С.С., Солодовников К.Н. Новые краниологические материалы из погребений каракольской культуры эпохи бронзы Горного Алтая // Изучение историко-культурного наследия народов Южной Сибири. – Горно-Алтайск: АКИН, 2005. – С. 35–47.
- Хохлов А.А. Морфогенетические процессы в Волго-Уралье в эпоху раннего голоцена (по краниологическим материалам мезолита – бронзового века). – Самара: Самар. гос. социал.-пед. ун-т, 2017. – 368 с.
- Хохлов А.А., Китов Е.П. Физический облик представителей ботайской энеолитической культуры в контексте проблемы формирования степного населения Казахстана // Казахское ханство в потоке истории: сб. науч. ст., посвящ. 550-летию образования Казахского ханства. – Алматы: Ин-т археологии им. А.Х. Маргулана, 2015. – С. 441–449.
- Чикишева Т.А. Динамика антропологической дифференциации населения юга Западной Сибири в эпохи неолита – раннего железа. – Новосибирск: Изд-во ИАЭТ СО РАН, 2012. – 467 с.
- Чикишева Т.А., Поздняков Д.В. Антропологические аспекты одиновской культуры (Западная Сибирь) // Археология, этнография и антропология Евразии. – 2019. – Т. 47, № 4. – С. 128–139.
- Чикишева Т.А., Поздняков Д.В. Заселение Барабинской лесостепи в эпоху неолита по антропологическим данным // Археология, этнография и антропология Евразии. – 2021. – Т. 49, № 1. – С. 133–145.
- Чикишева Т.А., Поздняков Д.В., Зубова А.В. Краниологические особенности палеопопуляции неолитического могильника Венгерово-2А в Барабинской лесостепи // Теория и практика археологических исследований. – 2015. – № 2. – С. 144–162.
- Biasutti R. Le razze e i popoli della terra. – Torino: Unione tipografi co-editrice torinese, 1941. – T. 1. – 825 p.
- Damgaard P., Martiniano R., Kamm J., Moreno-Mayar J.V., Kroonen G., Peyrot M., Barjamovic G., Rasmussen S., Zacho C., Baimukhanov N., Zaibert V., Merz V., Biddanda A., Merz I., Loman V., Evdokimov V., Usmanova E., Hemphill B., Seguin-Orlando A., Eylem Yediay F., Ullah I., Sjögren K.-G., Højholt Iversen K., Choin J., de la Fuente C., Ilardo M., Schroeder H., Moiseyev V., Gromov A., Polyakov A., Omura S., Yücel Senyurt S., Ahmad H., McKenzie C., Margaryan A., Hameed A., Samad A., Gul N., Hassan Khokhar M., Goriunova O., Bazaliiskii V., Novembre J., Weber A.W., Orlando L., Allentoft M.E., Nielsen R., Kristiansen K., Sikora M., Outram A.K., Durbin R., Willerslev E. The fi rst horse herders and the impact of early Bronze Age steppe expansions into Asia // Science. – 2018. – Vol. 360, iss. 6396. – URL: https://science.sciencemag.org/content/360/6396/eaar7711 (дата обращения: 10.07.2021).
- Flegontov P., Changmai P., Zidkova A., Logacheva M.D., Altinisik N.E., Flegontova O., Gelfand M.S., Gerasimov E.S., Khrameeva E.E., Konovalova O.P., Neretina T., Nikolsky Y.V., Starostin G., Stepanova V.V., Travinsky I.V., Tříska M., Tříska P., Tatarinova T.V. Genomic study of the Ket: A Paleo-Eskimo-related ethnic group with significant Ancient North Eurasian ancestry // Scientifi c Rep. – 2016. – N 6. – Article number 20768. – URL: https://doi.org.10.1038/srep20768 (дата обращения: 10.07.2021).
