Особенности адаптивных реакций гибридов подсолнечника на условия экстремальной засухи 2010 года на Европейской территории России

Автор: Дьяков А.Б., Борсуков А.А.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (159-160), 2014 года.
Бесплатный доступ
Испытания 12 гибридов подсолнечника провели в основных зонах его возделывания на европейской части России в 18 пунктах в остро засушливом 2010 г. и в 22 пунктах в 2011 г. Средние по опытам урожаи варьировали от 11 до 40 ц/га в 2010 г. и от 16 до 45 ц/га в 2011 г. Определяли параметры: средний урожай i-го гибрида а ь коэффициент вариации его урожаев CV средовое стандартное отклонение Si, эффект взаимодействия генотип-среда W i (эковалента), коэффициент регрессии урожаев на индексы среды bi, дисперсию регрессионных остатков S 2di, степень симилярности (однотипности) реакций гибридов оценивали коэффициентом детерминации СД Ъ урожаев индексами среды, по величинам аЈ и b вычисляли математические ожидания урожаев i- го генотипа при минимальном и максимальном индексах для оценок его устойчивости к стрессам и потенциала урожайности. По сравнению с 2011 г. в засушливом 2010 были выше оценки S i и CV i, но ниже а ь W i и S 2di. Такое снижение специфичности откликов гибридов на вариацию уровня главного стресса обусловило высокую - 90,6 до 98,3 % - симилярность их реакций. В более комфортных условиях 2011 г. были повышены не только оценки а ъ но и W i и S 2d при низких S i и CV i. При этом возросшая специфичность откликов гибридов была обусловлена их реакциями на качественно иные стрессы, связанные с повышенной увлажненностью территорий. Изученные гибриды классифицированы по сочетанию степени их устойчивостей к стрессам 2010 и 2011 гг. и реализации потенциалов урожайности в эти годы. Установлены и объяснены низкая эквивалентность оценок генотипов параметрами Si и CVi, но высокая - оценками S i и b i. Описан способ коррекции оценок S i. Показано, что на степень сими- лярности реакций генотипов отрицательно влияют значения S 2di и положительно - величины b i.
Подсолнечник, гибриды, реакция на стрессы разных лет, потенциал урожайности, параметры стабильности
Короткий адрес: https://sciup.org/142151200
IDR: 142151200 | УДК: 581.5:633.854.78(470)
Specifics of adaptive responses of sunflower hybrids on extreme drought conditions of 2010 in European territory of Russia
An ecological testing of 12 sunflower hybrids was conducted in its main growing areas of European part of Russia in 18 locations in the extremely dry 2010 and in 22 locations in 2011. The average tested yield was from 11 till 40 centners/ha in 2010 and from 16 till 45 centners/ha in 2011. We evaluated the following parameters: an average yield of hybrid i - а ъ a variation coefficient of its yield - CVi, a percentage and an environmental standard deviation - Si, an effect of interaction «genotype-environment» W i (ecovalence), a regression coefficient yields on an environmental index bi, a dispersion of regression residuals S 2di, a similarity (uniformity) degrees of hybrids reactions were evaluated by a coefficient of determination Cd i, a percentage of yields by environmental index, a mathematical expectations of yields of genotype i were calculated by using parameters а and bi within minimum and maximum indexes for its tolerance assessments to stresses and yield potential. As compared with 2011 in dry 2010 S ; and CV ; were higher, but a, W, and S 2d,. were lower. Such decrease of responses specificity of hybrids on a variation of the main stress degree determined high similarity of their reactions, from 90.6 till 98.3%. In more comfortable conditions of 2011 year there were not only increased assessments of the parameter а ъ but also for parameters Wi and S 2di within low Si and CVi. At the same time an increased specificity of hybrids responses was determined by their reactions on qualitatively different stresses linked with higher moisture of the territories. The tested hybrids were classified by a combination of degree of their resistance to stresses of 2010 and 2011 and yield potentials realization in these years. It was determined and explained that a lower equivalence of genotypes assessments by parameters Si and CV b but higher - by parameters Si and bi. It was described a method of assessments corrections Si. It was shown that values S 2^ negatively influence on similarity genotypes reactions degree and values bi - positively.
Текст научной статьи Особенности адаптивных реакций гибридов подсолнечника на условия экстремальной засухи 2010 года на Европейской территории России
Введение. Описывая достижения за 50 лет селекции растений, генетики и цитологии P.S. Baenziger с соавторами [1] основное внимание уделили успехам молекулярной биологии, которая сделала доступными для селекционеров многие новые признаки, находящиеся под простым генетическим контролем. При этом они признают, что и сегодня селекция ведётся в первую очередь на сложные полигенные признаки адаптивность и урожайность, для улучшения которых селекционеры продолжают использовать эмпирические способы определения целей селекции, подбора родителей для гибридизации, выявления стабильно урожайных в агроценозах генотипов среди как растений гибридных популяций, так и созданных сортов и гибридов. Особенно много сложных, нерешённых проблем ограничивают эффективность селекции растений на повышение уровня агроэкологической адаптивности [2; 3].
В основополагающем обзоре о значении взаимодействия генотип – среда R.W. Allard и A.D. Bradshaw [4] показали, что при селекции на адаптивность к предсказуемым условиям среды в случаях взаимодействий генотип – местообитание и генотип – агротехника селекционер имеет возможность планировать эмпирические испытания генотипов. Намного труднее их оценивать на адаптивность к непредсказуемым колебаниям условий внешней среды при взаимодействии генотип – погодные условия. S.A. Eberhart и W.A. Russell [5] пришли к выводу о неэффективности селек- ции на узкую адаптивность генотипов к субрегионам со всеми сходными климатическими и почвенными условиями потому, что непредсказуемая вариация погодных условий все равно потребует широкой адаптивности создаваемых сортов и гибридов.
Изучая в штате Калифорния (США) реакцию генотипов подсолнечника на сроки сева и густоты посевов в разные годы B.H. Beard и Shu Geng [6] выявили большую средовую вариацию оценок урожайности. Поскольку среди объектов их исследования были успешно возделывавшиеся в то время в разных условиях СССР, Аргентины, Австралии, США сорт Передовик и гибрид Romsun 52, авторы сделали вывод, что в основном разные генотипы подсолнечника реагируют симилярно (однообразно) на изменения внешних условий. В их опытах даже на фоне орошения при оптимальных густоте посевов и сроке сева средние урожаи в разные годы изменялись в два раза, причем в основном симилярными были реакции генотипов в неблагоприятный год, а в благоприятный проявилось значительное взаимодействие генотип – среда. В наших опытах [7] также изучалась реакция сортов и гибридов подсолнечника на различные густоты посевов в разные по количеству осадков годы. Усредненные по всем вариантам оценки урожайности оказались близкими в эти годы, хотя в одном случае продуктивность растений лимитировал дефицит влаги, а в другом – поражение растений фомопси-сом. Вследствие генотипических различий изученных сортов и гибридов по длительности вегетации и восприимчивости к фомопси-су их реакция на густоты посевов резко изменилась. Это проявилось в изменении рангов генотипов не только по урожайности, но и по величинам параметров стабильности и адаптивности. Описанные результаты дали основание полагать, что адаптивные реакции гибридов подсолнечника на разнообразие природных условий разных регионов России также могут различаться в экстремально засушливый и обычный по метеорологическим условиям годы.
Материалы и методы. Адаптивные реакции 12 простых межлинейных гибридов подсолнечника изучались в опытах экологических испытаний в разных почвенно-климатических зонах европейской территории России: от предгорий Кавказа на юге до Тамбовской области в северной части ареала возделывания масличного подсолнечника и от Белгородской области на западе до Башкортостана и Оренбургской области на востоке. Всего было проведено 18 опытов в 2010 г. и 22 опыта в 2011 г. О разной степени благоприятности условий для подсолнечника в эти годы можно судить по величинам средней по всем пунктам и всем гибридам урожайности, которая составила в экстремально засушливом 2010 24,4 ц/га, а в 2011 г. – 32,0 ц/га.
Опыты в эти годы во всех зонах проводились по одинаковой методике: повторность 3-кратная, делянки 4-рядные без защитных рядов, междурядья 0,7 м, учетная площадь 18,2 м2. Сев производили сеялкой BAURAL с нормами высева на густоту 56 тыс. раст./га в засушливых зонах (Поволжье и Южный Урал) и 59,5 тыс. раст./га в остальных регионах. Убирали урожай комбайном ZURN-150, массу семянок пересчитывали на 7%-ную влажность. Оценки чисел дней от даты сева до цветения гибридов приведены в среднем по данным учетов в отдельных опытах. В разные годы и в разных зонах этот показатель изменялся, но ранги гибридов по продолжительности вегетации сохранялись во всех опытах.
Для вычислений показателей абсолютной (S i ) и относительной (CV i , %) экологической изменчивости оценок урожайности гибридов использовали обычные формулы [8]. Величины вкладов каждого i-го генотипа в общую дисперсию взаимодействия генотип – среда (W i ) оценивали по формуле G. Wricke [9]. Параметры регрессий оценок урожайности на индексы среды (b i и S2d i ) вычисляли по методу Эберхарта и Расселла [5]. Степень симилярности адаптивных реакций гибридов оценивали коэффициентами детерминации (Cd i , %) оценок урожайности индексами среды, которые вычисляли по [10].
Результаты. По продолжительности вегетации изученные гибриды подсолнечника различаются в небольшой степени. Максимальная разница между датами цветения составила пять суток, а растения шести из 12 гибридов зацветают одновременно (табл. 1).
