Особенности морфогенеза яровой мягкой пшеницы в культуре in vitro в зависимости от условий произрастания

Автор: Никитина Е.Д., Хлебова Л.П.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Экология
Статья в выпуске: 2, 2015 года.
Бесплатный доступ
Изучено влияние условий произрастания 12 сортов яровой мягкой пшеницы на реализацию их морфогенетического потенциала в культуре незрелых зародышей. Установлена достоверная зависимость каллусо-, морфогенеза и регенерации растений от генотипического разнообразия сортов, условий выращивания донорных растений и генотип-средо- вого взаимодействия. Генотипическая специфичность исходного материала является доминирующим фактором, определяющим эффективность всех образовательных процессов in vitro (каллусогенеза - на 38 %, морфогенеза - на 33 %, регенерации растений - на 80 %). Реакция отдельных генотипов на условия выращивания доноров эксплантов специфична. Полевые условия наиболее благоприятны для генотипов с высоким уровнем регенерации, условия зимнее-весенней вегетации в теплице - для сортов с низким регенерационным потенциалом, так как повышают частоту выхода растений в 3,4-13,9 раза.
Мягкая пшеница, генотип, условия произрастания, незрелые зародыши, каллусогенез, морфогенез, регенерация
Короткий адрес: https://sciup.org/14113068
IDR: 14113068 | УДК: 581:633.11
Morphogenesis features of spring wheat in vitro culture depending on growing conditions
The effect of growth conditions in 12 spring wheat varieties upon the realization of their morphogenetic potential in the immature embryos culture has been studied. A significant dependence of callus formation, morphogenesis and plant regeneration upon the genotypic diversity of varieties, growing conditions of donor plants and genotype-environment interaction has been stated. Genotypic specificity of the initial material was a dominant factor in determining the effectiveness of all processes in vitro (callus formation by 38 %, morphogenesis by 33 %, plant regeneration by 80 %). The reactions of genotypes on growth conditions of donor explants were specific. Field conditions were the most favorable for genotypes with a high regeneration level, the conditions of the winter-spring vegetation in the greenhouse were the best for varieties with a low regenerative potential. The increasing the output of plants in 3,4-13,9 times were obtained.
Текст научной статьи Особенности морфогенеза яровой мягкой пшеницы в культуре in vitro в зависимости от условий произрастания
Введение. Феномен тотипотентности соматических растительных клеток позволяет индуцировать морфогенетические процессы в культуре in vitro , в результате которых de novo образуются ткани и органы, способные формировать целый организм [5]. Известно, что морфогенез является одним из наиболее сложных и трудно регулируемых событий в культуре ткани, особенно для семейства Gramineae. Результаты многих исследований [2, 21], а также данные наших экспериментов [3, 11, 13, 14] продемонстрировали несомненную генотипическую обусловленность данного процесса. Установлена зависимость способности к реализации генетических потенций от дифференцировки клеток экспланта. У пшеницы наилучшие результаты получены при использовании 14–17-дневных незрелых зародышей [1, 6, 7, 16]. Кроме того, реализацию морфогенетических возможностей клеточных культур определяют условия выращивания in vitro [8–10, 12, 15, 17, 22].
Из вышеизложенного следует, что одним из направлений исследовательской активности остается поиск факторов, обеспечивающих формирование морфогенного каллуса и последующую успешную регенерацию из него растений.
Если для культуры пыльников злаков однозначно установлено, что условия роста донорных растений существенно влияют на способность к андрогенезу, причем максимальный выход эмбриогенных структур наблюдается при выращивании исходного материала в полевую вегетацию [4, 20], то в отношении культуры незрелых зародышей нет однозначного мнения. Так, например, С. Меддок и соавт. не обнаружили различий между каллусогенной способностью незрелых зародышей пшеницы, сформировавшихся в разных условиях [21]. Другие исследователи показали незначительное различие эм-бриогенного потенциала каллусных культур в зависимости от физиологического состоя- ния исходных растений [18]. Экспланты одних генотипов демонстрировали сходную реакцию на культивирование in vitro, несмотря на различающиеся условия выращивания родительских форм, в то время как другие сорта продуцировали больше эмбриогенного каллуса при пассировании материала, выращенного в теплице. Таким образом, приведенные сведения не дают четкого представления о влиянии условий произрастания исходных сортов злаков на изменчивость морфогенетических процессов в культуре незрелых зародышей.
