Особенности клеточной структуры листьев культурного и дикорастущего подсолнечника Helianthus annuus L

Автор: Усатов А.В., Федоренко Г.Ф., Машкина Е.В., Тарасова В.А., Маркин Н.В.
Статья в выпуске: 2 (141), 2009 года.
Бесплатный доступ
Приведена сравнительная характеристика строения листовой ткани и ультраструктуры клеток культурной и дикорастущей форм подсолнечника Helianthus annuus L. Показано, что искусственный отбор способствовал формированию у культурной формы высокоэффективной структуры листовой паренхимы, обеспечивающей, прежде всего, интенсивный рост растений и их семенную продуктивность.
Подсолнечник, дикорастущая форма, ультраструктура, пластида, митохондрия, пероксисома
Короткий адрес: https://sciup.org/142150905
IDR: 142150905
Peculiarities of cellular structure of leaves in cultivated and wild sunflower Helianthus annuus L
The cultivated and wild forms of Helianthus annuus L. were studied. The comparative characteristic of the leaf structure and the cellular ultrastructure of these forms was done. It is shown that artificial selection resulted in formation of highly effective leaf parenchyma at cultivated sunflower that promotes the intensive development of plants and their seed productivity.
Текст научной статьи Особенности клеточной структуры листьев культурного и дикорастущего подсолнечника Helianthus annuus L
Введение. Главной целью искусственного отбора сельскохозяйственных культур является увеличение биомассы или семенной продуктивности растений. Однако конкретные пути достижения этой цели могут быть различны [1]. Известно, что основные физиологические характеристики организмов определяются особенностями организации клеток и внутриклеточных органелл [2, 3]. Поэтому целесообразно использовать культурные и дикорастущие формы одного вида в качестве моделей для изучения структурно-функциональных отношений, в частности различий в гомологичных органах и клеточных структурах растений. В связи с этим целью работы является сравнительный анализ структурной организации листовой ткани, клеток и энергетических клеточных органелл – хлоропластов и митохондрий у растений дикорастущей и культурной форм подсолнечника Helianthus annuus L.
Материалы и методы. Объектом исследования служили инбредные линии культурной (линия 3629) и дикорастущей форм подсолнечника H. annuus L . из генетической коллекции подсолнечника НИИ биологии ЮФУ, выращенные в полевых условиях на 10-метровых делянках площадью питания 60 × 60 см. В фазе появления 3-4 пары листьев измеряли высоту проростков. Содержание хлорофиллов определяли по общепринятой методике [4].
В экспериментах использовали высечки (1×2 мм) из 2-3 пары листьев. Анализировали клетки паренхимы среднего участка листа. Ультраструктурное исследование проводили с помощью модифицированного нами метода глутаро-осмиевой фиксации материала с последующей заливкой в эпон-аралдит [5].
Ультратонкие срезы получали на приборе EM UC26 (Leica, FRG) и дополнительно контрастировали солями тяжёлых металлов, а их исследование проводили в электронном микроскопе Tecnai 12 (Phillips, Holland). Полутонкие срезы толщиной 0,5-1 мкм для светооптического наблюдения дополнительно окрашивали метиленовым синим и исследовали в светооптическом микроскопе Jnaval (FRG).
Морфометрический анализ светооптических изображений включал подсчёт площади клеток и их количества, площади межклеточного пространства на единицу площади листовой ткани. На электронограммах определяли среднюю площадь среза хлоропласта и его внутренних мембран, отношение площади внутренних мембран к площади среза пластид, а также число тилакоидов на единицу площади внутренних мембран и количество пластид на единицу площади среза листа. Измерения площади клеточных фрагментов проводили с помощью специальной программы для количественного анализа микроизображений Olimpus Soft Imagin Solution ITEM. Достоверность полученных данных оценивали по t-критерию Стьюдента [6].
Результаты и обсуждение. Культурный и дикорастущий подсолнечник в первую очередь различается по габитусу. Если культурная форма представляет собой одностебельное растение с одним соцветием, то для дикорастущего подсолнечника характерна кустовая форма с высокой степенью ветвления, представленной ветвями от 1-го до 4-х порядков, и с варьирующим количеством соцветий (от 40 до 80 шт.), расположенных как на концах ветвей, так и в пазухах листьев. Несмотря на это, по семенной продуктивности одного растения дикорастущий подсолнечник в несколько раз уступает культурной форме (линия 3639).