Таблица 1
Влияние экстремальной засухи 2010 г. на абсолютную (стандартные отклонения S i ) и относительную (коэффициенты вариации CV i , %) вариабельность оценок урожайности (а i , ц/га) гибридов подсолнечника по данным испытаний на европейской территории России
|
№№ |
Гибрид |
Дней от сева до цветения |
Средний урожай, ц/га |
Вариабельность урожаев |
||||
|
2010 г. |
2011 г. |
абсолютная, S i |
относительная, CV i , % |
|||||
|
2010 г. |
2011 г. |
2010 г. |
2011 г. |
|||||
|
1 |
Савинка |
75 |
24,8 |
27,6 |
9,32 |
6,52 |
37,6 |
23,7 |
|
2 |
НК Роки |
76 |
25,0 |
33,3 |
9,22 |
6,57 |
36,8 |
19,7 |
|
3 |
Ригасол ОР |
76 |
21,4 |
29,3 |
9,01 |
6,98 |
42,0 |
23,8 |
|
4 |
Тристан |
78 |
25,0 |
30,1 |
10,28 |
8,10 |
41,2 |
26,9 |
|
5 |
НК Ферти |
78 |
24,3 |
31,1 |
10,24 |
6,50 |
42,2 |
20,9 |
|
6 |
НК Делфи |
78 |
24,5 |
31,4 |
11,07 |
7,07 |
45,1 |
22,5 |
|
7 |
Тутти |
78 |
23,7 |
33,1 |
10,09 |
7,19 |
42,6 |
21,7 |
|
8 |
НК Конди |
78 |
26,9 |
34,2 |
11,53 |
7,74 |
42,9 |
22,6 |
|
9 |
НК Неома |
78 |
24,1 |
32,9 |
10,41 |
7,94 |
43,3 |
24,2 |
|
10 |
НК Брио |
79 |
25,1 |
33,4 |
9,97 |
7,98 |
39,7 |
23,9 |
|
11 |
Арена ПР |
79 |
22,9 |
32,3 |
9,04 |
7,84 |
39,5 |
24,3 |
|
12 |
НК Армони |
80 |
26,3 |
33,8 |
11,06 |
7,88 |
42,0 |
23,3 |
Несмотря на небольшие различия по числу суток от даты сева до цветения, по данным опытов 2011 г. обнаруживается тенденция повышения величин усреднённых по всем опытам оценок урожайности гибридов с увеличением продолжительности вегетации. Такая тенденция не проявляется при рассмотрении результатов испытания 2010 г. Обусловлено это тем, что в условиях экстремальной засухи урожайность в наибольшей степени снизилась у ряда гибридов с более поздним наступлением критического периода, начинающегося с фазы цветения. Такое снижение составило у гибридов Тутти 9,4 ц/га, Арена ПР – 9,4 ц/га, НК Неома – 8,8 ц/га. У самого же скороспелого из изученных генотипов гибрида Савинка это снижение составило в 2010 г. по сравнению с 2011 г. только 2,8 ц/га. В этих условиях он по средней урожайности почти сравнялся с одним из лучших гибридов НК Брио. Выделились как по потенциалу урожайности в благоприятных условиях, так и по способности обеспечивать сравнительно высокие урожаи при засухе НК Конди и НК Армони, а также более скороспелый НК Роки. В опытах 2010 г. самым низкоурожайным в среднем оказался гибрид Рига-сол ОР, а Тристан в этих условиях занял пятое место по продуктивности, снизив урожай лишь на 5,1 ц/га.
По данным таблицы 1 недостаточно четко выявляется повышенная устойчивость к засухе гибридов с более ранним наступлением критического периода, т. к. в ней представлены усредненные по всем опытам оценки урожайности. Поскольку и в 2010 г. в ряде опытов на юге России и на севере ареала возделывания подсолнечника средние по всем гибридам урожаи превышали 30 ц/га и достигали 40 ц/га, средние оценки урожайности гибридов не достаточно четко отражают реакции генотипов на условия экстремальной засухи. В таблице 2 представлены данные испытаний в пунктах в сильной и средней степени подвергшихся засухе в 2010 г., а в районе Саратова урожайность подсолнечника была снижена и в 2011 г. из-за крайне низких запасов влаги в почве и провокационных дождей в период вегетативного роста растений.
Таблица 2
Оценки урожайности (ц/га) гибридов подсолнечника в разные по погодным условиям годы в пострадавших от засухи 2010 г. пунктах испытаний
|
№ № |
Гибрид |
Миллерово Ростовской области |
Павловск Воронежской области |
Белгород |
Саратов |
||||
|
2010 г. |
2011 г. |
2010 г. |
2011 г. |
2010 г. |
2011 г. |
2010 г. |
2011 г. |
||
|
1 |
Савинка |
14,3 |
23,9 |
20,7 |
36,7 |
25,7 |
34,5 |
15,6 |
20,1 |
|
2 |
НК Роки |
13,1 |
31,9 |
16,4 |
37,6 |
22,7 |
34,7 |
17,4 |
20,9 |
|
3 |
Ригасол ОР |
10,2 |
26,0 |
15,3 |
30,8 |
19,2 |
29,7 |
15,8 |
14,3 |
|
4 |
Тристан |
11,6 |
21,1 |
15,7 |
37,3 |
23,7 |
34,1 |
16,3 |
16,8 |
|
5 |
НК Ферти |
8,6 |
27,6 |
12,9 |
36,6 |
23,8 |
30,6 |
13,6 |
19,3 |
|
6 |
НК Делфи |
8,6 |
26,9 |
13,3 |
27,4 |
21,8 |
29,7 |
15,8 |
19,0 |
|
7 |
Тутти |
9,4 |
30,3 |
14,8 |
37,4 |
23,4 |
29,6 |
14,9 |
22,7 |
|
8 |
НК Конди |
13,0 |
29,0 |
13,9 |
42,6 |
24,8 |
38,9 |
17,6 |
25,8 |
|
9 |
НК Неома |
10,2 |
28,7 |
16,1 |
38,4 |
20,7 |
38,7 |
17,6 |
17,1 |
|
10 |
НК Брио |
13,1 |
28,3 |
13,8 |
40,4 |
21,6 |
40,6 |
17,3 |
22,8 |
|
11 |
Арена ПР |
12,0 |
29,4 |
14,2 |
38,5 |
20,7 |
35,3 |
15,3 |
16,6 |
|
12 |
НК Армони |
10,9 |
30,7 |
13,7 |
35,6 |
20,9 |
35,3 |
20,7 |
22,0 |
|
Среднее |
11,3 |
27,8 |
15,1 |
36,6 |
22,4 |
34,3 |
16,5 |
19,8 |
|
При рассмотрении представленных в таблице 2 данных достаточно четко выявляется тенденция повышенной в условиях экстремальной засухи урожайности раньше зацветающих гибридов, хотя по потенциалу урожайности в благоприятных условиях они уступают более позднеспелым. Аналогичные результаты были получены исследователями разных стран, изучавших реакцию сортов и гибридов подсолнечника на изменения климатических и погодных условий, а также на разные режимы орошения. Например, В.А. Смирнова [11] пришла к выводу: «… при недостаточном увлажнении поздние сорта не имеют преимуществ перед ранними сортами, которые лучше используют запасы влаги. Следовательно, в районах, не обеспеченных в достаточном количестве влагой, экономически выгодней выращивать ранние и средние сорта подсолнечника». Она показала также, что поздние по ее классификации сорта, созревающие одновременно с сортом Передовик, снижают урожаи семянок и при недостатке тепла.
Вследствие меньшей степени снижения при дефиците влаги урожайности скороспелых генотипов и невысокого потенциала их продуктивности обычно наблюдается тем меньшая экологическая вариабельность урожаев, чем менее продолжителен период вегетации генотипа. В силу этой причины экологическая вариация Si скороспелых сортов сои в условиях Краснодара была в три раза меньше, чем у среднеспелых сортов [12], а в засушливых центральных районах южной Италии стабильность оценок урожайности уль-траскороспелого сорта подсолнечника Енисей была значительно выше, чем сортов Передовик, Смена и ВНИИМК 8931 [13]. Изученные нами гибриды подсолнечника в меньшей степени различаются по длительности вегетационного периода, однако и при их сравнении обнаруживается тенденция снижения величин стандартного отклонения Si с уменьшением числа дней от даты сева до фазы цветения (см. табл. 1). Эта тенденция менее четко проявилась в 2010 г. вследствие меньшего снижения урожайности при засухе у скороспелых генотипов.
Значения параметра абсолютной вариабельности S i оказались более высокими у всех гибридов в 2010 г. по сравнению с 2011 г. Это превышение составило от 15 % у Арены ПР и 27 % у Тристана до 57 % у НК Делфи и 58 % у НК Ферти. А поскольку средняя урожайность в условиях экстремальной засухи была сниженной, различия по нестабильности между 2010 и 2011 гг., оцененной параметром относительной вариабельности урожаев CV i , были еще более значительными, превысив 100 % у НК Ферти и НК Делфи (см. табл. 1).
Таблица 3
Влияние экстремальной засухи 2010 г. на общие оценки взаимодействия гибрид – местность (W i ), их линейные (b i ) и нелинейные (S2d i ) компоненты, детерминацию оценок урожайности индексами среды (Cd i , %) по данным испытаний гибридов подсолнечника на европейской территории России
|
№ № |
Гибрид |
Эковалента W i |
Коэффициент регрессии b i |
Остаточная дисперсия S2d i |
Коэффициент детерминации Cd i , % |
||||
|
2010 г. |
2011 г. |
2010 г. |
2011 г. |
2010 г. |
2011 г. |
2010 г. |
2011 г. |
||
|
1 |
Савинка |
146,99 |
276,29 |
0,90 |
0,81 |
8,74 |
11,98 |
90,6 |
73,2 |
|
2 |
НК Роки |
59,77 |
151,46 |
0,92 |
0,88 |
3,28 |
6,82 |
96,4 |
84,9 |
|
3 |
Ригасол ОР |
87,50 |
155,69 |
0,89 |
0,93 |
4,57 |
7,57 |
94,7 |
85,2 |
|
4 |
Тристан |
106,66 |
177,70 |
1,01 |
1,10 |
7,10 |
8,38 |
93,7 |
87,8 |
|
5 |
НК Ферти |
113,10 |
62,63 |
1,00 |
0,91 |
7,54 |
2,74 |
93,3 |
93,8 |
|
6 |
НК Делфи |
52,98 |
192,22 |
1,11 |
0,93 |
2,22 |
9,36 |
98,3 |
82,2 |
|
7 |
Тутти |
37,56 |
114,86 |
1,01 |
0,98 |
2,49 |
5,73 |
97,7 |
89,4 |
|
8 |
НК Конди |
114,19 |
87,77 |
1,15 |
1,08 |
5,37 |
4,03 |
96,2 |
93,6 |
|
9 |
НК Неома |
77,39 |
119,91 |
1,03 |
1,10 |
5,05 |
5,48 |
95,6 |
91,7 |
|
10 |
НК Брио |
55,71 |
181,54 |
0,99 |
1,08 |
3,71 |
8,78 |
96,5 |
86,9 |
|
11 |
Арена ПР |
43,74 |
106,20 |
0,91 |
1,09 |
2,00 |
4,88 |
97,7 |
92,4 |
|
12 |
НК Армони |
153,00 |
106,92 |
1,08 |
1,10 |
9,54 |
4,87 |
92,7 |
92,5 |
Примечание: все оценки коэффициента Cd i , % переходят 0,1 %-ный уровень существенности
При рассмотрении данных таблицы 3 следует учитывать, что приведенные в ней параметры взаимодействия генотип – среда Wi и S2di отражают не всю вариацию оценок урожайности, а только влияние на их изменчивость генотипических различий между гибридами по реакции на разнообразие метеорологических и поч- венных условий проведения опытов в разных природных зонах России. Если реакции i-го гибрида не отличаются от усредненных реакций всех гибридов испытания, то значения его параметров взаимодействия генотип – среда должны быть равными Wi = 0; S2di = 0; bi = 1,00; Cdi = 100 %. В 2010 г. наиболее близкими к этому были параметры гибрида Тутти, в 2011 г. – НК Ферти. Особенно специфично реагировали на условия испытаний самые скороспелые гибриды и зацветший позже всех гибрид НК Армони.
