Особенности микросателлитных локусов панели, рекомендованной ISAG-FAO для генотипирования овец (Ovis aries L.)
")
Автор: Эркенов Т.А., Алтухова Н.С., Гребенчук А.Ю., Глазко Т.Т., Косовский Г.Ю.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Панели, предложенные ISAG и FAO для генотипирования микросателлитных локусов, получили широкое распространение в исследованиях, выполненных на сельскохозяйственных животных, в том числе на домашней овце (Ovis aries L.). К настоящему времени по десяткам и сотням пород разных видов накоплено большое количество данных, представляющих полученные результаты усредненно по разным микросателлитным локусам как дополнительные породные характеристики. В то же время до сих пор остается невыясненным источник противоречия между консервативностью праймеров и высоким полиморфизмом самих микросателлитных локусов. Практически отсутствуют исследования, позволяющие связывать полиморфизм микросателлитов панели с хорошо документированными межпородными различиями по фенотипическим характеристикам. Для выяснения потенциальных причин такого несоответствия в представленной работе на примере генотипирования овец карачаевской породы с использованием панели ISAG (ISAG, 2021) мы оценили возможные связи между полиморфизмом отдельных микросателлитов и их различиями по нуклеотидному составу элементарной единицы тандемного повтора — нуклеотидному кору, его локализации в межгенном или интронном участках белок-кодирующих генов, принадлежности фланговых белок-кодирующих генов к разным метаболическим путям. Панель состоит их 12 микросателлитов и включает четыре микросателлита с коровым мотивом AC, три — с CA, три — с TG и два — с GT. Число выявленных наблюдаемых аллелей по разным микросателлитам варьировало от 7 до 17 и не зависело от нуклеотидной последовательности коровых мотивов. С использованием программ Blastn и рекомендованных к панели ISAG праймеров выполнена их локализация в геноме овцы (ARS-UI_Ramb_v3.0, 2023). Все четыре микросателлита с наибольшим числом аллельных вариантов — INRA023 (предположительно), INRA005, INRA172 (по 15 аллелей) и OarFCB20 (17 аллелей) локализовались в межгенных пространствах независимо от корового мотива. Наименьшее число аллельных вариантов (семь) выявлено по локусам ETH152 и MAF214; первый расположен близко к белок-кодирующему гену MPPED1 (вовлечен в регуляцию клеточной дифференцировки) (K. Gupta и соавт., 2023), второй локализован в интроне гена TTC33 (контролер стабильности белков) (L.R. Serrano и соавт., 2025). Три микросателлита — McM42, McM527 и INRA006 локализовались в межгенных пространствах, остальные три — в интронах белок-кодирующих генов. Все белок-кодирующие гены, тесно связанные с микросателитами панели, относились к участникам фундаментальных метаболических путей внутриклеточного жизнеобеспечения. По-видимому, именно это и объясняет противоречия между консервативностью нуклеотидных последовательностей праймеров и высоким полиморфизмом микросателлитов. Учитывая интенсивность искусственного отбора, регулярные бонитировки овец, можно ожидать, что носители мутаций в последовательностях праймеров, тесно сцепленных с функционально важными белок-кодирующими генами, элиминируются из генофонда популяций. Сравнение данных, полученных у разных пород овец, генотипированных по микросателлитам с использованием этой же панели, свидетельствуют об относительно пониженной эффективности их использования для локализации генов-кандидатов контроля изменчивости хозяйственно ценных признаков, но очевидной важности для исключения ошибок происхождения животных.
Микросателлиты, полиморфизм, генетическое сцепление, метаболические пути, отбор
Короткий адрес: https://sciup.org/142247687
IDR: 142247687 | УДК: 636.012:636.32/.38:575.174.015 | DOI: 10.15389/agrobiology.2026.2.260rus
Features of microsatellite loci in the ISAG-FAO panel for sheep (Ovis aries L.)