Цель исследования. Изучить возможности морфогенетического потенциала каллус-ных культур сортов T. aestivum в зависимости от условий роста и развития растений – доноров эксплантов.
Материалы и методы. Материалом для исследования служили 12 сортов яровой мягкой пшеницы различного эколого-географического происхождения: Алтайская 81 , Алтайская 50 , Алтайская 88 , Целинная 20 , Скала , Зарница , Жница , Тулунская 10 , Ботаническая 2 , Спектр , Россиянка (Россия), Leones (Аргентина). Растения-доноры выращивали в селекционных теплицах арочного типа и в полевых условиях.
Тепличная вегетация продолжалась с января по май. Среднемесячная температура варьировала в пределах 21,0–24,9 °С. На протяжении февраля, марта, апреля и мая освещенность постепенно возрастала, равняясь 14,5; 17,8; 19,1 и 22,0 клк соответственно. Досвечивание в теплице производилось лампами ДМЗ-3000, что повышало уровень естественной освещенности на 10 клк.
Освещенность в полевых условиях в ясные солнечные дни превышала 50 клк, в пасмурную погоду составляла 10–14 клк. Среднемесячная температура в мае, июне, июле и августе была равна 12,7; 19,0; 21,5 и 16,5 °С соответственно.
Для индукции каллуса использовали незрелые зародыши размером 1,3–1,5 мм, которые пассировали на среду Линсмайера–Скуга (RM-64) [19], содержащую 0,8 % агара, 3,0 % сахарозы, 2 мг/л 2,4-Д. Клеточные культуры выращивали в темноте при температуре 26±1 °С, пересаживая каждые 30–35 дней на дифференцирующую среду с гормональным составом: 0,5 мг/л 2,4-Д и 0,5 мг/л кинетина. Выявленные зоны морфогенеза переносили на среду для регенерации, содержащую 0,2 мг/л индолилуксусной кислоты. Проростки, достигшие 5–7 см, высаживали в сосуды с почвой и доращивали до созревания в климатической камере при температуре 12 °С ночью и 17 °С днем с 16-часовым фотопериодом. Эксперимент выполнен в 4 повторениях по 60 зародышей на генотип. В ходе исследования определяли следующие показатели: число каллусов на эксплант (каллусогенез), число морфогенных клеточных линий на каллус (морфогенез), число регенерантов на морфогенный каллус (частота регенерации).
Статистический анализ данных проводили с использованием пакета прикладных программ ВИУА.
Результаты и обсуждение. Результаты тестирования сортов T. aestivum показали, что в полевую вегетацию средняя частота индукции клеточных линий (каллусогенез) составила 91,7 %, варьируя от 76,6 ( Жница ) до 100,0 % ( Целинная 20 , Алтайская 88 , Россиянка ) (табл. 1).
При выращивании донорных растений в условиях зимне-весенней вегетации в теплице средний результат оказался несколько ниже – 87,6 %. Пределы изменчивости составили от 72,9 ( Ботаническая 2 ) до 97,6 % ( Скала ). Однако реакция сортов на условия выращивания неоднозначна. Для одних ( Скала , Спектр , Жница , Зарница ) лучшими являются условия закрытого грунта, в то время как другие индуцируют клеточные линии значительно эффективнее от донорных сортов полевой вегетации.
Дисперсионный анализ полученных данных показал, что изменчивость каллусогенеза статистически значимо определяется разнообразием генотипов, условий произрастания растений-доноров и их взаимодействием (табл. 2).