Исследуемые формы различаются между собой и по динамике роста. В начале вегетации дикорастущая форма развивается медленно. Так, на стадии формирования 3-4-й пары листьев ее проростки в 2,5 раза уступают по высоте культурным растениям. Однако, начиная с фазы цветения, растения дикорастущей формы продолжают интенсивный рост, в то время как культурный подсолнечник в фазе цветения практически его прекращает. Изменение динамики развития культурных форм создает предпосылки для быстрого формирования биомассы, более раннего и выровненного созревания семян и в целом сокращения периода вегетации.
Увеличение скорости роста культурной формы подсолнечника может быть связано с изменением структурной организации как листовой ткани в целом, так и изменениями в структуре энергетических органелл – хлоропластов и митохондрий. Анализ светооптических фотографий клеток листовой ткани культурной формы показал, что мезофилл имеет четко выраженное деление на столбчатую и губчатую паренхимы. Столбчатая паренхима представлена 2-мя рядами достаточно плотно и упорядоченно (одинаково пространственно-ориентированных) прилегающих друг к другу клеток цилиндрической формы. Клетки губчатой паренхимы более округлы. Средняя площадь клетки столбчатой паренхимы в 1,5 раза превышает площадь клетки губчатой паренхимы. Однако количество клеток на единицу площади в губчатой паренхиме больше, чем в столбчатой (табл. 1).
У дикорастущей формы граница между столбчатой и губчатой паренхимами выражена менее чётко, чем у культурных растений. Скопления клеток эпидермиса образуют значительные по своим размерам (до трети толщины листовой пластинки) выступы и ворсинки, особенно многочисленные в верхнем слое. Средняя площадь клеток столбчатой ткани меньше по сравнению с клетками культурной формы подсолнечника, меньше и средняя площадь межклеточника (табл. 1). Форма клеток губчатой паренхимы более вытянута по сравнению с предыдущим вариантом, а их площадь и площадь межклеточника также достоверно меньше, чем у культурной формы.
Таблица 1 – Цитоморфологический анализ листовой ткани растений культурной и дикорастущей форм подсолнечника Helianthus annuus L.
|
Показатель |
Культурная форма |
Дикорастущая форма |
|
|
Площадь среза клетки (S), 10-8 см2 |
Столбчатая паренхима |
0,61 ± 0,04 |
0,46 ± 0,05* |
|
Губчатая паренхима |
0,42 ± 0,02 |
0,34 ± 0,03* |
|
|
Площадь среза межклеточного |
Столбчатая паренхима |
0,24 ± 0,02 |
0,17 ± 0,02* |
|
пространства (S), 10-8 см2 |
Губчатая паренхима |
0,29 ± 0,02 |
0,22 ± 0,01** |
|
Количество клеток на единицу |
Столбчатая паренхима |
1,26 ± 0,03 |
1,76 ± 0,04*** |
|
площади (1∙х 10-8 см2) |
Губчатая паренхима |
1,98 ± 0,04 |
2,24 ± 0,03 *** |
Примечание: *** – достоверные отличия по сравнению с культурной формой при P<0,001;
** – достоверные отличия по сравнению с культурной формой при P<0,01;
* – достоверные отличия по сравнению с культурной формой при P<0,05.
Таким образом, выявленные различия в площади межклеточного пространства столбчатого и губчатого слоёв листьев культурной формы, относительно дикорастущей, дают основание предположить наличие в них более развитой системы межклетников, позволяющей обеспечивать интенсивные процессы водо- и газообмена. Более того, высокая степень упорядоченной организации клеток палисадной паренхимы культурной формы предполагает оптимальные условия распределения светового потока в хлоропластах по сравнению с дикорастущими растениями для обеспечения эффективного процесса фотосинтеза. Дополнительное снижение уровня поглощенной фотосинтетически активной радиации дикорастущими растениями может быть вызвано интенсивным опушением их листьев, представленным многочисленными ворсинками и волосками, расположенными главным образом на верхней поверхности листовой пластинки.
Интересно отметить, что количество зеленых пигментов в листьях растений культурной (8,9 ± 0,3 мкг/г) и дикорастущей (8,4 ± 0,4 мкг/г) форм подсолнечника сходно.
Электронно-микроскопическое исследование хлоропластов культурных растений показало, что форма органелл в основном овальная с некоторыми изгибами и инвагинациями (рис. 1а). Внутри органелл можно видеть хорошо развитую систему мембран, состоящую из тилакоидов гран и тилакоидов стромы (рис. 1б). Подобный характер ультраструктуры хлоропластов свидетельствует об их высокой функциональной активности. Данные морфометрического анализа приведены в таблице 2.
