Особенности развития Dracocephalum fragile Turcz. ex Benth. (Lamiaceae) в Туве
 в Туве")
Автор: Денисова Гульнора Робеховна, Черемушкина Вера Алексеевна, Асташенков Алексей Юрьевич, Гусева Александра Алексеевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 4, 2017 года.
Бесплатный доступ
Изучены закономерности развития травянисто-длиннозернистого травянистого поликарпа Dracocephalum fragile Turcz. бывшие жители Бентама из Тувы. Основу системы отростков у взрослых составляет полициклический длиннозернисто-среднезернистый монокарпический отросток. Описана форма жизни и особенности формирования отростков в онтогенезе. Показаны сходства и различия онтоморфогенеза особей, растущих на гравии и осыпях. Онтогенез особей D. fragile является полным и сложным, состоящим из онтогенеза семенной особи и сниженных онтогенезов аффилированных особей. Онтогенез семенных особей происходит в последовательности следующих фаз морфогенеза: первичный побег (p, j) - первичный куст (im) - ложе (v, g1, g2, g3) - частица кущения (ss, s). Морфогенез особей вегетативного происхождения характеризуется следующими фазами: частичный побег (im) - частичные побеги и кустарники (v, g1, g2, g3, ss) - частичный побег (и). Ход онтоморфогенеза семян и вегетативных особей, растущих на каменистой осыпи, сходен с онтогенезом особей, растущих на гравии. Отмечено местообитание-зависимое морфологическое и динамическое поливариантность развития D. хрупких особей.
Короткий адрес: https://sciup.org/146279526
IDR: 146279526 | УДК: 581.522.5+582.929.4
Developmental patterns of Dracocephalum fragile Turcz. ex Benth. (Lamiaceae) in Tuva
We studied the developmental patterns of longrhizomatous-taproot herbaceous polycarpic of Dracocephalum fragile Turcz. ex Bentham individuals from Tuva. The base of the shoot system of adults is made up of polycyclic longrhizomatous-middlerosulate monocarpic shoot. A life form and features of shoot formation in ontogeny are described. Similarities and differences of ontomorphogenesis of individuals growing on the gravel and scree are shown. Ontogenesis of individuals of D. fragile is full and complex, consistig of ontogenesis of a seed individual and the reduced ontogeneses of affiliated individuals. Ontogenesis of seed individuals takes place in the sequence of the following phases of a morphogenesis: primary shoot (p,j) - primary bush (im) - bed (v,g1,g2,g3) - tillering particle (ss, s). The morphogenesis of individuals of a vegetative origin is characterized by the following phases: partial shoot (im) - partial shoots and bushes (v,g1,g2,g3, ss) - partial shoot (s). The course of an ontomorphogenesis of the seed and vegetative individuals growing on the scree is similar to ontogenesis of the individuals growing on the gravel. Habitat-dependent morphological and dynamic polyvariety of development of D. fragile individuals is pointed out.
Текст научной статьи Особенности развития Dracocephalum fragile Turcz. ex Benth. (Lamiaceae) в Туве
Введение. Согласно оценкам влияния глобальных климатических изменений на экосистемы, в будущем предполагается сокращение площади местообитаний многих видов растений. Так, в горах Алтае-Саянского экорегиона, по различным сценариям, к концу XXI века сократится площадь высокогорий на 10-23 % (Чебакова и др., 2011). В связи с чем, сохранение разнообразия видов является одной из главных задач современной биологии. Особое внимание в этом вопросе заслуживают редкие, эндемичные виды и виды, произрастающие на краю ареала. Одно из таких растений - Dracocephalum fragile Turcz. ex Benth. - змееголовник ломкий, который произрастает в Восточной Сибири и в Северной Монголии (Пешкова, 1997; Urgamal et al., 2014). На территории Сибири он встречается в Туве на нагорье Сангилен и хребте Хорумну-Тайга и в Бурятии на Восточном Саяне. Сообщества с участием D. fragile также распространены в сопредельных районах Монголии (Urgamal et al., 2014). Ареал вида - сангиленско-восточносаянско-северомонгольский, где. D. fragile является эндемиком (Малышев, 1965; Urgamal et al, 2014).