- Fu Q., Posth C., Hajdinjak M., Petr M., Mallick S., Fernandes D., Furtwängler A., Haak W., Meyer M., Mittnik A., Nickel B., Peltzer A., Rohland N., Slon V., Talamo S., Lazaridis I., Lipson M., Mathieson I., Schiffels S., Skoglund P., Derevianko A., Drozdov N., Slavinsky V., Tsybankov A., Cremonesi R., Mallegni F., Gély B., Vacca E., Morales González M., Straus L., Neugebauer-Maresch C., Teschler-Nicola M., Constantin S., Moldovan O., Benazzi S., Peresani M., Coppola D., Lari M., Ricci S., Ronchitelli A., Valentin F., Thevenet C., Wehrberger K., Grigorescu D., Rougier H., Crevecoeur I., Flas D., Semal P., Mannino M., Cupillard C., Bocherens H., Conard N., Harvati K., Moiseyev V., Drucker D., Svoboda J., Richards M., Caramelli D., Pinhasi R., Kelso J., Patterson N., Krause J., Pääbo S., Reich D. The genetic history of Ice Age Europe // Nature. – 2016. – Vol. 534, iss. 7606. – P. 200–205.
- Haak W., Lazaridis I., Patterson N., Rohland N., Mallick S., Llamas B., Brandt G., Nordenfelt S., Harney E., Stewardson K., Fu Q., Mittnik A., Bánffy E., Economou C., Francken M., Friederich S., Garrido Pena R., Hallgren F., Khartanovich V., Khokhlov A., Kunst M., Kuznetsov P., Meller H., Mochalov O., Moiseyev V., Nicklisch N., Pichler S.L., Risch R., Rojo Guerra M.A., Toth C., Széczényi-Nagy A., Wahl J., Meyer M., Krause J., Brown D., Anthony D., Cooper A., Alt K.W., Reich D. Massive migration from the steppe was a source for Indo-European languages in Europe // Nature. – 2015. – Vol. 522, iss. 7555. – P. 207–211.
- Hammer Ø. PAST – Paleontological Statistics. – Oslo: Univ. of Oslo, 2012. – 229 p. – URL: https://www.nhm.uio.no/english/research/infrastructure/past/ (дата обращения: 10.07.2021).
- Janhunen J. Proto-Uralic – what, where, and when? // Suomalais-ugrilaisen seuran toimituksia: (Mémoires de la Société Finno-Ougrienne). – 2009. – Vol. 258. – P. 156–167.
- Jeong C., Balanovsky O., Lukianova E., Kahbatkyzy N., Flegontov P., Zaporozhchenko V., Immel A., Wang C., Ixan O., Khussainova E., Bekmanov B., Zaibert V., Lavryashina M., Pocheshkhova E., Yusupov Y., Agdzhoyan A., Koshel S., Bukin A., Nymadawa P., Turdikulova S., Dalimova D., Churnosov M., Skhalyakho R., Daragan D., Bogunov Y., Bogunova A., Shtrunov A., Dubova N., Zhabagin M., Yepiskoposyan L., Churakov V., Pislegin N., Damba L., Saroyants L., Dibirova K., Atramentova L., Utevska O., Idrisov E., Kamenshchikova E., Evseeva I., Metspalu M., Outram A., Robbeets M., Djansugurova L., Balanovska E., Schiffels S., Haak W., Reich D., Krause J. The genetic history of admixture across Inner Eurasia // Nature Ecology and Evolution. – 2019. – Vol. 3, iss. 6. – P. 966–976.
- Jeong C., Wang K., Wilkin S., Treal Taylor W., Miller B., Bemmann J., Stahl R., Chiovelli C., Knolle F., Ulziibayar S., Khatanbaatar D., Erdenebaatar D., Erdenebat U., Ochir A., Ankhsanaa G., Vanchigdash C., Ochir B., Munkhbayar C., Tumen D., Kovalev A., Kradin N., Bazarov B., Miyagashev D., Konovalov P., Zhambaltarova E., Ventresca Miller A., Haak W., Schiffels S., Krause J., Boivin N., Erdene M., Hendy J., Warinner C. A dynamic 6,000-year genetic history of Eurasia’s Eastern steppe // Cell. – 2020. – Vol. 183, iss. 4. – P. 890–904.
- Jones E., Zarina G., Moiseyev V., Lightfoot E., Nigst P., Manica A., Pinhasi R., Bradley D. The Neolithic transition in the Baltic was not driven by admixture with Early European Farmers // Current Biology. – 2017. – Vol. 27, iss. 4. – P. 576–582.