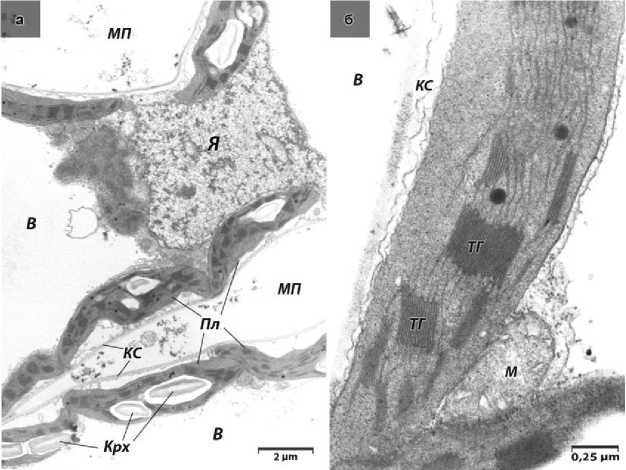
Рисунок 1 – Электронограммы культурной формы подсолнечника 3629 : (а) участки 2-х контактирующих клеток; (б) фрагменты 2-х пластид; В – клеточная вакуоль; КС – клеточная стенка; Крх – крахмальные зерна; М – митохондрия; МП – меж-клеточное пространство; Пл – пластида; ТГ – тилакоиды гран; Я – ядро.
Таблица 2 – Морфометрический анализ срезов хлоропластов культурной и дикорастущей форм подсолнечника
|
Линия |
Площадь среза (µm2) |
Отношение площади внутренних мембран к площади хлоропласта |
Количество тилакоидов на единицу площади внутренних мембран (2,8∙102µm2) |
Количество пластид на единицу площади среза листа (1,8∙103µm2) |
|
|
хлоропласт |
внутренние мембраны |
||||
|
Культурная форма |
15,7 ± 1,4 |
4,5 ± 0,3 |
0,29 |
20,3 ± 1,4 |
24,1 ± 1,3 |
|
Дикорастущая форма |
13,9 ± 2,5 |
4,2 ± 0,4 |
0,30 |
18,1 ± 1,9 |
19,8 ± 1,5* |
Примечание: *достоверно отличается от значений культурной формы (уровень значимости р < 0,05)
В полостях и изгибах, образованных хлоропластами, можно наблюдать плотно прилегающие митохондрии округлой и эллипсовидной формы. Среди них преобладают органеллы округлой формы, содержащие просветленный матрикс (рис. 1б).
По своей форме и ультраструктурной организации хлоропласты дикорастущего подсолнечника отличаются более высокой степенью гетерогенности. Так, наряду с пластидами овальной формы (схожей с культурным подсолнечником, встречаются органеллы значительно вытянутые относительно продольной оси (рис. 2а).
Средняя площадь среза пластид составляет 13,9 ± 2,5 µm2, что достоверно неотличимо от соответствующего показателя культурной формы (15,7 ± 1,4 µm2). Отношение площади внутренних мембран к площади пластиды и количество тилакоидов на единицу площади внутренних мембран у дикорастущего и культурного подсолнечника также находится на одном уровне (табл. 2). Однако количество хлоропластов на единицу площади среза листовой пластинки (2-я пара листьев) у дикорастущей формы на 18 % меньше, чем у растений линии 3629.
Крахмальные зёрна в строме дикорастущей формы более многочисленны по сравнению с культурной, отмечено значительное число органелл, содержащих до 4-х и более единиц (рис. 2б). Часто внутренняя оболочка хлоропластов дикорастущего подсолнечника образует большое число трубчатых выростов, располагающихся в периферической части пластиды – периферический ретикулум (рис. 2в).
Среди митохондрий дикорастущей формы преобладают органеллы эллипсовидной и округлой формы (рис. 2в, 2г). Строма некоторых пластид образует глубокие инвагинации, в которых могут быть расположены слегка набухшие митохондрии (рис. 2г). Форма таких митохондрий более округлая, чем те, которые расположены вне стромы пластиды, матрикс просветлён, кристы малочисленны, т.е. налицо ультраструктурные признаки повышенной функциональной нагрузки, испытываемой этими органеллами [7].