The panels of polymerase chain reaction conditions and primers for genotyping microsatellite loci proposed by ISAG and FAO have been widely used for a number of farm animal species, including domestic sheep (Ovis aries L.). A large amount of data has been accumulated across dozens and hundreds of breeds, in which the obtained results are presented as averages for different microsatellite loci as additional breed-specific characteristics. However, the source of the discrepancy between the conservatism of primers and the high polymorphism of the microsatellite loci themselves remains unclear, as does the virtual absence of studies linking the polymorphism of the microsatellite panel with well-documented interbreed differences in phenotypic characteristics. To elucidate the potential sources of this discrepancy, this study assessed possible links between the polymorphism of individual microsatellites and their differences in the nucleotide composition of the tandem repeat elementary unit (nucleotide core), its localization in the intergenic or intronic regions of protein-coding genes, and the affiliation to flanking regions of genes involved in different metabolic pathways. As an example, Karachay sheep were genotyped by the ISAG panel (ISAG, 2021). The panel consists of 12 microsatellites and includes 4 microsatellites with the AC core motif, 3 with CA, 3 with TG, and 2 with GT. The number of observed alleles identified for different microsatellites varied from 7 to 17 and did not depend on the core motifs. Using the Blastn software and primers recommended for the ISAG panel, they were localized in the sheep genome (ARS-UI_Ramb_v3.0, 2023). All four microsatellites with the highest number of allelic variants were localized in the intergenic spaces: INRA023 (presumably), INRA005, INRA172 (15 alleles) and OarFCB20 (17 alleles), regardless of the motif. The smallest number of allelic variants, 7 was identified at the ETH152 and MAF214 loci. The first one is located close to the protein-coding gene MPPED1 (involved in the regulation of cell differentiation (K. Gupta et al., 2023), the second is localized in the intron of the TTC33 gene (protein stability controller (L.R. Serrano et al., 2025). Three microsatellites McM42, McM527 and INRA006 were localized in the intergenic spaces, the remaining three in introns of protein-coding genes. All protein-coding genes closely associated with the microsatellites of the panel were involved in fundamental metabolic pathways of intracellular life support. Apparently, the contradictions between the conservatism of the nucleotide sequences of primers and the high polymorphism of microsatellites may be related to genetic linkages between primers and life important genes. Given the intensity of artificial selection and regular sheep grading, it can be expected that carriers of mutations in primer sequences closely linked to functionally important protein-coding genes are quickly eliminated from the gene pool of populations. Comparison of data obtained in different sheep breeds genotyped for microsatellites using the same panel indicates a relatively reduced efficiency in localizing candidate genes controlling variation in economically valuable traits, but their obvious importance in eliminating errors in animal origin.
Текст научной статьи Особенности микросателлитных локусов панели, рекомендованной ISAG-FAO для генотипирования овец (Ovis aries L.)
Микросателлиты, или короткие тандемные повторы (simple tandem repeats STR или simple sequence repeat SSR), — хорошо известные ДНК-маркеры, которые используются в различных областях, требующих высокополиморфных и локус-специфических генетических систем. Широко распространено их применение, например, в селекционных программах в целях исключения при работе с сельскохозяйственными видами животных ошибок происхождения, анализе сцепления между различными локусами, контроле популяционно-генетических характеристик пород (1). Исключение ошибок происхождения критично для оценки генетических параметров и племенной ценности животных в племенной работе, прогнозирования 260
генетического прогресса и управления генетическим разнообразием.
Длины элементарных единиц тандемных повторов микросателлит-ных локусов (нуклеотидного кора) варьируют от 1 до 6 пар нуклеотидов (п.н.). К настоящему времени создана база данных для разных видов животных и растений, в которой представлена частота встречаемости микро-сталлитных локусов, различающихся по длинам элементарной единицы тандемного повтора и локализации в разных геномных участках видоспецифичных геномов (MSBD, .
Международное сообщество генетики животных (International Society for Animal Genetics, ISAG) и Международная организация по продовольствию и сельскому хозяйству (Food and Agriculture Organization, FAO) разработали панели для генотипирования микросателлитных локусов у животных, в частности для домашней овцы (Ovis aries L.) .