Морфогенез в среднем по всем сортам эффективнее реализовался у эксплантов мягкой пшеницы летнего периода развития, достигнув 65,7 %. Размах изменчивости по признаку составил от 28,4 (Тулунская 10) до 91,2 % (Leones). Средняя частота формирова- ния морфогенных каллусов в тепличную вегетацию оказалась несколько ниже – 55,7 %, варьируя от 43,8 (Ботаническая 2) до 76,7 % (Скала). Условия выращивания исходных растений в теплице снижали морфогенетические возможности каллусных тканей большинства генотипов, за исключением сортов Тулунская 10, Зарница и Алтайская 88.
Таблица 1
Реакция сортов яровой мягкой пшеницы, выращенных в различных условиях, на культивирование in vitro
|
Сорт |
Каллусогенез, % |
Морфогенез, % |
Частота регенерации растений, % |
|||
|
1 |
2 |
1 |
2 |
1 |
2 |
|
|
Скала |
81,8 |
97,6 |
88,9 |
76,7 |
356,3 |
275,8 |
|
Спектр |
84,8 |
96,8 |
78,6 |
53,3 |
402,3 |
140,0 |
|
Ботаническая 2 |
97,6 |
72,9 |
73,7 |
43,8 |
94,3 |
115,4 |
|
Leones |
95,3 |
81,8 |
91,2 |
52,2 |
146,2 |
49,4 |
|
Тулунская 10 |
92,7 |
76,3 |
28,4 |
53,6 |
157,1 |
155,6 |
|
Целинная 20 |
100,0 |
88,0 |
75,2 |
46,6 |
23,5 |
17,1 |
|
Жница |
76,6 |
81,6 |
63,9 |
45,0 |
117,4 |
83,3 |
|
Зарница |
86,0 |
95,3 |
43,2 |
68,3 |
29,7 |
46,4 |
|
Алтайская 81 |
98,4 |
88,8 |
52,6 |
50,6 |
6,6 |
22,7 |
|
Алтайская 50 |
86,6 |
86,4 |
69,0 |
53,7 |
3,6 |
30,8 |
|
Алтайская 88 |
100,0 |
94,5 |
56,1 |
60,5 |
3,3 |
23,1 |
|
Россиянка |
100,0 |
91,0 |
67,9 |
64,0 |
1,8 |
25,0 |
|
Среднее значение |
91,7 |
87,6 |
65,7 |
55,7 |
111,8 |
81,3 |
Примечание. 1 – полевая вегетация; 2 – зимне-весенняя вегетация в теплице.
Данные дисперсионного анализа подтверждают достоверность наблюдаемых различий по признаку «морфогенез» как по ге- нотипам, так и по условиям роста и развития исходных сортов, а также по их взаимодействию (табл. 2).
Таблица 2
Влияние генотипов и условий произрастания исходного материала на процессы культивирования незрелых зародышей яровой мягкой пшеницы
|
Источник вариации |
F ф |
||
|
Каллусогенез |
Морфогенез |
Регенерация растений |
|
|
Генотипы |
4,5** |
6,7* |
50,8* |
|
Условия |
5,3** |
20,6* |
6,6** |
|
Взаимодействие «генотип – условия» |
3,1* |
5,8* |
6,0* |
Примечание. F ф – фактическое значение F-критерия по фактору; *, ** – достоверно при 1 и 5 % уровне значимости соответственно.