При сравнении величин приведенных в таблице 3 параметров, вычисленных по данным испытаний 2010 и 2011 гг., обнаруживаются значительные различия. У большинства гибридов значения эковалент W i и остаточных дисперсий S2d i оказались более высокими не в экстремально засушливый, а в обычный год, но диапазон изменчивости величин коэффициента регрессии оценок урожайности на индексы среды b i был больше в 2010 г., чем в 2011 г. Особенно четко проявились различия в эти годы значений коэффициента детерминации оценок урожайности индексами среды. Только у двух гибридов они оказались практически равными в эти годы, а десять гибридов реагировали более симилярно (однообразно) на вариацию условий среды в экстремально засушливом году, чем при испытаниях в обычный год. Если же учесть большое разнообразие экологических условий проведения испытаний на европейской территории России, то следует признать, что и в 2011 г. симилярность реакций гибридов оказалась также достаточно высокой. Тем более что величины Cd i от 73 до 85 % наблюдались у скороспелых генотипов, а это означает, что при включении в испытания только гибридов такого типа величины Cd i будут более высокими.
Обсуждение. Задачей селекции является не только создание высокоурожайных сортов и гибридов, но и обеспечение постоянства проявлений генотипов в разных условиях среды, их устойчивости к плохим и отзывчивости на хорошие условия. Разный смысл, вкладываемый в такие понятия, как стабильность, устой- чивость, адаптивность, толерантность к стрессам, а также множество предложенных для измерения этих свойств статистических параметров, не только приводят ученых в растерянность, какой из методов использовать в своей работе, но их применение может вводить в заблуждение вследствие отсутствия ясного понимания разных проявлений взаимодействия генотип – среда [14; 15]. В соответствии с разным пониманием того, какие особенности реакций генотипов на условия среды считать проявлением их стабильности, C.S. Lin, M.R. Binns и L.P. Lefkovitch [14], а также И. Пешек, И. Гартманн и И. Нидерле [15] классифицировали девять наиболее часто используемых параметров на три типа устойчивости. Однако B.S. Baenziger с соавторами [1] констатировали, что до сих пор в этом спорном вопросе сохраняется существенная неразбериха и между селекционерами нет консенсуса по вопросу о том, что является основой стабильности сорта и может ли стабильность быть целью селекции или нет. Примером этого может служить описанное А.Б. Дьяковым и М.В. Труновой [16] встречающееся в литературе разнообразие мнений о том, какую особенность генотипа отражает коэффициент регрессии оценок его урожайности на индексы среды bi. Разные авторы считают этот параметр мерой адаптивности, стабильности, отзывчивости на благоприятные условия, толерантности к стрессам или пластичности сорта (гибрида). У авторов, признающих параметры bi мерой пластичности генотипа, различные мнения о том, какая величина этого параметра свидетельствует о высокой пластичности: bi > 1,0; bi = 1,0 или bi < 1,0.
В первом томе монографии «Экология» Ю. Одум [17] отмечает важность учетов не только средних урожаев, но и их колебаний из года в год (с. 258), однако констатирует: «… на практике люди разных профессий (…) могут вкладывать в термин «стабильность» разный смысл, особенно при попытках оценить меру стабильности и выразить ее количественно. Соответственно много путаницы по этому вопросу и в литературе» (с. 66).
Примером этого могут служить различные рекомендации о возможностях использования параметров стандартное (среднее квадратичное) отклонение (Si или σ) и коэффициент вариации (CVi) для оценивания степени изменчивости признака. C.S. Lin с соавторами [14] и И. Пешек с соавторами [15] считают эти параметры эквивалентными при CVi < 20 %. Во многих руководствах по статистическим методам параметр Si (σ) считается непригодным для этой цели из-за зависимости его оценок от средней величины признака. Например, Г.Н. Зайцев [18] пишет: «Однако для сравнения вариабельности двух или более совокупностей величины сигмы не могут быть использованы, за исключением редкого случая, когда средние арифметические одинаковы или близки друг к другу. Во всех остальных случаях для сравнения совокупностей по их вариабельности необходимо вычислять коэффициент вариации». При этом считается, что оценки параметра CVi не зависят от средней величины признака. К. Мазер и Дж. Джинкс [19], напротив, считают, что при больших различиях средних значений признака генотипов их стабильность следует оценивать не коэффициентами вариации, а величинами дисперсий S2i. Дж. У. Снедекор (8) показал, что без учета средних величин и Si использование коэффициента вариации может приводить к заблуждениям. По этой причине С.М. Гершензон [20] пришел к выводу: «Нужно, однако, подчеркнуть, что не следует пользоваться коэффициентом изменчивости там, где можно обойтись средним квадратическим отклонением – в математическом отношении последнее значительно точнее характеризует изменчивость признака». С.С. Четвериков [21] также писал, что использование коэффициента вариации как меры изменчивости вызывает ряд серьезных возражений как с формально математической, так и с биологической стороны. Описанные проблемы использования абсолютных и относительных оценок вариабельности количественного признака потребовали провести анализ корреляционных связей величин Si и CVi изученных гибридов в 2010 и 2011 гг. с другими параметрами для обоснованной биологической интерпретации данных таблицы 1. Результаты этого анализа представлены в таблице 4.
Авторы классификации типов стабильности пришли к выводу об эквивалентности параметров S2 i и CV i на том основании, что при CV i < 20 % дисперсия логарифма величины признака приблизительно равна отношению дисперсии значения признака к квадрату его средней величины [14; 15]. Однако для характеристики генотипов важно не это равенство, а степень воспроизводимости и правильности оценок стабильности при использовании параметров S i или CV i . Представленные в таблице 4 данные свидетельствуют о далеко не полной степени их воспроизводимости: в 2010 и 2011 гг . пр и средних для всех гибридах оценках CV 41,2 и 23,1 % коэффициенты корреляции между значениями S i и CV i составили r = 0,696 и r = 0,691. Этим коэффициентам соответствует степень эквивалентности оценок стабильности этими параметрами только 48,4 и 47,7 %. Это подтверждает выводы Дж. У. Снедекора (8) о том, что широко применяемые в статистике вычисления относительных величин ведут к утрате некоторых сведений об изучаемых объектах и к недопустимым ошибкам вследствие этого, а также о том, что это в полной мере относится к такому показателю как коэффициент вариации.
Таблица 4
Коэффициенты корреляции между величинами параметров экологической вариабельности оценок урожайности гибридов подсолнечника и параметров взаимодействия генотип - среда при испытаниях в разные по количеству осадков годы
|
Параметры |
Стандартное отклонение, S i |
Коэффициент вариации, CV i , % |
||
|
2010 г. |
2011 г. |
2010 г. |
2011 г. |
|
|
a i , ц/га |
0,703** |
0,459 |
–0,021 |
–0,324 |
|
CV i , % |
0,696* |
0,691* |
1 |
1 |
|
W i |
0,276 |
–0,214 |
–0,070 |
0,304 |
|
b i |
0,990*** |
0,964*** |
0,725** |
0,546 |
|
S2d i |
0,225 |
–0,171 |
–0,100 |
0,324 |
|
Cd i , % |
0,087 |
0,496 |
0,309 |
–0,044 |
Коэффициенты корреляции переходят уровни существенности:* – 5 %-ный; ** – 1 %-ный;
*** – 0,1 %-ный
О различиях в оценках нестабильности гибридов параметрами S i и CV i можно судить и по другим данным таблицы 4. Оказалось, что со значениями параметра b i очень тесно и высоко значимо коррелируют оценки стандартного отклонения S i и в значительно меньшей мере – величины коэффициента вариации CV i . О степени этих различий надо судить по величинам коэффициента детерминации: коэффициентам r = 0,990 и r = 0,964 соответствует 98 и 93 % сопряженной изменчивости оценок параметров b i и S i , а при коэффициентах r = 0,725 и r =0,546 доля сопряженной вариации параметров b i и CV i составила лишь 53 и 30 %. Это также свидетельствует об утрате значительной части информации о средовой вариабельности оценок урожайности гибридов при использовании для этой цели коэффициента вариации CV i . Существенно различна также и теснота связей параметров S i с a i и CV i с a i .
По данным испытаний гибридов подсолнечника в 2010 и 2011 гг. коэффициенты корреляции между величинами коэффициентов вариации CVi и средними оценками урожайности аi оказались или близкими к нулю, или несущественно отличающимися от нуля (табл. 4). Это можно было бы считать свидетельством в пользу представления об отсутствии зависимости коэффициентов вариации от средней величины признака. Однако приведенные в разных публикациях факты и результаты собственных исследований свидетельствуют о том, что лишь в отдельных ситуациях коэффициент корреляции между этими параметрами может быть близким к нулю, а часто он значимо отрицателен. Отрицательные корреляции между коэффициентами вариации и средними величинами признаков были обнаружены при изучении органов животных [22], средовой изменчивости урожаев пшеницы, ячменя, сахарной свеклы, картофеля [23], подсолнечника [24; 25], сор- тов персика [26]. Детальным анализом выявлены две причины таких корреляций. Во-первых, по ссылке А. В. Яблокова [22] Я.Я. Рогинский указал на одну причину этого: так как в формуле вычисления CV средняя величина признака является знаменателем, то с ее увеличением должен уменьшаться коэффициент вариации. Вторая причина таких отрицательных корреляций была установлена в результате изучения характера зависимостей величин стандартных (средних квадратичных) отклонений σ от средних величин признаков x̅. Регрессионным анализом этих зависимостей, по данным обширного экспериментального материала, было установлено, что линии таких регрессий проходят выше начала координат [25; 26; 27]. На этом основании был сделан вывод [27], что вследствие такого характера этих регрессий коэффициенты вариации CV должны увеличиваться по мере уменьшения средней величины x̅ признака, т. к. в случае независимости CV от x̅ регрессия σ на x̅ должна описываться прямой линией, проходящей через начало координат. Поэтому по данным опытов с различными культурами величины стандартных отклонений коррелировали с урожаями положительно, а коэффициенты вариации – отрицательно [23; 25; 26].