Панель, разработанная для домашней овцы , включает 12 динуклео-тидных микросателлитных локусов с описанием необходимых праймеров (F — лидирующий, R — терминирующий), температуры отжига, условий проведения полимеразной цепной реакции и потенциальных длин ампли-фицируемых фрагментов. При составлении панели микросателлитов учитывались несколько параметров: высокая степень полиморфизма и разнообразия в популяциях животных соответствующих видов для удобства выявления различий между индивидами; локализация в надежно секвениро-ванных геномных участках; определенная эволюционная консервативность последовательностей праймеров; возможность амплификации одновременно нескольких локусов; имеющиеся данные об ассоциациях между полиморфизмом локусов и фенотипическими характеристиками. ISAG для каждого вида рекомендовало соответствующую панель после длительного согласования по перечисленным параметрам с участием многих экспертов.
Разработанные панели широко использовались в течение последних 20 лет, и с их помощью получен большой объем данных о генотипах разных пород, в частности почти по 200 породам у овец (2).
В то же время накопленный потенциал выполненных исследований до сих пор остается недостаточно использованным. Так, не известны причины различий по числу аллельных вариантов, непрерывности рядов их изменчивости (наличие/отсутствие наиболее часто встречающихся аллелей, условно называемых приватными или эффективными) у разных микроса-теллитных локусов с одним и тем же коровым мотивом, их изменчивости по индексу фиксации, а также высокой консервативности их флангов (праймеров) у десятков различных пород и даже у близкородственных видов.
В поисках возможных источников выявляемой консервативности флангов STR представляет интерес проанализировать связь этих событий с различиями в коровых мотивах STR, с их локализацией в различных аутосомах, интронах и межгенных пространствах и генетическим сцеплением с функционально различными белок-кодирующими генами. Подобных исследований в доступной литературе мы не обнаружили.
Целью настоящей работы было выполнение анализа полиморфизма микросателлитных локусов стандартной панели ISAG для домашней овцы на примере адаптированной к высокогорному пастбищному разведению популяции овец карачаевской породы.
Методика. Исходные результаты генотипирования овец карачаевской породы опубликованы ранее (3). В работе использовали нуклеотидные последовательности генома Ovis aries (ARS-UI_Ramb_v3.0, 2023) и последовательности праймеров панели ISAG, рекомендованных для генотипирования овец (4, 5). Позиционирование последовательностей праймеров в хромосо- мах и районах генома овцы определяли с использованием программы .
Информация об отобранных структурных генах получена из международной базы данных GenBank . Их функциональную активность изучали с помощью базы данных KEGG (Kyoto Encyclopedia of Genes and Genomes, . Сравнение нуклеотидных последовательностей выполняли методами выравнивания, используя программу .
Статистическая обработка данных проводилась с использованием программы GenAlEx 6.5 (6), MS Excel, для оценки статистической достоверности различий средних арифметических выборок применяли t - критерий Стьюдента.
Результаты. На первом этапе наших исследований мы подразделили микросателлиты панели ISAG по нуклеотидным последовательностям элементарной единицы STR. В геноме домашней овцы наиболее часто встречаются микросателиты с динуклеотидным мотивом AC . В панели ISAG к таким микросателлитам принадлежат локусы McM042, INRA063, INRA023 и ETH152. Три микросателлита несут тандемные повторы CA (INRA006, CSRD247 и MAF065); три микросателлита — тандемные повторы TG (McM527, INRA172, OarFSB20); два микросателлита — повторы GT (INRA005 и MAF214). То есть из коровых мотивов микросателлитов панели исключены микросателлиты, предрасположенные к формированию пурин-пиримидиновых треков (AG, GA и TC, CT), среди которых, судя по базе данных микросателлитов, в геноме овцы микросателлит AG второй по частоте встречаемости после AC.
С использованием программы GenAlEx 6.5 (7) у 50 овец карачаевской породы мы рассчитали общее число аллелей и число эффективных аллелей, индекс разнообразия Шеннона, наблюдаемую и ожидаемую гетерозиготность, индекс фиксации (табл. 1). Число наблюдаемых аллельных вариантов (Na) у группы STR с коровым мотивом AC — 10,0 ± 1,7; CA — 9,7±0,7; TG — 13,7±2,2 и GT — 11,0±3,0.
Полученные данные свидетельствуют о том, что полиморфизм применяемых для генотипирования микросателлитных локусов не зависит от их коровых мотивов (см. табл. 1).