Регенерационные процессы в среднем по генотипам активнее протекали в клеточных культурах, полученных из эксплантов летней вегетации. Частота регенерации оказалась равной 111,8 %, в условиях теплицы она составила 81,3 % (табл. 1). Однако реакция изученных образцов была неоднозначной и определялась физиологическим состоянием донорных растений. Условия открытого грунта оказали благоприятное воздействие на реализацию регенерационного потенциала одних сортов (Скала, Leones, Спектр, Целинная 20, Жница) и снижали выход растений-регенерантов у остальных генотипов. Следует отметить, что сорта с низкой регенерационной активностью значительно повышали ее при использовании условий теплицы для выращивания доноров эксплантов. В результате частота выхода регенерантов у сортов Алтайская 81, Алтайская 50, Алтайская 88 и Россиянка возросла в 3,4; 8,6; 7,0 и 13,9 раза соответственно. Указанные сорта обнаружили низкий выход регенерантов (от эксплантов, выращиваемых в поле), несмотря на относи- тельно высокую долю морфогенных каллусов (52,6–69,0 %). Это объясняется тем, что большинство морфогенных клеточных линий развивались по пути ризогенеза, что исключило формирование in vitro полноценных растений. Следовательно, условия развития зародышей (будущих эксплантов) ex vivo в зимне-весеннюю вегетацию в теплице способствовали в дальнейшем индукции эмбриоидогенеза и гем-моризогенеза после введения их в культуру. Это привело к столь существенному изменению направления морфогенеза и многократно повысило выход растений-регенерантов.
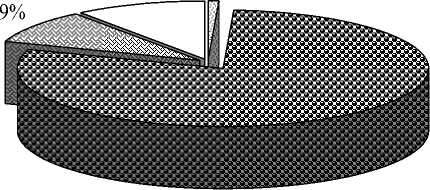
Результаты дисперсионного анализа подтвердили статистически значимое влияние на признак «регенерация растений» всех источников вариации (табл. 2). Сравнивая доли вклада генотипического разнообразия и условий в образовательные процессы при культивировании незрелых зародышей пшеницы, мы видим, что каллусо-, морфогенез и регенерация растений определяются в большей степени генотипом и взаимодействием «генотип – условия» (рис. 1).
4%
31%
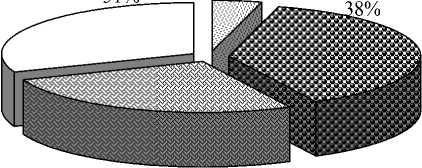
27%
Каллусогенез
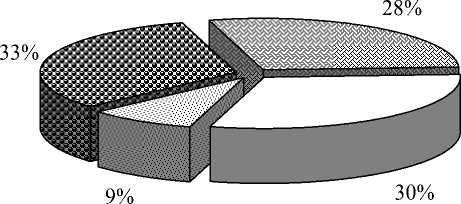
Морфогенез
-
□ Условия
^ Генотип
0 Взаимодействие генотип х условия
-
□ Случайное
Регенерация растений
Рис. 1. Вклад источников вариации в общую изменчивость образовательных процессов в культуре незрелых зародышей яровой мягкой пшеницы
Анализ данных по влиянию условий выращивания растений – доноров эксплантов свидетельствует о постоянном, но низком вкладе этого фактора в изменчивость различных культуральных процессов. Количественная оценка влияния данного фактора в зависимости от стадии развития культуры варьировала от 1 до 9 %. Тем не менее полевые условия способствовали увеличению средних значений всех изучаемых процессов в культуре незрелых зародышей: каллусогенез повысился на 4,1 %, морфогенез – на 10,0 % и регенерация растений – на 30,5 %.
Следует также принять во внимание, что генотип-средовое взаимодействие существенно для всех изученных признаков. Его вклад в наблюдаемую изменчивость каллусо-, морфогенеза и регенерацию растений составила 27, 28 и 9 % соответственно. Различные генотипы, подвергнутые одинаковому изменению условий среды, реагировали на него по-разному.
Заключение. Таким образом, различные образовательные процессы в культуре незрелых зародышей пшеницы в большей мере зависят от наследственности сортов и лишь в некоторой степени – от комплекса факторов среды, в которых эти сорта произрастают. Реакция отдельных генотипов на условия выращивания доноров эксплантов специфична. Полевые условия наиболее благоприятны для генотипов с высоким уровнем регенерации, условия зимнее-весенней вегетации в теплице – для сортов с низким регенерационным потенциалом.