Необоснованность представления о том, что коэффициент вариации во всех случаях является универсальной мерой степени нестабильности признака, особенно убедительно обнаруживается в ситуациях, когда стандартное отклонение практически неизменно, а варьируют только средние величины признака [25; 26]. Например, Д.И. Шашко [28] установил, что закономерное повышение коэффициентов вариации урожаев подсолнечника, яровой и озимой пшениц, ячменя, кукурузы, гороха и сахарной свеклы по мере усиления засушливости климата природных зон СССР обусловлено последовательным снижением на этом градиенте средних уровней урожайности при остающихся примерно на одном уровне величинах стандартных отклонений.
В тех ситуациях, когда варьируют не только средние величины признака изучаемых объектов, но и абсолютная их нестабильность, корреляцию между CV и x̅ определяет соотношение между степенью различий объектов по x̅ и σ (a i и S i в наших оценках гибридов). Чем меньше различаются изучаемые объекты по вариабельности признака и больше различия по их средним величинам, тем более тесны отрицательные корреляции между значениями CV и x̅, оценки которых по нашим данным достигали r = -0,78 [26] и даже r = -0,968 [25]. При таких корреляциях параметром CV не оценивается вариабельность признаков, а объекты оцениваются и ранжируются по средним величинам изучаемого признака [25; 26]. Если же снижена изменчивость средних величин, как это имело место при испытании гибридов подсолнечника в 2010 г., коэффициент корреляции между CV i и a i практически не отличался от нуля (см. табл. 1, 4). И в этом случае использование коэффициента вариации также ведет к искажению оценок вариабельности признака, а, следовательно, к ошибкам при отборе наиболее стабильных генотипов.
В обзорах, посвященных анализу стабильности, C.S. Lin et al. [14] и И. Пешек с соавторами [15] утверждают, что селекционеры редко используют для оценки генотипов параметры CVi и S2i (как и Si) из-за нежелательных связей этих показателей с оценками урожайности. В определенной степени это справедливо в отношении параметра Si, так как при положительной его корреляции с оценками урожайности ai при отборах по минимальному значению Si могут получать преимущественно низкоурожайные генотипы. По данным опытов 2010 г., когда корреляция между Si и ai достигла r = 0,703** (табл. 4), у гибридов Ригасол ОР и Арена ПР минимальные оценки нестабильности Si сочетались с самыми низкими оценками урожайности, но среди сравнительно стабильных в 2010 г. гиб- ридов оказался достаточно урожайный НК Роки, подтвердивший такую характеристику и в 2011 г. В отличие от параметра Si близкие к нулю и тем более отрицательные корреляции между CVi и ai благоприятствуют отборам на сочетание минимальных коэффициентов вариации с высокой урожайностью, хотя фактически при этом не оценивается стабильность. Это еще раз свидетельствует о неэквивалентности параметров Si и CVi.
Таким образом, оценки нестабильности генотипов могут искажаться при использовании для этой цели не только коэффициентов вариации CVi, но и стандартных отклонений Si. Для повышения точности таких оценок параметром Si в его величины необходимо вносить такие поправки, которые делали бы откорректированные значения Ŝi независимыми от средних величин признака ai. Для устранения влияния одной из независимых переменных на величину отклика зависимой переменной используется регрессионный метод [29]. Возможность такой коррекции основана на свойстве линии регрессии расчленять величины отклонений точек на графике от среднего значения зависимой переменной на две части: обусловленную вариацией независимой переменной и не связанную с ее вариацией [8]. В данном случае отклонение точки, соответствующей величине Si i-го гибрида, от линии регрессии значений Si на оценки ai соответствуют той части величины Si, которая не зависит от оценки ai того же гибрида. Поэтому эта доля от всей Si, которая обозначена Ŝi, отражает собственно нестабильность оценок урожайности изучаемого i-го генотипа. Дж. У. Снедекор [8] обозначает используемые для таких целей линии регрессии как подвижные статистические контроли, позволяющие элиминировать искажающее влияние второй сопутствующей независимой переменной. Вариант этого метода для корреляции величин Si был разработан и использован для повышения точности оценок стабильности урожаев подсолнечника [25], его эффек- тивность была подтверждена анализом данных испытаний сортов персика [26]. Результаты использования этого метода для изучения соотношения представленных в таблице 1 величин параметров Si и аi показаны на графике (рис. 1).
Точки экспериментальных данных на графике (рис. 1) распределились так, что четко выявились два существенно различающихся массива данных. Это соответствует намного большей степени вариабельности оценок урожайности гибридов в условиях экстремальной засухи 2010 г. по сравнению с условиями 2011 г. Использование ранее [25; 26] этого графоаналитического метода во всех случаях позволяло наглядно расчленять совокупности изучаемых объектов на группы с разной степенью нестабильности за счет различий генотипов по восприимчивости к патогенам или скороспелости, а также территорий с разными почвенноклиматическими условиями.
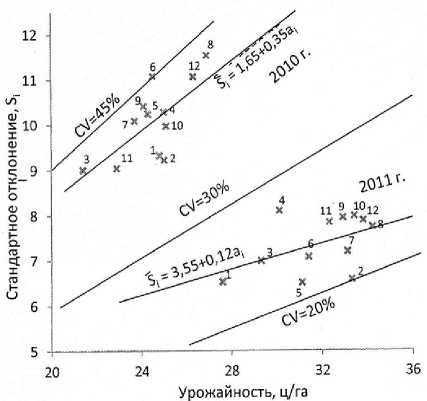
Рисунок 1 – Регрессии средовых стандартных отклонений S i на средние оценки а i урожайности гибридов подсолнечника в 2010 и 2011 гг. в сравнении с линиями равных коэффициентов вариации CV 20; 30 и 45 %
Поскольку координатами такого графика являются значения стандартного отклонения и урожайности, для каждой его точки можно вычислить величину коэф- фициента вариации и построить линии равных величин этих коэффициентов. Для сравнения с регрессиями на график (рис. 1) нанесены три таких линии для CV 20; 30 и 45 %, что соответствует линиям Si = 0,2аi; Si = 0,3аi и Si = 0,45аi. Построение таких линий на этом графике позволило наглядно и удобно реализовать требование Дж. У. Снедекора [8] одновременно сопоставлять величины стандартных отклонений, средних величин признака и коэффициента вариации для избежания заблуждений при изучении нестабильности признаков. В данном случае на графике (рис. 1) четко видно, что разброс точек данных 2011 г. ограничивается линиями равных CV от 20 до 30 %, а точки данных 2010 г. достигают CV = 45 %. Видно также то, что построенные по экспериментальным данным обе линии регрессии Si на аi лежат под углом к линиям равных CV. Этому соответствуют превышающие единицу свободные члены уравнений этих линий регрессии 1,65 и 3,55. А это означает, что линии регрессии проходят выше начала координат, поэтому по мере снижения урожайности должна возрастать в среднем величина коэффициента вариации при равных прочих условиях.
Соответствующая данным опытов 2010 г. совокупность экспериментальных точек на графике (рис. 1) смещена вверх и влево относительно построенной по данным 2011 г. регрессии Si на аi. Ранее с помощью аналогичных графиков тоже было обнаружено, что обусловленное действием разных причин повышение нестабильности признаков различных объектов также приводит к сдвигам соответствующих им точек в том же направлении, при этом их смещение происходит поперек линий равных величин CV [25; 26]. Эту закономерность нельзя выявить, если при изучении изменчивости признаков использовать только коэффициенты их вариации. В то же время учет такого характера сдвигов точек на графике зависимости Si от аi явился основой разработки метода коррекции оценок Si, позволяющего вычислять независимые от величин аi показатели вариабельности признаков Ŝi: соответствующие отдельным объектам точки также ложатся тем выше линии регрессии Si на аi, чем менее стабилен признак этого объекта, и тем ниже этой линии, чем стабильней объект. Результаты вычислений Ŝi по данным оценок аi и Si гибридов в 2010 и 2011 гг. (см. табл. 1) представлены в таблице 5.
Таблица 5
Использование регрессий S i на а i для вычисления прогнозных оценок S , стандартных отклонений и независимых от средних величин а i показателей абсолютной вариабельности Ŝ i урожаев гибридов подсолнечника в сравнении с коэффициентами регрессии на индексы среды b i
|
№ № |
Гибрид |
2010 г. |
2011 г. |
Коэффициент регрессии b i |
|||
|
S , = 1,65 + 0,35a j |
Ŝ i = S i - S , |
S , = 3,55 + 0,12a j |
Ŝ i = S i - S , |
2010 г. |
2011 г. |
||
|
1 |
Савинка |
10,33 |
–1,01 |
6,86 |
–0,34 |
0,90 |
0,81 |
|
2 |
НК Роки |
10,40 |
–1,18 |
7,55 |
–0,98 |
0,92 |
0,88 |
|
3 |
Ригасол ОР |
9,14 |
–0,13 |
7,07 |
–0,09 |
0,89 |
0,93 |
|
4 |
Тристан |
10,40 |
–0,12 |
7,16 |
0,94 |
1,01 |
1,10 |
|
5 |
НК Ферти |
10,16 |
0,08 |
7,28 |
–0,78 |
1,00 |
0,91 |
|
6 |
НК Делфи |
10,22 |
0,85 |
7,32 |
–0,25 |
1,11 |
0,93 |
|
7 |
Тутти |
9,94 |
0,15 |
7,52 |
–0,33 |
1,01 |
0,98 |
|
8 |
НК Конди |
11,06 |
0,47 |
7,65 |
0,09 |
1,15 |
1,08 |
|
9 |
НК Неома |
10,08 |
0,33 |
7,50 |
0,44 |
1,03 |
1,10 |
|
10 |
НК Брио |
10,44 |
–0,47 |
7,56 |
0,42 |
0,99 |
1,08 |
|
11 |
Арена ПР |
9,66 |
–0,62 |
7,43 |
0,41 |
0,91 |
1,09 |
|
12 |
НК Армони |
10,86 |
0,20 |
7,61 |
0,27 |
1,08 |
1,10 |
Независимый от средней величины признака показатель его абсолютной вариабельности Ŝi определяется как разность между общей величиной стандартного отклонения Si и ее зависимой от средней величины признака прогнозной оценки Sj, то есть Si = Si- Sj. Вычисляются прогнозные оценки стандартных отклонений Sj с помощью формул их зависимости от средних величин аi. Как показано на графике (см. рис. 1), значения параметров этих формул различны для данных 2010 и 2011 гг. Сравнение размахов изменчивости значений Si (см. табл. 1) и вычисленных прогнозных оценок Sj (табл. 5) свидетельствует о том, что различия изученных гибридов подсолнечника по величинам параметра Si в значительной степени обусловлены их зависимостью от средних оценок уро- жайности аi: отношения максимальных величин к их минимальным оценкам составили для Si 128 %, для S, 121 % по данным 2010 г., а в 2011 г. для Si 125 %, для S, 115 %. Поэтому независимая от ai доля всей Si, обозначаемая Ŝi, остающаяся после вычитания S, из Si, составляет небольшую величину по сравнению с полным значением средового стандартного отклонения Si оценок урожайности гибрида (табл. 5).