Для того чтобы определить возможное влияние фланговых последовательностей, окружающих микросателлит, с использованием версии секвениро-ванного генома домашней овцы GCF_016772045.2 (ARS-UI_Ramb_v3.0) и соответствующих программ BLASTn , был выполнен поиск хромосом и координат локализации праймеров, рекомендованных ISAG для генотипирования овец. По нашим данным, полная гомология к последовательности лидирующего праймера микросателлита ETH152/D5S2 (F TACTCGTAGGGCAGGCTGCCTG, подчеркиванием выделен потенциальный квадруплекс G4) обнаруживается в хромосоме 3. При поиске в геноме овцы участка, фланкированного рекомендованной ISAG парой праймеров для локуса ETH152/D5S2, с помощью программы Primer-blast выяснилось, что такой участок действительно есть на хромосоме 3 в районе с координатами 220143855-220144049 п.н. И очень близко к нему расположен белок-кодирующий ген MPPED1 (metallophosphoesterase domain containing 1, NC_056056.1) с координатами 220092444-220179131 п.н., то есть до начала гена MPPED1 в межгенном пространстве. Однако R праймер в программе Primer-blast отличался от рекомендованного на две мононук-леотидные замены, одна разрушала G4-квадруплекс, другая создавала его в иной позиции: R праймер от ISAG — 5´-GAGACCTCAGGGTTGGTG-ATCAG-3´, в Primer-blast — 5´-GAGACCTCAGGGTTGTTGGTCAG-3´.
1. Полиморфизм 12 микросателлитных локусов из панели ISAG у овец ( Ovis aries L.) карачаевской породы 2018-2022 годов рождения (ООО Племенной завод «Махар», Карачаево-Черкесская Республика, 2025 год)
|
Показатель |
Коровый мотив AC |
Коровый мотив CA |
Коровый мотив TG |
Коровый мотив GT |
||||||||
|
STR |
McM042 |
INRA063 |
INRA023 |
ETH152 |
INRA006 |
CSRD247 |
MAF065 |
McM527 |
INRA172 |
OarFCB20 |
MAF214 |
INRA005 |
|
Na |
9 |
10 |
15 |
7 |
9 |
9 |
11 |
9 |
15 |
17 |
7 |
15 |
|
Ne |
2,790 |
4,307 |
7,133 |
3,482 |
2,483 |
2,463 |
6,061 |
4,425 |
5,787 |
10,163 |
2,809 |
9,542 |
|
I |
1,325 |
1,845 |
2,246 |
1,493 |
1,368 |
1,338 |
1,998 |
1,703 |
2,134 |
2,489 |
1,236 |
2,421 |
|
H o |
0,840 |
0,660 |
0,700 |
0,680 |
0,460 |
0,500 |
0,960 |
0,800 |
0,820 |
0,800 |
0,600 |
0,960 |
|
H e |
0,642 |
0,768 |
0,860 |
0,713 |
0,597 |
0,594 |
0,835 |
0,774 |
0,827 |
0,902 |
0,644 |
0,895 |
|
F |
- 0,309 |
0,140 |
0,186 |
0,046 |
0,230 |
0,158 |
- 0,150 |
-0,034 |
0,009 |
0,113 |
0,068 |
- 0,072 |
П римечание.
Na — число выявленных аллелей,
Ne — число эффективных аллелей = 1/ ∑ p i 2 , где p i — частоты аллелей;
I — индекс разнообразия Шеннона = - ∑ (p i ½ ln p i );
H o — наблюдаемая гетерозиготность;
H e — ожидаемая гетерозиготность = 1 - ∑ p i 2 ;
F — индекс фиксации = (H e - H o )/H e = 1 - (H o /H e ).
Следует отметить, что нуклеотидные последовательности, гомологичные отдельным участкам праймеров к этому микросателлиту, встречаются почти во всех хромосомах овцы и чаще всего сохраняют сочетание нуклеотидов, потенциально предрасположенное к формированию G4-квад-руплекса, в том числе на хромосоме 5 (5´-A GGG CA GG CTGCCTG-3´).