Интересно отметить, что в цитоплазме дикорастущего подсолнечника, в отличие от культурного, отмечено значительное количество пероксисом. Как правило, они образуют «триады» – комплекс из трёх органелл: митохондрия, пероксисома и пластида (рис. 2г). Пероксисомы фотосинтезирующих тканей метаболизируют гликолевую кислоту (гликолат). Классическая схема метаболизма гликолата включает согласованную работу хлоропласта, пероксисомы и митохондрии, что морфологически отражено на наших электронных микрофотографиях как ассоциация этих органелл. Метаболизм гликолата играет ключевую роль в фазу развития растений, когда осуществляется переход от использования запасных питательных веществ к фотосинтетическому метаболизму [8, 9].
И, напротив, в литературе описаны растения с нарушением фотодыхания, при этом у них наблюдается нарушение взаимодействия между пероксисомами и хлоропластами, что приводит к нарушению потока метаболитов [10]. Так как интенсивно протекающий гликолатный цикл может сильно снизить продуктивность С3 – растений (к которым относиться и подсолнечник), то некоторые селекционеры считают важной задачей выведение селькохозяйственных сортов, у которых этот процесс подавлен [11].
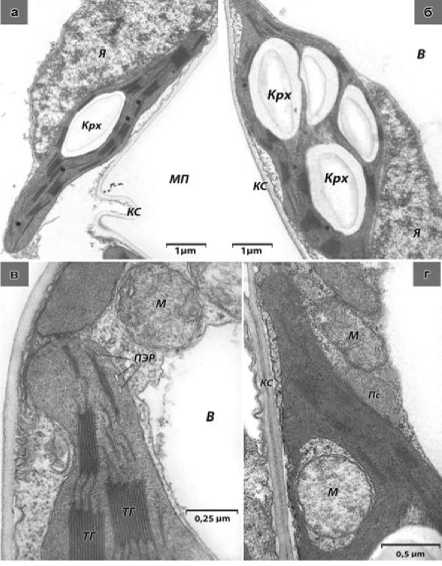
Рисунок 2 – Электронограммы дикорастущего подсолнечника: а) участок клетки с хлоропластом удлинённой формы; б) хлоропласт с 4-я крахмальными зёрнами; в) участок цитоплазмы клетки, содержащей фрагменты пластиды с везикулами периферического ретикулума и митохондриями округлой формы; г) участок цитоплазмы клетки, в котором расположены плотно-контактирующие митохондрия, пероксисома и хлоропласт, «инваги-нирующий» набухшую митохондрию.
В – клеточная вакуоль; КС – клеточная стенка; Крх – крахмальные зерна; М – митохондрия; МП – межклеточное пространство; Пл – пластида; ПС – пероксисома; ТГ – тилакоиды гран; Я – ядро; ПЭР -периферический ретикулум
Сравнительный анализ ультраструктурной организации митохондрий дикорастущего и культурного подсолнечника свидетельствует, что хотя существенных различий в тонком строении не выявлено, их локализация в цитоплазме и способ контакта с другими органеллами, в частности с хлоропластами иной, что, вероятно, связано с изменением (усилением) их функциональной активности.
Очевидно, выявленные нами особенности пространственной организации и тонкого строения клеток и органелл лежат в основе клеточных механизмов, определяющих различную степень продуктивности и устойчивости культурной и дикорастущей форм подсолнечника к действию внешних факторов среды. Функциональная активность хлоропластов и митохондрий, как основных продуцентов энергии в растительной клетке должна быть взаимосвязана и взаимозависима. На примере подсолнечника видно, что наличие активно функционирующих митохондрий в ассоциации с многочисленными пероксисомами и функционально активными хлоропластами отражает высокую интенсивность гликолатного цикла у дикорастущей формы по сравнению с культурной. У последней, судя по незначительному количеству пероксисом, гликолатный цикл в значительной степени подавлен. Вероятно, это является причиной того, что дикорастущий подсолнечник, не обладая высокой скоростью роста и продуктивностью, проявляет устойчивость к различным экстремальным факторам среды. На влияние пероксидазного метаболизма на резистентность растений к стрессу указывают и другие авторы [12].
Выводы. Многолетний искусственный отбор дикорастущего подсолнечника привел к формированию у культурной формы высокоэффективной структуры листовой ткани, обеспечивающей, прежде всего, интенсивный рост растений и их семенную продуктивность.
Исследование выполнено при финансовой поддержке Министерства науки и образования РФ (грант «Развитие научного потенциала высшей школы (2009-2010)» № 2.1.1/4947).