Особенностью откорректированных показателей абсолютной вариабельности Ŝ i урожаев гибридов является то, что часть их отрицательная, а часть – положительная (табл. 5). Отрицательными оценками Ŝ i характеризуются сравнительно более стабильные гибриды, соответствующие которым точки на графике (см. рис. 1) располагаются ниже линии регрессии S i на а i . У гибридов, точки которых на том же графике расположены выше линии регрессии, а оценки Ŝ i положительны (табл. 5), повышена средовая вариабельность урожаев, причем степень их нестабильности тем выше, чем более высокую оценку Ŝ i получил гибрид.
При сопоставлении параметров а i и S i изученных гибридов обнаруживаются тенденции повышения как урожайности, так и оценок нестабильности проявления этого признака с увеличением числа дней от даты сева до цветения (см. табл. 1). Вследствие этих ковариаций частично совпадают ранги нестабильности гибридов при их оценках параметрами S i и Ŝ i . Однако в случаях нарушения указанных тенденций более точными оказываются оценки параметром Ŝ i . Например, среди рано цветущих гибридов самой низкой в 2010 г. оказалась оценка S i гибрида Рига-сол ОР за счет самой низкой его урожайности, а при оценке по Ŝ i он занял 5-е место. Гибрид же НК Роки занял 3-е место при оценке по S i за счет повышенной урожайности, но первое место при оценке по Ŝ i , что подтвердилось у него и в 2011 г.
Для сопоставления с вычисленными величинами Ŝ i в таблице 5 показаны оценки коэффициентов регрессий b i урожаев гибридов на индексы среды. При сравнении величин этих параметров обнаруживается тесная положительная корреляция между ними. Более того, практически во всех случаях отрицательным величинам параметра Ŝ i соответствуют величины коэффициентов регрессий b i , равные единице или меньше ее. Это означает, что на графике (см. рис. 1) точкам, расположенным ниже линий регрессий S i на а i , соответствуют координаты гибридов подсолнечника, характеризующихся также еще и низкими величинами параметра b i . Расположенные же выше таких линий регрессий точки отражают свойства тех гибридов, у которых не только повышены оценки независимых от а i показателей вариабельности урожаев Ŝ i , но и увеличены значения коэффициентов регрессий b i оценок урожайности на индексы среды. Аналогичное распределение точек генотипов с разными оценками параметра b i относительно линий регрессий S i на а i было обнаружено и при изучении сортов персика разных сроков созревания [26]. Эти факты являются важным свидетельством, подтверждающим обоснованность вычисления и использования независимых от средних величин признаков оценок их абсолютной вариабельности Ŝ i .
Как было показано в таблице 4, очень высокими оказались также коэффициенты корреляции между величинами параметров Si и bi, что подтверждается и результатами исследований с генотипами других видов возделываемых растений [12; 16; 30; 31]. При построении графика зависимости между этими параметрами (рис. 2) по данным испытаний гибридов подсолнечника (см. табл. 1 и 5) обнаруживается несовпадение линий регрессий для оценок параметров, полученных по данным испытаний в условиях экстре- мально засушливого 2010 г. и обычного 2011 г. На этом графике (рис. 2) оценки параметра Si и линия регрессии в целом сместились по данным 2010 г. вправо относительно показателей 2011 г., что соответствует повышенной экологической вариабельности урожаев в условиях засухи.
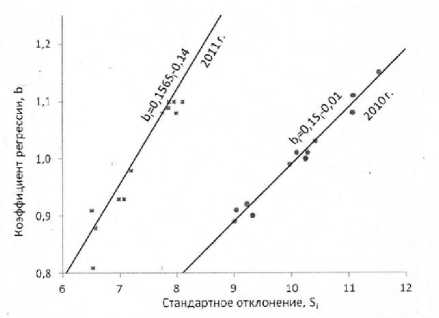
Рисунок 2 – Регрессии значений параметра b i гибридов подсолнечника на средовые стандартные отклонения оценок их урожайности S i по данным испытаний на европейской территории России в 2010 г. и в 2011 г.
Поскольку корреляции между оценками генотипов параметрами S i и b i очень высоки, возникают вопросы, во-первых, что является причиной таких корреляций, во-вторых, содержат ли параметры S i и b i одинаковую или разную информацию об оцениваемых ими генотипах. С целью выяснения причин высоких корреляций между значениями этих параметров H.C. Becker [30] проанализировал формулу вычисления дисперсии S2 i оценок урожайности i-го генотипа:
S j 2 = S2 + 2(b i - 1)S £ + (b i - 1)2S2 + S ^ , (1)
где S 2 = Е ) (х .] -х )2 , т. е. это дисперсия индексов среды; S2d i – остаточная дисперсия.
Вычислением величин четырех дисперсий этой формулы по данным экологических сортоиспытаний кукурузы, ячменя и овса H.C. Becker [30] показал, что дисперсия S2p в каждом из испытаний одинакова для всех генотипов, что следует и из описания метода Эберхартом и Расселлом [5]; поэтому различия дисперсий S2i в основном определяются величиной произведения 2(Ьг — 1)^2, а следовательно, оценкой bi i-го генотипа. Чем больше отношение произведения 2(bz —1)^2 к значению дисперсии Wi, тем выше корреляция между оценками S2i и bi с учетом формулы (5) для Wi.
Для того чтобы не только выяснить причину высокой корреляции между оценками генотипов параметрами S i и b i , но и понять, в какой мере эквивалентна отражаемая ими информация, нами была проанализирована другая формула [16]. Известно, что при использовании метода приведенной главной оси по К. Пирсону уравнение прямой регрессии у на х имеет вид:
у = ^ • % + к, (2)
а по методу наименьших квадратов (МНК):
у = г — • % + к. (3)
^ %
Соответственно коэффициент регрессии для формулы (2) вычисляется как соотношение стандартных отклонений зависимой переменной σy и независимой σx, а для формулы (3) – как то же соотношение, умноженное на коэффициент корреляции между y и x. При вычислениях значений параметра bi по Эберхарту и Расселлу стандартному отклонению σy соответствует параметр Si, а знаменателю σx – стандартное отклонение индексов среды Sp, поэтому ь< = г^. (4) ^Р
Поскольку величина Sp одинакова для всех генотипов одного экологического сортоиспытания, то при приближающихся к 1 или одинаковых корреляциях между Si и Sp оценки генотипов параметрами Si и bi должны не только тесно коррелировать, но и содержать в основном экви- валентную информацию, которая определяется показателем вариабельности урожаев Si.
Этот вывод противоречит общепризнанным классификациям параметров стабильности и адаптивности (14, 15, 32), в соответствии с которыми параметр S2 i (как и S i ) относится к статическому (биологическому) типу, а b i – к динамическому (агрономическому) типу характеристик экологической изменчивости оценок урожайности генотипов. Однако авторы этих классификаций [14; 15] признают, что параметр b i может быть отнесен к статическому типу оценок, если считать оптимальной его величину не b i = 1,00, а b i = 0. Однако, судя по характеру распределения значений S i и b i на графике (рис. 2), с таким же основанием можно считать оптимальным генотип не с минимальной, а со средней величиной параметра S i .
В связи с этим важно отметить, что общий уровень величин S i характеризует не степень нестабильности испытуемых генотипов, а разнообразие условий проведения опытов. В частности, существенно возросшие значения S i в 2010 г. были обусловлены увеличенным диапазоном условий влагообеспеченности посевов подсолнечника от благоприятных на юге европейской части России до максимально стрессовых в Поволжье. Разнообразие же генотипов по устойчивости к стрессам оценивается только различиями их оценок величинами S i , а точнее – значениями параметра Ŝ i , вычисляемых как отклонения от регрессии S i на а i , что соответствует отклонениям величин b i от значения b i =1,00, о чем свидетельствуют данные таблицы 5.
Хотя параметры Si и bi содержат эквивалентную информацию, каждый из них имеет свои преимущества при их использовании для оценок генотипов. Оценки Si проще вычисляются и ясней интерпретируются. Важно также, что для их вычислений не требуется определять индексы среды, поэтому нет ограничения на число генотипов в экологическом сортоиспыта- нии, которое может быть снижено до двух сортов или гибридов.
В отличие от стандартных отклонений S i коэффициент регрессии на индексы среды b i не содержит информацию о степени разнообразия условий проведения опытов экологического сортоиспытания, так как в соответствии с уравнением (4) величина b i определяется соотношением стандартных отклонений S i и S p . Это ясно видно на графике (рис. 2): соответствующие 12 гибридам координаты точек на нем по данным испытаний 2010 г. по сравнению с оценками 2011 г. сдвинуты только вправо вдоль оси абсцисс, тогда как диапазон вариации ординат точек, соответствующих величинам параметра b i , практически одинаков по данным испытаний этих двух лет. Следовательно, параметр b i оценивает только различия генотипов по их реакциям на изменения условий, а степень разнообразия условий проведения опытов сортоиспытания характеризует стандартное отклонение S p вариации индексов среды.
Безусловно, важным преимуществом параметра b i по сравнению с S i является то, что в сочетании с данными о средних оценках урожайности генотипа а i и величинами индексов среды можно вычислять математические ожидания урожайности i-го генотипа при максимальном индексе среды для оценки потенциала его урожайности и при минимальном индексе – для оценки устойчивости к неблагоприятным условиям [12]. В таблице 6 приведены такие оценки изученных гибридов подсолнечника, вычисленные по данным таблиц 1 и 3.