- Kılınç G.M., Kashuba N., Koptekin D., Bergfeldt N., Dönertaş H., Rodriguez-Varela R., Shergin D., Ivanov G., Kichigin D., Pestereva K., Volkov D., Mandryka P., Kharinskii A., Tishkin A., Ineshin E., Kovychev E., Stepanov A., Dalén L., Günther T., Kırdök E., Jakobsson M., Somel M., Krzewińska M., Stora J., Götherström A. Human population dynamics and Yersinia pestis in ancient northeast Asia // Science Advances. – 2021. – Vol. 7, iss. 2. – URL: https://advances.sciencemag.org/content/7/2/eabc4587 (дата обращения: 10.07.2021).
- Kozintsev A. Ethnic epigenetics: a new approach // Homo. – 1992. – Vol. 43, iss. 3. – P. 213–244.
- Lazaridis I., Belfer-Cohen A., Mallick S., Patterson N., Cheronet O., Rohland N., Bar-Oz G., Bar-Yosef O., Jakeli N., Kvavadze E., Lordkipanidze D., Matzkevich Z., Meshveliani T., Culleton B., Kennett D., Pinhasi R., Reich D. Paleolithic DNA from the Caucasus reveals core of West Eurasian ancestry // bioRxiv. – 2018. – URL: https://doi.org/10.1101/423079 (дата обращения: 10.07.2021).
- Mao X., Zhang H., Qiao S., Liu Y., Chang F., Xie P., Zhang M., Wang T., Li M., Cao P., Yang R., Liu F., Dai Q., Feng X., Ping W., Lei C., Olsen J., Bennett E., Fu Q. The deep population history of northern East Asia from the Late Pleistocene to the Holocene // Cell. – 2021. – Vol. 184, iss. 12. – P. 3254–3266. – URL: https://doi.org/10.1016/j.cell.2021.04.040 (дата обращения: 10.07.2021).
- Mittnik A., Wang C., Pfrengle S., Daubaras M., Zariņa G., Hallgren F., Allmäe R., Khartanovich V., Moiseyev V., Tõrv M., Furtwängler A., Valtueña A., Feldman M., Economou C., Oinonen M., Vasks A., Balanovska E., Reich D., Jankauskas R., Haak W., Schiffels S., Krause J. The genetic prehistory of the Baltic Sea Region // Nature Communications. – 2018. – Vol. 9, iss. 1. – Article number 442. – URL: https://doi.org/10.1038/s41467-018-02825-9 (дата обращения: 10.07.2021).
- Narasimhan V., Patterson N., Moorjani P., Rohland N., Bernardos R., Mallick S., Lazaridis I., Nakatsuka N., Olalde I., Lipson M., Kim A., Olivieri L., Coppa A., Vidale M., Mallory J., Moiseyev V., Kitov E., Monge J., Adamski N., Alex N., Broomandkhoshbacht N., Candilio F., Callan K., Cheronet O., Culleton B., Ferry M., Fernandes D., Freilich S., Gamarra B., Gaudio D., Hajdinjak M., Harney É., Harper T., Keating D., Lawson M., Mah M., Mandl K., Michel M., Novak M., Oppenheimer J., Rai N., Sirak K., Slon V., Stewardson K., Zalzala F., Zhang Z., Akhatov G., Bagashev A., Bagnera A., Baitanayev B., Bendezu-Sarmiento J., Bissembaev A., Bonora G., Chargynov T., Chikisheva T., Dashkovskiy P., Derevianko A., Dobeš M., Douka K., Dubova N., Duisengali M., Enshin D., Epimakhov A., Fribus A., Fuller D., Goryachev A., Gromov A., Grushin S., Hanks B., Judd M., Kazizov E., Khokhlov A., Krygin A., Kupriyanova E., Kuznetsov P., Luiselli D., Maksudov F., Mamedov A., Mamirov T., Meiklejohn C., Merrett D., Micheli R., Mochalov O., Mustafokulov S., Nayak A., Pettener D., Potts R., Razhev D., Rykun M., Sarno S., Savenkova T., Sikhymbaeva K., Slepchenko S., Soltobaev O., Stepanova N., Svyatko S., Tabaldiev K., Teschler-Nicola M., Tishkin A., Tkachev V.,Vasilyev S., Velemínský P., Voyakin D., Yermolayeva A., Zahir M., Zubkov V., Zubova A., Shinde V., Lalueza-Fox C., Meyer M., Anthony D., Boivin N., Thangaraj K., Kennett D., Frachetti M., Pinhasi R., Reich D. The formation of human populations in South and Central Asia // Science. – 2019. – Vol. 365, iss. 6457. – P. 1002–1007. – URL: https://doi.org/10.1126/science.aat7487 (дата обращения: 10.07.2021).