Известно, что G4-квадруплексы представляют собой распространенные мишени связывания факторов регуляции транскрипции, которые содержат цинковые пальцы (10), кроме того, G4-квадруплексы оказывают влияние на геномную нестабильность (11). По-видимому, сложная и динамичная структурно-функциональная роль G4-квадруплексов в генетическом аппарате может быть одной из причин того, что у большого числа разных пород консервативность праймеров при генотипировании вступает в явное противоречие с трудностями, возникающими при определении хромосомной локализации для ряда микросателлитных локусов.
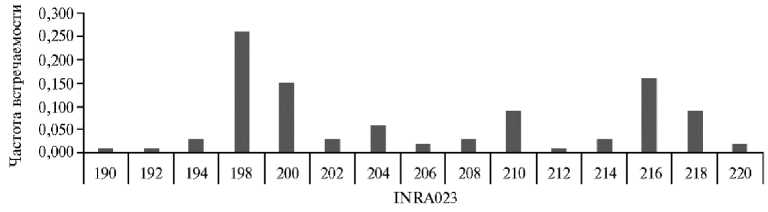
Рис. 1. Частота встречаемости аллельных вариантов локуса INRA023 из панели ISAG у овец ( Ovis aries L.) карачаевской породы 2018-2022 годов рождения (ООО Племенной завод «Махар», Карачаево-Черкесская Республика, 2025 год).
Следующий после ETH152 микросателлитный локус — INRA023, с которым обнаружились похожие сложности при локализации его праймеров в геноме овцы, не несет последовательностей G4-квадруплексов (F GAGTAGAGCTACAAGATAAACTTC и R TAACTACAGGGTGTTAGATG-AACTC), но так же, как и в случае праймеров локуса ETH152/D5S2, протяженные участки гомологичных последовательностей встречаются во всех хромосомах овцы, особенно в хромосоме 2. В то же время результаты использования программы Primer-blast показали, что в геноме овцы отсутствуют участки, фланкированные этими последовательностями. По-види-мому, это обусловлено двумя причинами: низким числом пар GC в обоих праймерах (F — 37.50 %, R — 40.00 %) и распространенностью микросателлитов с коровым мотивом AC в межгенных районах . Предположению об относительно повышенной селективно нейтральной изменчивости соответствуют результаты генотипирования локуса INRA023: его принадлежность к четырем из 12 микросателлит-ных локусов с наибольшим числом аллельных вариантов (см. табл. 1) и присутствие почти всех потенциальных вариантов аллелей динуклеотидного микросателлита в исследованной выборке овец, за исключением только одного варианта длиной 196 п.н. (рис. 1).
С двумя другими микросателлитными локусами с коровыми мотивами AC — MCM042 и INRA063 таких проблем не возникало.
При локализации микросателлита INRA005 с коровым мотивом GT в референсном геноме овец породы рамбулье выяснилось, что праймеры, рекомендованные ISAG для генотипирования по этому локусу, отличаются от выявляемых с использованием программы Primer-blast: праймеры ISAG —
F TTCAGGCATACCCTACACCACATG, R AAATATTAGCCAACTGAAAA-CTGGG; полученные праймеры — F TTCAGGCACAGCCTACACCACATG и R AAATATTAGCCAATTGAAAACTGGG с локализацией в хромосоме 10 в координатах 73568590-73568722 п.н. И так же, как в случае микросателлита INRA023, микросателлит INRA005 относится к группе с наибольшим числом аллелей, и их представленность соответствует нейтральному распре- делению числа тандемных динуклеотидных повторов, за исключением аллеля длиной 119 п.н. (рис. 2).
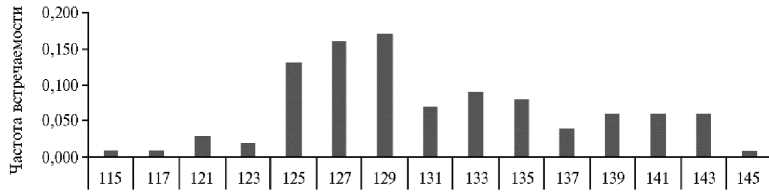
[NRA005
Рис . 2. Частота встречаемости аллельных вариантов локуса INRA005 из панели ISAG у овец ( Ovis aries L.) карачаевской породы 2018-2022 годов рождения (ООО Племенной завод «Махар», Карачаево-Черкесская Республика, 2025 год) .