-
1. Батыгина Т. Б. Прикладные аспекты эмбриологии. Автономность зародыша и эмбриокультура цветковых растений / Т. Б. Батыгина, В. Е. Васильева // Ботанический журн. – 1987. – Т. 72, № 2. – С. 155–161.
-
2. Гапоненко А. К. Регенерация растений различных генотипов пшеницы Тriticum aestivum L . in vitro / А. К. Гапоненко, М. А. Мунтян // Доклады АН СССР. – 1984. – Т. 278, № 5. – С. 12–31.
-
3. Григорьева Л. П. Скрининг сортов пшеницы по способности к морфогенезу в культуре незрелых зародышей in vitro / Л. П. Григорьева, И. А. Шлецер // Известия Алтайского гос. ун-та. – 2006. – № 3 (51). – С. 64–66.
-
4. Дьячук Т. В. Культура пыльников пшеницы и ее использование в селекции / Т. В. Дьячук, П. А. Дьячук // Биология культивируемых клеток и биотехнология : Международная конференция. – Новосибирск, 1988. – С. 207–208.
-
5. Ежова Т. А. Генетический контроль тотипотентности растительных клеток в культуре in vitro / Т. А. Ежова // Онтогенез. – 2003. – Т. 34, № 4. – С. 245–252.
-
6. Круглова Н. Н. Незрелый зародыш пшеницы как морфогенетический компетентный эксплант / Н. Н. Круглова, А. А. Катасонова // Физиология и биохимия растений. – 2009. – Т. 41, № 2. – Р. 124–131.
-
7. Круглова Н. Н. Периодизация эмбриогенеза пшеницы как методологический аспект биотехнологических разработок / Н. Н. Круглова // Известия Уфимского научного центра РАН. – 2012. – № 2. – С. 21–24.
-
8. Никитина Е. Д. Влияние температуры и освещения на прямое прорастание незрелых зародышей Triticum aestivum L. в культуре in vitro / Е. Д. Никитина, Л. П. Хлебова // Известия Алтайского гос. ун-та. – 2014. – № 3–1 (83). – С. 46–50.
-
9. Никитина Е. Д. Влияние фитогормонов на морфогенез и регенерацию растений каллусных тканей Triticum aestivum / Е. Д. Никитина // Генетика. – 1994. – № 30 (прил.). – С. 109–110.
-
10. Никитина Е. Д. Разработка отдельных элементов технологии клеточной селекции яровой пшеницы на устойчивость к абиотическим стрессам / Е. Д. Никитина, Л. П. Хлебова, О. В. Ере-щенко // Известия Алтайского гос. ун-та. – 2014. – Т. 2, № 3 (79). – С. 50–54.
-
11. Никитина Е. Д. Роль генотипа в реализации морфогенетических процессов в культуре незрелых зародышей у Triticum aestivum L. / Е. Д. Никитина // Сибирский вестн. сельскохозяйственной науки. – 2004. – № 2 (152). – С. 32–34.
-
12. Никитина Е. Д. Роль физических факторов в реализации морфогенетического потенциала клеточных культур T. aestivum / Е. Д. Никитина // Новые методы биотехнологии растений : материалы II Российского симпозиума. – Пущино : Пущинский НЦ, 1993. – С. 159.
-
13. Никитина Е. Д. Создание источников устойчивости яровой пшеницы к воздействию никеля методами клеточной селекции in vitro / Е. Д. Никитина, Л. П. Хлебова, Г. Г. Соколова // Известия Алтайского гос. ун-та. – 2013. – № 3–1 (79). – С. 088–090.
-
14. Создание стрессоустойчивого материала яровой мягкой пшеницы с использованием клеточной селекции in vitro / Е. Д. Никитина [и др.] // Известия Алтайского гос. ун-та. – 2013. – № 3–2 (79). – С. 95–98.
-
15. Суханов Н. П. Условия получения каллуса и регенерантов в культуре незрелых зародышей пшеницы / Н. П. Суханов, Н. Д. Папазян // Апо-
миксис и цитоэмбриология растений. – 1983. – № 5. – С. 124–128.