- Raghavan M., Skoglund P., Graf K.E., Metspalu M., Albrechtsen A., Moltke I., Rasmussen S., Stafford T.W., Orlando L., Metspalu E., Karmin M., Tambets K., Rootsi S., Mägi R., Campos P.F., Balanovska E., Balanovsky O.P., Khusnutdinova E., Litvinov S., Osipova L.P., Fedorova S.A., Voevoda M.I., DeGiorgio M., Sicheritz-Ponten T., Brunak S., Demeshchenko S., Kivisild T., Villems R., Nielsen R., Jakobsson M., Willerslev E. Upper Paleolithic Siberian genome reveals dual ancestry of Native Americans // Nature. – 2014. – Vol. 505, iss. 7481. – P. 87–91.
- Sikora M., Pitulko V., Sousa V., Allentoft M., Vinner L., Rasmussen S., Margaryan A., Damgaard P.,Fuente C., Renaud G., Yang M., Fu Q., Dupanloup I., Giampoudakis K., Nogués-Bravo D., Rahbek C., Kroonen G., Peyrot M., McColl H., Vasilyev S., Veselovskaya E., Gerasimova M., Pavlova E., Chasnyk V., Nikolskiy P., Gromov A., Khartanovich V., Moiseyev V., Grebenyuk P., Fedorchenko A., Lebedintsev A., Slobodin S., Malyarchuk B., Martiniano R., Meldgaard M., Arppe L., Palo J., Sundell T., Mannermaa K., Putkonen M., Alexandersen V., Primeau C., Baimukhanov N., Malhi R., Sjögren K., Kristiansen K., Wessman A., Sajantila A., Mirazon Lahr M., Durbin R., Nielsen R., Meltzer D., Excoffier L., Willerslev E. The population history of Northeastern Siberia since the Pleistocene // Nature. – 2019. – Vol. 570, iss. 7760. – P. 182–188.
- Wang C., Yeh H., Popov A., Zhang H., Matsumura H., Sirak K., Cheronet O., Kovalev A., Rohland N., Kim A., Mallick S., Bernardos R., Tumen D., Zhao J., Liu Y.,Liu J., Mah M., Wang K., Zhang Z., Adamski N., Broomandkhoshbacht N., Callan K., Candilio F., Duffett Carlson K., Culleton B., Eccles L., Freilich S., Keating D., Lawson A., Mandl K., Michel M., Oppenheimer J., Özdoğan T., Stewardson K., Wen S., Yan S., Zalzala F., Chuang R., Huang C., Looh H., Shiung C., Nikitin Y., Tabarev A., Tishkin A., Lin S., Sun Z., Wu X., Yang T., Hu X., Chen L., Du H., Bayarsaikhan J., Mijiddorj E., Erdenebaatar D., Iderkhangai T., Myagmar E., Kanzawa-Kiriyama H., Nishino M., Shinoda K., Shubina O., Guo J., Cai W., Deng Q., Kang L., Li D., Li D., Lin R., Nini, Shrestha R., Wang L., Wie L., Xie G., Yao H., Zhang M.,He G., Yang X., Hu R., Robbeets M., Schiffels S., Kennett D., Jin L., Li H., Krause J., Pinhasi R., Reich R. Genomic insights into the formation of human populations in East Asia // Nature. – 2021. – Vol. 591, iss. 7850. – P. 413–419. – URL: https://doi.org/10.1038/s41586-021-03336-2 (дата обращения: 10.07.2021).
- Yu H., Spyrou M., Karapetian M., Shnaider S., Radzevičiūte R., Nägele K., Neumann G., Penske S., Zech J., Lucas M., LeRoux P., Roberts P., Pavlenok G., Buzhilova A., Posth C., Jeong C., Krause J. Paleolithic to Bronze Age Siberians reveal connections with fi rst Americans and across Eurasia // Cell. – 2020. – Vol. 181, iss. 6. – P. 1232–1245.