Хотя ожидаемые оценки урожайности (табл. 6) вычислялись с использованием значений bi, причины различий генотипов по величинам этого параметра или отсутствия различий становятся понятными лишь при сравнении математических ожиданий урожаев при минимальном и максимальном индексах среды. Например, в 2010 г. гибриды Савинка и Ригасол ОР имели близкие значения bi = 0,90 и bi = 0,89, но по урожайности при индексах среды 11 и 40 ц/га Савинка получил ранги 1 и 10 вследствие наибольшей устойчивости к экстремальной засухе, но ограниченного потенциала урожайности в относительно благоприятных условиях, у гибрида же Ригасол ОР оба эти свойства проявились в минимальной степени. В этом же году у лучших гибридов НК Конди и НК Брио значения bi оказались разными: bi = 1,15 и bi = 0,99. Высокая величина bi у НК Конди обусловлена тем, что его устойчивость к засухе оказалась близкой к средней (11,4 ц/га при индексе 11 ц/га), но по отзывчивости на благоприятные условия он в 2010 г. занял 1-е место, так как математическое ожидание его урожая превысило индекс среды на 4,7 ц/га. Более низкое значение bi у НК Брио связано с тем, что по устойчивости к засухе он уступил только Савинке и НК Роки, но его отзывчивость на комфортные условия 2010 г. лишь на 0,5 ц/га превысила среднюю.
Таблица 6
Вычисленные по уравнениям регрессии оценок урожайности на индексы среды математические ожидания урожаев (ц/га) гибридов подсолнечника при минимальных и максимальных индексах для условий, сложившихся на европейской территории России в 2010 и 2011 гг., и степень реализаций потенциалов гибридов в условиях стрессов этих лет
|
№№ |
Гибрид |
2010 г. |
2011 г. |
||||
|
индекс среды, ц/га |
отношение а:б |
индекс среды, ц/га |
отношение а:б |
||||
|
а 11 |
б 40 |
а 16 |
б 45 |
||||
|
1 |
Савинка |
12,7 |
38,7 |
0,328 |
14,7 |
38,1 |
0,386 |
|
2 |
НК Роки |
12,6 |
39,2 |
0,321 |
19,3 |
44,8 |
0,431 |
|
3 |
Ригасол ОР |
9,4 |
35,2 |
0,267 |
14,5 |
41,6 |
0,348 |
|
4 |
Тристан |
11,3 |
40,6 |
0,278 |
12,6 |
44,6 |
0,282 |
|
5 |
НК Ферти |
10,7 |
39,8 |
0,269 |
16,6 |
43,1 |
0,385 |
|
6 |
НК Делфи |
9,5 |
41,8 |
0,227 |
16,7 |
43,6 |
0,383 |
|
7 |
Тутти |
10,0 |
39,4 |
0,254 |
17,4 |
46,0 |
0,378 |
|
8 |
НК Конди |
11,4 |
44,7 |
0,255 |
16,9 |
48,4 |
0,349 |
|
9 |
НК Неома |
10,1 |
40,1 |
0,252 |
15,3 |
47,3 |
0,323 |
|
10 |
НК Брио |
11,7 |
40,5 |
0,289 |
16,3 |
47,5 |
0,343 |
|
11 |
Арена ПР |
10,7 |
36,9 |
0,290 |
14,9 |
46,6 |
0,320 |
|
12 |
НК Армони |
11,7 |
43,1 |
0,271 |
16,3 |
48,2 |
0,338 |
Математические ожидания оценок урожайности при минимальных и максимальных индексах среды надежнее характеризуют уровни устойчивости генотипов к стрессам и потенциалы их продуктивности в благоприятных условиях потому, что они вычисляются с использованием данных всех опытов экологического испытания с учетом оцениваемых параметром bi тенденций снижения урожаев по мере роста проявлений стрессовых факторов и повышения урожаев на градиенте улучшения условий внешней среды. В частности, в условиях экстремальной засухи 2010 г. самыми высокими оказались оценки математически ожидаемых урожаев гибридов Савинка и НК Роки при индексе среды 11 ц/га (табл. 6), что подтверждает сделанный по данным таблиц 1 и 2 вывод о наибольшей устойчивости к таким условиям рано зацветающих генотипов подсолнечника. Минимальной в этих условиях оказалась ожидаемая оценка урожая Ригасол ОР, но причиной этого может быть общий низкий потенциал урожайности этого гибрида, проявляющийся оба года как в комфортных условиях, так и при стрессах. О возможном влиянии потенциалов урожайности генотипов на оценки их ожидаемых урожаев при стрессах свидетельствует и то, что у самых урожайных в благоприятных условиях гибридов НК Конди, НК Брио и НК Армони в оба года вычисленные оценки урожаев при минимальных индексах среды превысили эти индексы.
Математические ожидания урожаев лишь у одного гибрида Ригасол ОР оказались ниже, а у трех – НК Конди, НК Брио и НК Армони – выше как минимальных, так и максимальных индексов среды в оба года. У остальных же генотипов проявились различия по реакциям на условия 2010 и 2011 гг. (табл. 6). Этому соответствуют и различия по оценкам параметров взаимодействия генотип – среда W i и S2d i , которые оказались в среднем повышенными по данным испытаний 2011 г. (см. табл. 3). Проявляя наибольшую устойчивость к стрессовым факторам 2010 г., Савинка и НК Роки качественно различно реагировали на неблагоприятные условия 2011 г., на фоне которых при индексе среды 16 ц/га математически ожидаемая урожайность
Савинки оказалась ниже этого уровня на 1,3 ц/га, а НК Роки – выше его на 3,3 ц/га, за счет чего этот скороспелый гибрид превзошел по урожайности в этих условиях все остальные гибриды (табл. 6). Гибрид Тристан проявил несколько повышенную устойчивость к стрессам экстремально засушливого года по оценкам ожидаемых урожаев при индексах как 11, так и 40 ц/га, но к стрессовым факторам 2011 г. оказался самым неустойчивым при ожидаемой урожайности ниже минимального индекса среды на 3,4 ц/га. Гибрид Тутти, напротив, оказался неустойчивым к условиям экстремальной засухи 2010 г. при индексах среды как 11, так и 40 ц/га, но на фоне стрессовых факторов 2011 г. при индексе среды 16 ц/га занял второе место после НК Роки и 6-е место по ожидаемому потенциалу урожайности при индексе 45 ц/га.
Специфичный тип взаимодействия генотип – среда проявили гибриды НК Делфи и НК Ферти (табл. 6). В условиях 2010 г. при индексе среды 11 ц/га обнаружилась низкая их устойчивость к экстремальной засухе, особенно у НК Делфи, но при индексе 40 ц/га ожидаемая урожайность НК Ферти оказалась близкой к средней, а у НК Делфи на 1,8 ц/га выше ее. Однако оба эти гибрида проявили повышенную устойчивость к стрессам 2011 г., но в этот год оценки их потенциалов урожайности при индексе 45 ц/га оказались сниженными на 1,9 и 1,3 ц/га по сравнению со средним уровнем. У гибрида НК Неома выявлена сниженная устойчивость к стрессам как 2010, так и 2011 г., а оценка потенциала урожайности оказалась средней в 2010 г. и повышенной в 2011 г.
По сравнению с показателями 2011 г. в условиях засухи 2010 г. снизились математически ожидаемые урожаи не только в условиях наибольших стрессов при минимальном индексе среды, но у большинства гибридов, кроме Савинки, в меньшей степени реализовался потенциал урожайности при максимальном индексе среды 40 ц/га, чем при 45 ц/га 2011 г. (табл. 6).
При этом доля реализации этого потенциала, соответствующего индексу 40 ц/га, составила при индексе 11 ц/га от 0,252 до 0,328, то есть от 25,2 до 32,8 %, тогда как в 2011 г. урожайность при индексе 16 ц/га по сравнению с ее оценкой при индексе 45 ц/га составила от 28,2 до 43,1 %. В оба года при минимальных индексах среды наибольшей была доля реализации потенциалов урожайности у рано зацветающих гибридов Савинка и, особенно, НК Роки. Уникальной особенностью гибрида НК Роки является то, что при потенциале урожайности немного не достигающем уровней максимальных индексов среды, средние оценки его урожайности увеличены по всем зонам России за счет повышенной засухоустойчивости и, особенно, устойчивости к таким стрессам, которые проявлялись в обычном по влагообеспеченности 2011 г., поэтому в оба года была высокой доля реализации потенциала его урожайности на фонах индексов среды как 11 ц/га, так и 16 ц/га. Противоположные показатели адаптивности проявил гибрид Тристан: его урожаи, превысившие в 2010 г. индексы 11 и 40 ц/га, свидетельствуют о повышенной засухоустойчивости, но, судя по аналогичным показателям 2011 г., он проявил явно низкую устойчивость к преобладающим в этом году стрессам на европейской территории России. В результате этого у Тристана не повысилась доля реализации потенциала урожайности при индексе 16 ц/га в отличие от остальных гибридов.
Гибриды подсолнечника, склонные к наибольшим проявлениям взаимодействия генотип – год, выявляются не только по изменениям в 2011 г. по сравнению с 2010 г. математических ожиданий их урожаев при минимальных и максимальных индексах среды (табл. 6). При сравнении данных таблиц 6 и 5 обнаруживается, что у проявивших наибольшую степень такого взаимодействия гибридов в эти годы качественно различна и степень стабильности урожаев, оцениваемая параметром Ŝi. Этот параметр меняет в 2011 г. знак с минуса на плюс у гибридов Тристан, НК Брио и Арена ПР и с плюса на минус у НК Ферти, НК Делфи и Тутти. Это означает, что у Тристана, НК Брио и Арена ПР в 2011 г. возросла экологическая вариабельность, а у НК Ферти, НК Делфи и Тутти она снизилась.
Смены рангов генотипов при их испытаниях в разные годы и в разных пунктах затрудняет выбор лучших гибридов по данным экологических испытаний. Поэтому важно изучить взаимосвязи между параметрами взаимодействия генотип – среда, выявить причины их вариации. Оценки коэффициентов корреляций между такими параметрами представлены в таблице 7.
Таблица 7
Коэффициенты корреляций между величинами параметров взаимодействия генотип – среда по данным испытаний гибридов подсолнечника на европейской территории России в экстремально засушливом 2010 г. (над диагональю) и в обычном по количеству осадков 2011 г. (под диагональю)
|
Параметры |
W i |
b i |
S2d i |
Cd i , % |
а i , ц/га |
|
W i |
1 |
0,142 |
0,963*** |
–0,898*** |
0,454 |
|
b i |
–0,464 |
1 |
0,085 |
0,228 |
0,659* |
|
S2d i |
0,991*** |
–0,425 |
1 |
–0,939*** |
0,416 |
|
Cd i , % |
–0,949*** |
0,708** |
–0,930*** |
1 |
–0,189 |
|
а i , ц/га |
–0,644* |
0,588* |
–0,613* |
0,695* |
1 |
Коэффициенты корреляции переходят уровни существенности: * – 5 %-ный; ** – 1 %-ный;
*** – 0,1 %-ный где ^j dZj— сумма квадратов отклонений оценок i-го генотипа от регрессии величин его урожаев на индексы среды.