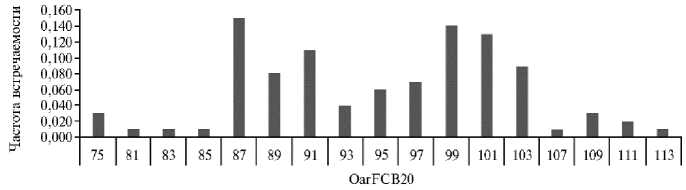
Рис. 3. Частота встречаемости аллельных вариантов локуса OarFCB20 из панели ISAG у овец ( Ovis aries L.) карачаевской породы 2018-2022 годов рождения (ООО Племенной завод «Махар», Карачаево-Черкесская Республика, 2025 год) .
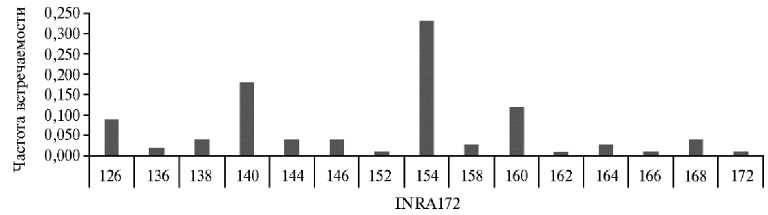
Рис. 4. Частота встречаемости аллельных вариантов локуса INRA172 из панели ISAG у овец ( Ovis aries L.) карачаевской породы 2018-2022 годов рождения (ООО Племенной завод «Махар», Карачаево-Черкесская Республика, 2025 год) .
На следующем этапе работы мы попытались уточнить особенности полиморфизма микросателлитных локусов в зависимости от их локализации в межгенных пространствах и в интронах. В результате получены следующие данные. Все четыре микросателлита с наибольшим числом аллельных вариантов локализовались в межгенных пространствах: INRA023 (предположительно), INRA005, INRA172 (по 15 аллелей) и OarFCB20 (17 аллелей). Распределение частот разных аллельных вариантов у трех микросателлит-ных локусов образовывало почти правильную последовательность, не поз- воляющую надежно выделить эффективные аллели (см. рис. 1-3), за исключением микросателлита INRA172 (рис. 4), локализованного между геном HIF1AN (hypoxia inducible factor 1 subunit alpha inhibitor), продукт которого участвует в подавлении регуляции сигнального пути Notch и индукции дифференцировки миобластов (12), и геном PAX2 фактора регуляции транскрипции (критический регулятор раннего эмбрионального развития) (13).
Наименьшее число аллельных вариантов было выявлено по локусам ETH152 и MAF214, причем первый локализован в хромосоме 3 близко к началу гена эстеразы MPPED1 (metallophosphoesterase domain containing 1, экспрессируется в головном мозге на ранних этапах развития, предполагается ее участие в регуляции клеточной дифференцировки) (14), второй — в интроне гена TTC33 (tetratricopeptide repeat domain 33), участвующего в стабилизации и убиквинизации неправильно свернутых белков в процессе их деградации и утилизации (15).
Три других микросателлита имели по 9 аллельных вариантов и локализовались в межгенных пространствах: McM42 между геном фактора регуляции транскрипции HNF4G (16) и геном, кодирующим сплайсосомную РНК (U6 сплайсосома, LOC114116512 U6 ); INRA006 между геном субъединицы GIRK3 критического модулятора калиевого канала (клеточного возбудителя) (17) и геном субъединицы ATP1A4 (18), контролирующей градиенты Na+ и K+ в плазматических мембранах; McM527 между генами тРНК цистеина ( TRNAC-GCA ) и COX7B — субъединицы 7 цитохром-С-оксидазы (ключевой фермент дыхательной цепи митохондрий) (19).