-
16. Хамула П. В. Влияние генотипа и размера зародыша мягкой пшеницы на частоту каллусооб-разования / П. В. Хамула, В. Д. Солодовниченко, Л. В. Базько // Селекционно-генетические аспекты повышения продуктивности зерновых культур. – Мироновка, 1987. – С. 45– 48.
-
17. Effect of 2,4-dichlorophenoxyacetic acid and NaCl on the establishment of callus and plant regeneration in durum and bread wheat / A. Pellegrineschi [et al.] // Plant Cell, Tissue, Organ Culture. – 2004. – Vol. 77, № 3. – Р. 245–250.
-
18. Identification of callus types for long-term maintenance and regeneration from commercial cultivars of wheat ( Triticum aestivum L.) / F. E. Redway [et al.] // TAG. – 1990. – № 79. – P. 609–617.
-
19. Linsmaier E. Organic growth factor reqular-ments of tobacco tissue culture / E. Linsmaier, F. Sko-og // Physiol. Plant. – 1965. – Vol. 18, № 1. – P. 100–127.
-
20. Ouyang J. W. Induction of pollen plants in Triticum aestivum / J. W. Ouyang // Haploids of higher plants in vitro . – Springer-Verlag ; Berlin ; Heidelberg ; New York, 1986. – P. 26–41.
-
21. Plant regeneration from cultured immature embryos and inflorescences of 25 cultivars of wheat ( T. aestivum ) / S. E. Maddock [et al.] // J. Exp. Bot. – 1983. – Vol. 34, № 144. – P. 915–926.
-
22. Przetakiewicz A. The effect of auxin on plant regeneration of wheat, barley and Triticale / A. Prze-takiewicz, W. Orczyk, A. Nadolska-Orczyk // Plant Cell, Tissue and Organ Culture. – 2003. – Vol. 73, № 3. – Р. 245–256.
MORPHOGENESIS FEATURES OF SPRING WHEAT IN VITRO CULTURE DEPENDING ON GROWING CONDITIONS
E.D. Nikitina 1 , L.P Khlebova2
-
1Altai Research Institute of Agriculture, Barnaul, 2Altai State University, Barnaul
Список литературы Особенности морфогенеза яровой мягкой пшеницы в культуре in vitro в зависимости от условий произрастания
- Батыгина Т. Б. Прикладные аспекты эмбриологии. Автономность зародыша и эмбриокультура цветковых растений/Т. Б. Батыгина, В. Е. Васильева//Ботанический журн. -1987. -Т. 72, № 2. -С. 155-161.
- Гапоненко А. К. Регенерация растений различных генотипов пшеницы Тriticum aestivum L. in vitro/А. К. Гапоненко, М. А. Мунтян//Доклады АН СССР. -1984. -Т. 278, № 5. -С. 12-31.
- Григорьева Л. П. Скрининг сортов пшеницы по способности к морфогенезу в культуре незрелых зародышей in vitro/Л. П. Григорьева, И. А. Шлецер//Известия Алтайского гос. ун-та. -2006. -№ 3 (51). -С. 64-66.
- Дьячук Т. В. Культура пыльников пшеницы и ее использование в селекции/Т. В. Дьячук, П. А. Дьячук//Биология культивируемых клеток и биотехнология: Международная конференция. -Новосибирск, 1988. -С. 207-208.
- Ежова Т. А. Генетический контроль тотипотентности растительных клеток в культуре in vitro/Т. А. Ежова//Онтогенез. -2003. -Т. 34, № 4. -С. 245-252.
- Круглова Н. Н. Незрелый зародыш пшеницы как морфогенетический компетентный эксплант/Н. Н. Круглова, А. А. Катасонова//Физиология и биохимия растений. -2009. -Т. 41, № 2. -Р. 124-131.
- Круглова Н. Н. Периодизация эмбриогенеза пшеницы как методологический аспект биотехнологических разработок/Н. Н. Круглова//Известия Уфимского научного центра РАН. -2012. -№ 2. -С. 21-24.