В последнее время для разработки программ по сохранению видов используют данные о побегообразовании, сезонном развитии и жизненной форме растений, что позволяет определить новые тенденции их сохранения (Савиных, 2013).
Развитие особей D. fragile в природе ранее не изучалось. В связи с этим, цель нашей работы - изучение строения взрослых особей и онтоморфогенеза D. fragile на северо-западной границе ареала в Туве.
Методика. Онтоморфогенез особей D. fragile изучен в Туве на нагорье Сангилен в долине р. Балытыг-Хем на высоте 2081 над ур. м.: в группировке на галечнике, закустаренном Pentaphylloides fruticosa (L.) О. Schwarz и Salix saxatilis Turcz. ex Ledeb., где из трав преобладали D. fragile, Elytrigia gmelinii (Trin.) Nevski, Poa alpina L., Sedum hybridum L., и в разнотравно-овсяницевом сообществе на осыпи, где преимущественно произрастали D. fragile, Festuca kryloviana Reverd, Carex duriuscula C.A. Mey, Oxytropis tragacanthoides Fisch., Artemisia santolinifolia Turcz. ex Bess.
Жизненная форма D. fragile описана по экологоморфологической классификации жизненных форм И.Г. Серебрякова (1962).
При изучении онтогенеза D. fragile опирались на концепцию дискретного описания онтогенеза (Работнов, 1950; Уранов, 1975). Выделение фаз морфогенеза проводили в соответствии с представлениями о нём (Смирнова и др., 1976; Серебрякова, 1980; Савиных, Черемушкина, 2015). Описание структуры взрослых особей и побегов основано на подходах И.Г. Серебрякова (1959, 1962). Также использованы представления о структурно-биологических единицах Л.Е. Гатцук (1994; 2008), Е.Л. Нухимовского (1997), Ю.А. Боброва (2009) и Е.Б. Колеговой, В. А. Черемушкиной (2012). Модель побегообразования дана согласно классификации Н.П. Савиных (2012).
Результаты и обсуждение. По данным М.Ю. Телятникова (2016), в восточной части Восточного Саяна D. fragile произрастает в двух ассоциациях семигумидно-семиаридного биоклиматического сектора. В ассоциации Tralictro alpine-Betuletum humillis особи D. fragile встречаются в 21-40 % описаний. Эти сообщества характерны для нижней части высокогорного пояса и приурочены к склонам речных террас и бортам долин рек преимущественно южной экспозиции на высоте 1990-2020 м над ур.м. В ассоциации Pulsatillo ambiguae-Caricetum pediformis особи D. fragile отмечены редко (не более 19% описаний). Ц,енозы приурочены к пологим средним и нижним частям склонов гор, на высоте 1650-2100 м над ур. м. По данным И.М. Гаджиева с соавт. (2002), D. fragile в Монголии встречается в ассоциациях Dracocephalo fragilis - Elymetum sibirici.
D. fragile - петрофит. Цветет с конца июня по первую половину августа (Malyshev, 1965).
Изучение его взрослых особей показало, что D. fragile - длиннокорневищно -стержнекорневой поликарпик.
Основу побеговой системы взрослых особей D. fragile составляют разветвленные полициклические длиннокорневищносреднерозеточные монокарпические побеги (рис.1).