Оставшиеся три гена были локализованы в интронах белок-кодиру-ющих генов: INRA063 (10 аллелей) в интроне гена URI1 (prefoldin like chaperone, влияет на клеточную пролиферацию) (20); CSRD247 (9 аллелей) в интроне гена PHKB (phosphorylase kinase beta subunit, один из ключевых генов, вовлеченных в углеводный обмен) (21); MAF65 (11 аллелей) в интроне гена GRIK4 (glutamate receptor, ionotropic, kainate 4, кодирует субъединицу ионотропного глутаматного каинатного рецептора, участвующего в возбудимости клеток и влияющего на ранние этапы развития эмбрионов) (22). Число наблюдаемых аллельных вариантов (Na) у группы восьми STR, локализованных в межгенных пространствах, в среднем отличалось (статистически недостоверно) от выявленного у четырех STR, расположенных в интронах белок-кодирующих генов (12,0±1,4 против 9,3±0,8). В этом случае обнаруживается определенная тенденция к уменьшенному числу Na у STR, локализованных в инронах.
На основании полученных данных можно заключить, что на полиморфизм микросателлитных локусов, включенных в панель ISAG для генотипирования овец, не оказывают существенного влияния коровые последовательности элементарных единиц рассмотренных тандемных динуклеотид-ных повторов, но изменчивость числа аллельных вариантов заметно зависит от локализации микросателлита в межгенных пространствах или в интронах белок-кодирующих генов, а также от критичности связанных с ними биохимических функций белок-кодирующих генов (регуляция возбудимости нервных клеток, клеточная дифференцировка, ключевые звенья клеточного метаболизма).
Для того чтобы оценить, влияет ли аутосомная локализация микро-сателлитных локусов на их полиморфизм, далее были рассмотрены плотность и некоторые биохимические функции белок-кодирующих генов во флангах длиной 1 Мб (1 млн п.н.), окружающих соответствующий микро-сателлитный локус. Для анализа эта длина была выбрана в связи с предположением о том, что длина петли хроматина, объединяющая белок-кодиру-ющие гены с потенциально сходным механизмом регуляции транскрипции может составлять приблизительно 1 Мб (23). Как правило, длина топологически связанных геномных областей может варьировать от 100 Кб до 1 Мб и более (23, 24).
2. Хромосомная локализация и представленность белок-кодирующих генов во флангах рассмотренных микросателлитных локусов из панели ISAG ( длина фдангов 1 Мб) у овец ( Ovis aries L.) карачаевской породы 2018-2022 годов рождения (ООО Племенной завод «Махар», Карачаево-Черкесская Республика, 2025 год)
Примечание. 2? — предварительная локализация микросателлита INRA023 в хромосоме 2; ТФ —факторы регуляции транскрипции; оборот белков — белки, участвующие в модификациях и деградации других белков.
Судя по полученным данным, полиморфизм микросателлитов не зависел от плотности белок-кодирующих генов, локализованных во флангах каждого локуса длиной 1 Мб (табл. 2) и колебался от 4 (OarFCB20) до 37 генов (INRA172 и INRA006).
Следует также отметить, что при поиске участков гомологии к праймерам микросателлитных локусов в геноме домашней овцы обнаруживается, что короткие фрагменты такой гомологии встречаются в большинстве хромосом. Это позволяет предполагать, что некоторые из них представляют сайты связывания с факторами регуляции транскрипции, и именно этим объясняется их широкая представленность. Известно, что длина сайтов связывания транскрипционных факторов обычно составляет всего 6-12 п.н., причем в большинстве случаев они функционируют кооперативно (25, 26).
Обсуждая полученные результаты, отметим, что влияние разных факторов естественного отбора на полиморфизм микросателлитов достаточно подробно описан в литературе, в частности выявлено выраженное влияние географической широты разведения: чем она выше, тем ниже полиморфизм (27). По-видимому, сходное влияние может оказывать сложность факторов искусственного отбора: чем ниже специализация, чем больше признаков участвует в отборе, тем выше полиморфизм.
Имеющиеся данные свидетельствуют о том, что стандартная панель микросателлитных локусов ISAG, первоначально разработанная для исключения ошибок происхождения, может отражать общую интенсивность давления различных факторов искусственного и естественного отборов, тогда как для выявления породоспецифичных характеристик требуется сравнительный анализ полиморфизма отдельных микросателлитных локусов.