- Никитина Е. Д. Влияние температуры и освещения на прямое прорастание незрелых зародышей Triticum aestivum L. в культуре in vitro/Е. Д. Никитина, Л. П. Хлебова//Известия Алтайского гос. ун-та. -2014. -№ 3-1 (83). -С. 46-50.
- Никитина Е. Д. Влияние фитогормонов на морфогенез и регенерацию растений каллусных тканей Triticum aestivum/Е. Д. Никитина//Генетика. -1994. -№ 30 (прил.). -С. 109-110.
- Никитина Е. Д. Разработка отдельных элементов технологии клеточной селекции яровой пшеницы на устойчивость к абиотическим стрессам/Е. Д. Никитина, Л. П. Хлебова, О. В. Ерещенко//Известия Алтайского гос. ун-та. -2014. -Т. 2, № 3 (79). -С. 50-54.
- Никитина Е. Д. Роль генотипа в реализации морфогенетических процессов в культуре незрелых зародышей у Triticum aestivum L./Е. Д. Никитина//Сибирский вестн. сельскохозяйственной науки. -2004. -№ 2 (152). -С. 32-34.
- Никитина Е. Д. Роль физических факторов в реализации морфогенетического потенциала клеточных культур T. aestivum/Е. Д. Никитина//Новые методы биотехнологии растений: материалы II Российского симпозиума. -Пущино: Пущинский НЦ, 1993. -С. 159.
- Никитина Е. Д. Создание источников устойчивости яровой пшеницы к воздействию никеля методами клеточной селекции in vitro/Е. Д. Никитина, Л. П. Хлебова, Г. Г. Соколова//Известия Алтайского гос. ун-та. -2013. -№ 3-1 (79). -С. 088-090.
- Создание стрессоустойчивого материала яровой мягкой пшеницы с использованием клеточной селекции in vitro/Е. Д. Никитина //Известия Алтайского гос. ун-та. -2013. -№ 3-2 (79). -С. 95-98.
- Суханов Н. П. Условия получения каллуса и регенерантов в культуре незрелых зародышей пшеницы/Н. П. Суханов, Н. Д. Папазян//Апомиксис и цитоэмбриология растений. -1983. -№ 5. -С. 124-128.
- Хамула П. В. Влияние генотипа и размера зародыша мягкой пшеницы на частоту каллусообразования/П. В. Хамула, В. Д. Солодовниченко, Л. В. Базько//Селекционно-генетические аспекты повышения продуктивности зерновых культур. -Мироновка, 1987. -С. 45-48.
- Effect of 2,4-dichlorophenoxyacetic acid and NaCl on the establishment of callus and plant regeneration in durum and bread wheat/A. Pellegrineschi //Plant Cell, Tissue, Organ Culture. -2004. -Vol. 77, № 3. -Р. 245-250.
- Identification of callus types for long-term maintenance and regeneration from commercial cultivars of wheat (Triticum aestivum L.)/F. E. Redway //TAG. -1990. -№ 79. -P. 609-617.
- Linsmaier E. Organic growth factor reqularments of tobacco tissue culture/E. Linsmaier, F. Skoog//Physiol. Plant. -1965. -Vol. 18, № 1. -P. 100-127.
- Ouyang J. W. Induction of pollen plants in Triticum aestivum/J. W. Ouyang//Haploids of higher plants in vitro. -Springer-Verlag; Berlin; Heidelberg; New York, 1986. -P. 26-41.
- Plant regeneration from cultured immature embryos and inflorescences of 25 cultivars of wheat (T. aestivum)/S. E. Maddock //J. Exp. Bot. -1983. -Vol. 34, № 144. -P. 915-926.
- Przetakiewicz A. The effect of auxin on plant regeneration of wheat, barley and Triticale/A. Przetakiewicz, W. Orczyk, A. Nadolska-Orczyk//Plant Cell, Tissue and Organ Culture. -2003. -Vol. 73, № 3. -Р. 245-256.