Поскольку S i — -^аі 1 П—2 ' *
из формулы (5) следует, что значения эковаленты W i тем больше, чем значительней отклонение b i от b i = 1,00 и больше средний квадрат отклонений x ij от линии регрессии S2d i . Следовательно, в изменчивость параметра W i включены проявления взаимодействия генотип – среда как прогнозируемые по оценкам b i , так и непредсказуемые, измеряемые параметром S2d i . Представленные в таблице 7 коэффициенты корреляции величин W i и S2d i свидетельствуют о том, что изменчивость суммарных оценок этих компонентов параметром W i определялась по данным испытаний гибридов непредсказуемой вариацией величин S2d i на 92,7 % в 2010 г. и на 98,2 % в 2011 г. Поскольку эти оценки были обусловлены как в 2010 г., так и в 2011 г. специфическими реакциями изученных гибридов на разнообразие почвенно-климатических условий разных зон европейской части России, эффекты взаимодействий генотип – местность оказались в основном непрогнозируемыми, особенно в 2011 г.
Из формул (5) и (6) выводится уравнение зависимости величин параметра S2di от значений эковаленты Wi, которое ет вид:
име-
Представленные в таблице 7 данные свидетельствуют о том, что наиболее тесны и высоко существенны корреляции между параметрами W i , S2d i и Cd i . Причины таких высоких коэффициентов корреляций становятся понятными при рассмотрении формул связи между величинами этих параметров.
В 1967 г. F.W. Schnell (цитируется по 32) предложил уравнение связи величин W i с параметрами регрессий оценок урожайности генотипов на индексы среды:
C Z _ _
Sd. — a' n
—
- [W j —(b j —1)22(x .j —x .. )2l.
I j J
Уравнение (7) свидетельствует о
том,
W , — (Ь , —l/^x .j —x .. )2+^d 2j ,
что остаточная дисперсия S2d i является полностью не прогнозируемым компонентом эковаленты в отличие от второго ее компонента, параметр b i которого в сочетании с индексами среды (x .j — x) позволяет вычислять ожидаемые оценки урожайности. Поэтому K.M. Eskridge [33], как и S.A. Eberhart и W.A. Russell [5], считает, что параметр S2d i является идеальной мерой непредсказуемых нарушений реакций i-го генотипа на условия
среды. Однако авторы классификаций параметров стабильности и адаптивности считают неприемлемым отнесение остаточной дисперсии S2d i к числу таких параметров [14; 15]. В частности, И. Пешек с соавторами [15] пишут: «…средний квадрат отклонений от регрессии не может обладать свойствами параметра устойчивости, применительного к предсказанию или обобщению». В то же время они относят эковаленту к числу параметров устойчивости, хотя по G. Wricke [9] отклонения эковалент от W i = 0 обусловлены также непредсказуемыми реакциями генотипа на условия среды. А насколько эффективно для получения важных выводов и обобщений может быть использование параметра S2d i можно показать на двух примерах. Анализом изменчивости оценок урожайности в опытах агроэкологического испытания сортов и гибридов подсолнечника было установлено, что величины параметра S2d i сортов-популяций варьировали от 0,006 до 0,009, трехлинейных гибридов – от 0,015 до 0,042, простых гибридов – от 0,031 до 0,073 [34]. Изучая влияние разного числа (от 1 до 14) генотипов в сор-тосмесях сои на их урожайность и показатели стабильности, A.K. Walker и W.R. Fehr [35] обнаружили, что число компонентов смесей не влияло на оценки их урожайности и величины коэффициентов регрессии на индексы среды b i , но средний квадрат отклонений от регрессий S2d i закономерно снижался от 17,52 при одном генотипе в посеве до 6,54 при девяти сортах в смесях, оставаясь на этом уровне при дальнейшем увеличении числа компонентов. Эти и другие аналогичные примеры позволяют полагать, что снижение проявлений взаимодействия генотип – среда по мере увеличения генетической гетерогенности популяций, которое R.W. Allard и A.D. Bradshaw [4] назвали «буферным эффектом», обусловлено в значительной мере уменьшением средних квадратов отклонений от регрессии S2d i .
Возросшее в 2011 г. у большинства изученных гибридов проявление взаимодействия генотип – среда, выявленное по увеличению эковалент W i (см. табл. 3) и связанное со спецификой реакций генотипов на стрессовые факторы в условиях лучшей обеспеченности влагой по сравнению с 2010 г. (см. табл. 6), почти полностью было обусловлено непредсказуемой изменчивостью урожаев, оцениваемой параметром S2d i (табл. 7). Это также свидетельствует о том, что этот показатель является важным компонентом характеристик особенностей адаптивных реакций генотипов. Однако следует учитывать, что величина S2d i является лишь мерой степени отклонения отклика i-го генотипа от усредненной реакции на те же условия всех генотипов данного испытания независимо от того, положительна или отрицательна специфичная реакция этого генотипа. Например, при анализе данных опытов экологического испытания сортов сои одинаково высокими оказались оценки S2d i как самого засухоустойчивого, так и самого чувствительного к засухе сортов. G. Wricke [9] также показал, что и эковалента равна нулю W i = 0, если во всех опытах экологического испытания урожаи i-го сорта отклоняются на одинаковую величину от средних урожаев всех сортов, а высокие оценки W i получают сорта как с увеличенной реакцией на изменения условий, так и совсем не реагирующие на их изменения.
Различия между специфическими адаптивными реакциями гибридов подсолнечника на условия 2010 и 2011 гг. особенно четко проявились по сдвигам оценок коэффициента детерминации: только у НК Ферти и НК Армони значение параметра Cd i в эти годы было одинаковым, а у остальных гибридов его величина в 2011 г. заметно снизилась (см. табл. 3). Поэтому важно понять, особенности каких адаптивных реакций отражает этот параметр и с чем связаны изменения его величины.
Вычисление коэффициента детерминации основано на определении возведенного в квадрат коэффициента корреляции между оценками урожайности i-го генотипа и индексами среды опытов экологического испытания. Это позволяет определить долю изменчивости урожаев i-го генотипа, обусловленную изменчивостью индексов среды, а по разности – и долю вклада в вариацию оценок урожайности специфических реакций генотипа на условия среды, измеряемые этими индексами. Если в качестве коэффициента детерминации используется просто величина r2 i , эти вклады в общую изменчивость урожаев оцениваются в долях от единицы, если же коэффициент детерминации вычисляется как Cd, % = 100 r2, эти оценки выражаются в процентах. В обоих случаях оценки доли зависимой от индексов среды вариации признака i-го генотипа являются характеристикой степени симилярности его реакций на изменения условий, так как индексами среды измеряется усредненная реакция всех генотипов испытания на эти условия.
C.S. Lin с соавторами [14], как и И. Пешек и др. [15] не включили коэффициент детерминации в число параметров стабильности и адаптивности, рассматривая лишь высокое значение r2 как одно из условий возможности использования параметра bi. Однако в публикациях ряда авторов показано, что использование коэффициента детерминации позволяет получать важную информацию об адаптивности изучаемых генотипов. Например, A.R. Pedersen et al. [36] считают, что для описания и предсказания отклика сорта в серии экспериментов следует одновременно использовать три параметра: bi, S2di и r2i. Анализом данных испытаний сортов озимой пшеницы они установили, что при сниженных оценках r2i сортов менее надежны предсказания их урожайности, а разность 1–r2i является индикатором степени отклика сорта на специфические условия среды. Ими был выявлен также оптимальный диапазон оценок r2i: «Испытываемые генотипы обычно давали r2i между 64 и 85 %, а сорта, рекомендованные для выращивания в Мичигане, имели r2i от 73 до 82 %».
По данным наших испытаний гибридов подсолнечника величины коэффициентов детерминации Cd i в оба года тесно отрицательно коррелировали со значениями как эковалент W i , так и остаточных дисперсий S2d i (табл. 7). H.C. Becker и J. Leon [31] считают даже, что параметр r2 i может служить равноценной заменой оценок S2d i , так как
?
Гі2 = 1-4. (8) Зі
Поскольку по сравнению с 2010 г. в 2011 г. возросли средние по всем пунктам оценки урожайности a i всех гибридов (см. табл. 1) и у большинства гибридов увеличились значения S2d i (см. табл. 3), казалось правильным предположение, что это повышение урожайности в 2011 г. связано с проявлением специфических реакций генотипов, позволяющих лучше использовать благоприятные условия среды. Однако по результатам испытаний 2011 г. корреляции оценок a i с S2d i и с W i оказались отрицательными, существенными, хотя по данным 2010 г. они были положительными (табл. 7). Соответственно, корреляция a i с Cd i по данным 2011 г. стала существенной, положительной. Это означает, что причиной возросших в 2011 г. величин S2d i и W i , а также снижения коэффициентов детерминации оценок урожайности индексами среды Cd i является проявление специфических адаптивных реакций гибридов на характерные для этого года стрессовые факторы, хотя в целом более благоприятные условия привели к повышению средних оценок урожайности a i .
Поскольку в формулы вычисления величин Wi (5) и S2di (7) включены значения bi, можно было бы ожидать наличие корреляций между вариацией этих параметров. Однако в оба года коэффициенты корреляции как между Wi и bi, так и меж- ду S2di и bi оказались незначимыми (табл. 7). Причинами этого может быть как преобладание вариации квадратов отклонений от регрессий Σd2ij по сравнению с изменчивостью bi, так и то, что в формулы (5) и (7)] включены значения не bi, а разности (bi – 1).
Хотя на основании формулы (8) H.C. Becker и J. Leon [32] не связывают вариацию величин r2 i , а значит и Cd i , с изменчивостью b i , а считают ее обусловленной вариацией S2d i , по данным 2011 г. обнаружена высокая, положительная, существенная корреляция между величинами Cd i и b i , а в 2010 г. она была также положительной, хотя и не существенной (табл. 7). Следует учесть, что формула (8) основана на отношении S2d i к S i , а по данным статьи [16], таблицы 4 и графика (см. рис. 2), параметр S i очень тесно связан с коэффициентом регрессии b i , поэтому отношение S2d i к S i эквивалентно отношению S2d i к b i . Наличие этой связи становится очевидным при рассмотрении формулы вычисления коэффициента детерминации, которую предложил M.J. Pinthus [37]:
,2 ti^Xj-xJ2
S2di = 2,74 легла значительно выше обоих регрессий, а точки остальных гибридов с диапазоном величин S2d i от 4,03 до 7,57 расположились вдоль линии Cd i = 47,9 + 41,4b i (коэффициент корреляции b i с Cd i r = 0,957 переходит 5 %-ный уровень существенности). Следовательно, зависимость коэффициента детерминации Cd i от значений b i выявляется более четко, если производить стратификацию данных по уровням остаточных дисперсий S2d i . В результате такой же стратификации данных испытаний гибридов в 2010 г. на аналогичном графике (рис. 4) также выявляется зависимость величин параметра Cd i от коэффициентов регрессии b i .