К настоящему времени появляются публикации, в которых полиморфизм микросателлитных локусов оказывается ассоциирован с изменчивостью фенотипических характеристик, например связь INRA006 с некоторыми показателями липидного обмена (с геном транспортера жирных кислот) (28, 29), INRA023 — с выходом баранины (30).
Выявленные ассоциации относительно слабы и касаются признаков, имеющих сложную полигенную природу. На этом основании ряд исследователей полагают, что полиморфизм по микросателлитным локусам используемой панели в целом мало соотносится с характеристиками по признакам продуктивности у овец (за исключением потенциальной связи между наблюдаемой гетерозиготностью и репродуктивной функцией) (31).
Полученные нами данные свидетельствуют о том, что в качестве основного источника ассоциаций полиморфизма микросателлитных локусов и фенотипических характеристик допустимо рассматривать биохимические функции генетически сцепленных с микросателлитами белок-кодирующих генов. В частности, этим можно было бы объяснить противоречие между консервативностью нуклеотидных последовательностей праймеров для микросателлитов у разных пород и полиморфизмом генетически сцепленных с такими праймерами микросателлитных локусов. Не исключено, что определенный вклад в эту изменчивость вносит разная локализация микросателлитов в межгенных пространствах и в интронах белок-кодирующих генов.
Анализ белок-кодирующих генов, генетически сцепленных с микросателлитами панели, позволяет полагать, что биохимическая функция этих генов настолько фундаментальна, что любой полиморфизм их кодирующих последовательностей приводил бы к глубоким нарушениям развития. Это следует из описаний болезней (в частности, у человека), связанных с мутациями рассматриваемых генов . При подобных нарушениях развития у овец это приводило бы к элиминации носителей таких мутаций на ранних стадиях постэмбрионального развития и тем более после начала регулярных бонитировок ягнят с 4-месячного возраста. По-видимому, в поисках ассоциаций между полиморфизмом микросателлитов панели, рекомендованной ISAG, необходимо учитывать влияние достаточно интенсивного искусственного отбора, проводимого с животными сельскохозяйственных видов.
Таким образом, панель микросателлитов, рекомендованная ISAG— FAO для генотипирования овец, включает 12 микросателлитных локусов, полиморфизм каждого из которых варьирует. Для выявления возможных причин наблюдаемых при этом различий на примере популяции карачаевской овцы выполнен анализ потенциального влияния на полиморфизм микросателлитов нуклеотидной последовательности их элементарной единицы, локализации праймеров для каждого микросателлита в межгенном 268
пространстве или в интронах белок-кодирующих генов, принадлежности белок-кодирующих генов, с которыми генетически сцеплены микросателлиты, к разным метаболическим путям. Получены данные, которые показывают, что на полиморфизм микросателлитов может влиять их локализациия в межгенном пространстве, а также генетическая сцепленность с генами белков, обеспечивающих основные биохимической функции. Полиморфизм микросателлитов не зависел от плотности локализации белок-коди-рующих генов во флангах микросателлитов длиной 1 Мб (примерная длина петли хроматина как единицы регуляции экспрессии генов). Можно ожидать, что отсутствие связей между полиморфизмом микросателлитов и изменчивостью ряда хозяйственно ценных признаков обусловлено элиминацией из генофонда мутаций по белок-кодирующим генам, вовлеченным в фундаментальные метаболические пути, которая происходит в результате действия факторов естественного и искусственного отборов. По-видимому, рекомендованные ISAG панели микросателлитных локусов, которые у сельскохозяйственных видов животных высокоэффективны для исключения ошибок происхождения в селекционно-племенной работе, намного менее перспективны для хромосомной локализации генов-кандидатов контроля изменчивости, обусловленной высокой интенсивностью искусственного отбора по фенотипическим признакам.
Авторы выражают сердечную благодарность к.б.н. Ларисе Михайловне Федоровой за интерес, проявленный к работе, плодотворную дискуссию и полезные советы при подготовке статьи к опубликованию.
ФГБОУ БО Северо-Кавказская государственная академия,