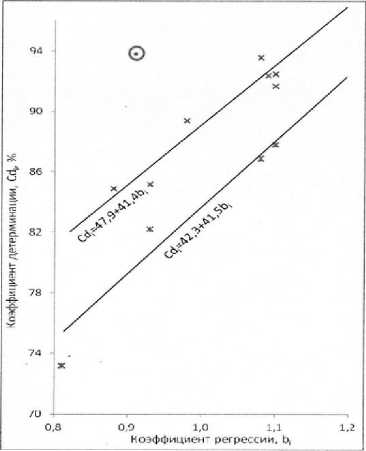
Рисунок 3 – Зависимости коэффициентов детерминации оценок урожайности индексами среды Cd i (%) от коэффициентов регрессии оценок урожайности на индексы среды bi с учетом диапазонов остаточных дисперсий S2d i ( • - 2,74; x — от 4,03 до 7,57; * — от 8,38 до 11,98) по данным испытаний гибридов подсолнечника на европейской территории России в 2011 г.
Вычисленный по результатам испытаний всех 12 гибридов в 2010 г. коэффициент корреляции между b i и Cd i оказался хотя и положительным, но небольшим и не значимым даже на 10 %-ном уровне (см. табл. 7). Однако соответствующие этим гибридам точки на графике (рис. 4)
распределились также таким образом, что величины остаточных дисперсий S2d i этих генотипов закономерно возрастают в направлении от верхней левой к нижней правой части графика: от S2d i = 2,00 у Арены ПР до S2d i = 9,54 у НК Армони. При этом точки гибридов НК Делфи, Тутти, НК Брио, НК Роки и Ригасол ОР легли вдоль линии Cd i = 85 + 12b i с коэффициентом корреляции r = 0,910 (переходит 5 %-ный уровень существенности), а точки гибридов НК Конди, НК Неома, НК Ферти, Тристан и Савинка расположились вдоль линии Cd i = 71 + 21,7b i с коэффициентом корреляции r = 0,921 (переходит 5 %-ный уровень существенности).
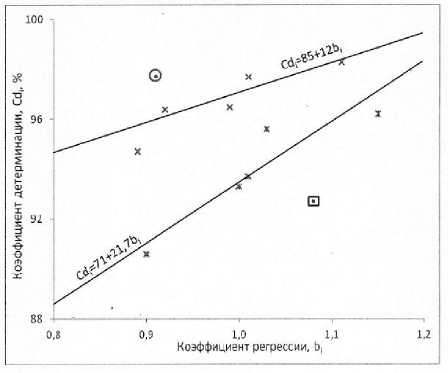
Рисунок 4 – Зависимости коэффициентов детерминации оценок урожайности индексами среды Cd i (%) от коэффициентов регрессии оценок урожайности на индексы среды b i с учетом диапазонов S2d i ( • – 2,00; х — от 2,22 до 4,57; * - от 5,05 до 8,74; • – 9,54) по данным испытаний гибридов подсолнечника на европейской территории России в 2010 г.
Представленные на графиках (рис. 3 и 4) результаты регрессионного анализа соотношения параметров bi и Cdi с учетом S2di свидетельствуют о том, что симиляр-ность откликов гибридов на разнообразие условий проведения опытов испытаний снижается с увеличением остаточных дисперсий и с уменьшением коэффициентов регрессий на индексы среды, что со- ответствует формуле (9). Из этой же формулы следует, что на величину Cdi, а значит и на степень симилярности откликов генотипов положительно влияет также диапазон изменчивости индексов среды (x.j — х..) опытов экологического испытания. Величины произведения b2^j(x.j-х..)2 i-го генотипа соответствуют значению средовой дисперсии S2i оценок его урожайности, а как свидетельствует формула (8), отношение S2di к S2i определяет величину r2i и степень симилярности реакций генотипов на изменения внешних условий.
Проведенный корреляционный (см. табл. 4 и 7) и регрессионный (см. табл. 6 и рис. 2, 3 и 4) анализы позволяют объяснить причины выявленных различий по влиянию условий проведения испытаний в 2010 и 2011 гг. на оценки параметров Cdi и S2di гибридов подсолнечника. На европейской территории России в экстремально засушливом 2010 г. условия проведения опытов максимально различались по степени обеспеченности посевов влагой, поэтому были увеличены различия индексов среды (x.j - х..) и соответственно b2 Zj(x.j - х .)2, что обусловило высокие значения стандартных отклонений Si средовой вариабельности оценок урожайности гибридов и положительно повлияло на оценки симилярности их откликов Cdi. Поскольку главным стрессовым фактором в 2010 г. являлся дефицит влаги, специфичность реакций генотипов на который обычно невысока, сниженными оказались оценки параметра S2di и, как следствие, Wi. Это также положительно сказалось на величины Cdi. В опытах 2011 г. в большой степени проявились эффекты взаимодействия генотип – среда, что может быть связано с различиями гибридов по реакции на поражение растений патогенами, на ненастную погоду при уборке урожая и на другие стрессы. Это проявилось в увеличении значений S2di и Wi, которое оказалось отрицательно связанным с уровнями оценок урожайности. Уменьшение диапазона ин- дексов среды привело к снижению средовой вариации Si оценок урожайности, что в сочетании с повышением остаточных дисперсий S2di обусловило сниженные значения коэффициентов детерминации Cdi.
Величину параметра b i авторы ряда публикаций считают мерой отзывчивости i-го генотипа на благоприятные внешние условия, но высокое значение b i может быть и следствием значительного снижения оценок его урожайности на фоне стрессов [16]. Коэффициенты корреляции между величинами b i и средними оценками урожайности a i в 2010 и 2011 гг. оказались положительными, средней величины, соответствующими 43,4 и 34,6 % сопряженной изменчивости этих параметров (см. табл. 7). Теснота этой корреляции определяется характером взаимного расположения линий регрессии оценок урожайности изученных генотипов на индексы среды: при схождении этих линий в одной точке корреляция между b i и a i наиболее тесна, а по мере расширения области сходимости и особенно при ее отсутствии, величины коэффициентов такой корреляции стремятся к нулю [38]. Поэтому величины этих корреляций непосредственно не связаны с особенностями метеорологических условий 2010 и 2011 гг.
Выводы. 1. В условиях экстремальной засухи 2010 г. на европейской территории России снизилась по сравнению с 2011 г. средняя величина и возросла вариабельность оценок урожайности изученных гибридов подсолнечника при высокой (от 90,6 до 98,3 %) симилярности адаптивных реакции вследствие меньшей их специфичности в основном за счет снижения остаточных дисперсий и частично вследствие изменения наклонов регрессии на индексы среды по сравнению с 2011 г.
-
2. В более благоприятных условиях 2011 г. возросли как оценки урожайности гибридов, так и специфичность их адаптивных реакций, поэтому симилярность этих реакций оказалась снижена (70,2–93,8 %).
-
3. Возросшие в 2011 г. оценки W i взаимодействия генотип – местность, в основном за счет увеличения остаточных дисперсий S2 di , были обусловлены специфическими реакциями гибридов не на благоприятные условия, а на стрессы, связанные с повышенной увлажненностью территорий.
-
4. Вычисления математических ожиданий урожаев i-го генотипа при минимальном и максимальном индексах среды позволяют не только оценить его устойчивость к стрессам и потенциал урожайности, но и выявить качественные различия по проявлению этих свойств в разные по погодным условиям годы.
-
5. У зацветающих на 3–5 дней раньше других гибридов Савинка и НК Рокки в оба года снижены оценки потенциалов урожайности при максимальных индексах среды, но при низких индексах среды в экстремально засушливом году их урожайность была самой высокой. К стрессам 2011 г. устойчивость Савинки оказалась самой низкой, а НК Рокки – выше всех остальных гибридов.
-
6. Адаптивность гибрида Тристан к условиям засухи 2010 г. была выше средней, но к стрессам 2011 г. он оказался самым неустойчивым, тогда как гибрид Тутти проявил высокую устойчивость к стрессам в 2011 г., но был неустойчив к засухе в 2010 г. Гибрид Арена ПР оказался неустойчивым к стрессам как в 2010, так и в 2011 гг. Реализация его потенциала была выше средней только при максимальных индексах среды в 2011 г.
-
7. Устойчивость гибридов НК Ферти и НК Делфи к засухе в 2010 г. была низкой, а к стрессам в 2011 г. – повышенной, но они слабо реагировали на улучшение условий в 2011 г.
-
8. Гибриды НК Конди, НК Армони проявили превышающие средние уровни устойчивости к стрессам в 2010 и 2011 гг. при минимальных индексах среды и наиболее высокие потенциалы урожайности при максимальных индексах в оба года, а НК Брио только в 2011 г. По сравнению с
НК Брио его устойчивый к гербицидам аналог НК Неома оказался неустойчивым к стрессам в 2010 и 2011 гг. и проявил высокий потенциал урожайности только в 2011 г. при наибольших индексах среды.
-
9. Использованием для оценки нестабильности генотипов коэффициента вариации CV i вместо стандартного отклонения S i не достигается независимость оценок от средних величин признака, но при этом утрачивается часть информации, что ведет к низкой эквивалентности оценок S i и CV i . Предложен независимый от средних величин параметр нестабильности Ŝ i ,вычисленный как отклонение значений S i от их регрессий на средние величины.
-
10. Показана и объяснена высокая эквивалентность коэффициента регрессии на индексы среды b i с параметрами S i и Ŝ i , причем отрицательным значениям Ŝ i соответствует b i ˂ 1,0, положительным – b i ˃ 1,0. У гибридов с разными реакциями к стрессам в 2010 и 2011 гг. меняются в эти годы величины и знаки Ŝ i.
-
11. Анализом регрессии оценок симиляр-ности Сd i на значения остаточных дисперсий S2d i с предварительной стратификацией данных показано, что на величины Сd i отрицательно влияют величины S2d i и положительно – b i .