При полном цикле развития полициклические длиннокорневищно-среднерозеточные побеги состоят из трех-пяти годичных приростов. Геофильные участки этих побегов длиной 8-40 см нарастают моноподиально под землей от года до трех лет и становятся коммуникационным корневищем. Каждый годичный прирост корневища состоит из 2-3 укороченных и 7-11 удлиненных метамеров. Придаточные корни образуются в узлах и междоузлиях почти всех метамеров геофильной части побега. Закрытая верхушечная почка растущего корневища на 2-4 год развертывается в годичный надземный верхнерозеточный побег, состоящий из 3 коротких метамеров с чешуевидными листьями и 1-2 удлиненных, 2-3 укороченных метамеров с зелеными листьями. Заканчивается ось побега открытой терминальной почкой. Осенью надземная часть побега полегает, засыпается, но не укореняется. На следующий год верхушечная почка трогается в рост, и образуется удлиненный ортотропный годичный генеративный побег. Ветвление полициклического побега мезотонное и происходит за счет раскрытия почек предпоследнего и предшествующему ему годичных приростов. Первыми трогаются в рост почки удлиненной части последнего годичного прироста корневища. Растущие длиннокорневищные побеги участвуют в разрастании особи. В год, когда главный побег формирует верхнерозеточный вегетативный годичный побег, из нижней и (или) средней пар почек его укороченной базальной части текущего годичного прироста формируются 1-2 боковых силлептических дициклических монокарпических побега. В первый год жизни, одновременно с главным побегом, они развиваются как верхнерозеточные, на следующий год зацветают. В год, когда главный побег зацветает, из верхней пары почек укороченной базальной части предпоследнего годичного прироста развертываются 1-2 моноциклических ортотропных удлиненных генеративных побега, из почек удлиненных метамеров - верхнерозеточные вегетативные побеги. После отцветания ортотропная часть годичных генеративных побегов отмирает до среднерозеточной части, которая у полициклических побегов представлена тремя, у дициклических двумя укороченными метамерами с настоящими листьями. Моноциклические побеги полностью отмирают и не участвуют в побегообразовании. Постепенно некорневищная часть побегов отмирает и в структуре особей выделяется лишь геофильные участки полициклических побегов, почки которых реализуются постепенно в течение всей жизни побега с образованием длиннокорневищных или дициклических среднерозеточных монокарпических побегов. Длиннокорневищные побеги в последующем становятся полициклическими длиннокорневищно-среднерозеточными побегами замещения.
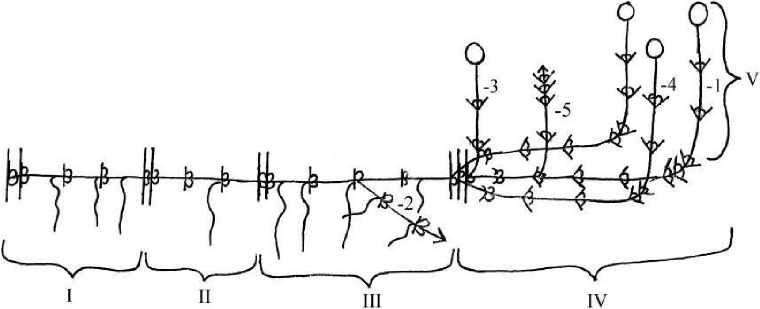
Р и с . 1 . Структура разветвленного полициклического длиннокорневищносреднерозеточного побега.
1-длиннокорневищно-среднерозеточный генеративный побег;
2- растущее корневище;
3- моноциклический удлиненный генеративный побег;
4- дициклический среднерозеточный генеративный побег;
5- верхнерозеточный вегетативный побег;
I -V - годичные приросты.
Онтоморфогенез семенных особей D. fragile на галечнике
Прорастание семян надземное. Проростки появляются весной или осенью и имеют две яйцевидные с цельным краем, клиновидные у основания семядоли и 1-2 пары черешковых настоящих продолговатояйцевидных листьев (рис. 2). В пазухах семядольных и первой пары настоящих листьев закладываются почки. У всех растений гипокотиль хорошо выражен - от 0,5 до 0,6 см, эпикотиль -0,1 см. Главный корень 2,2-2,5 см в длину.
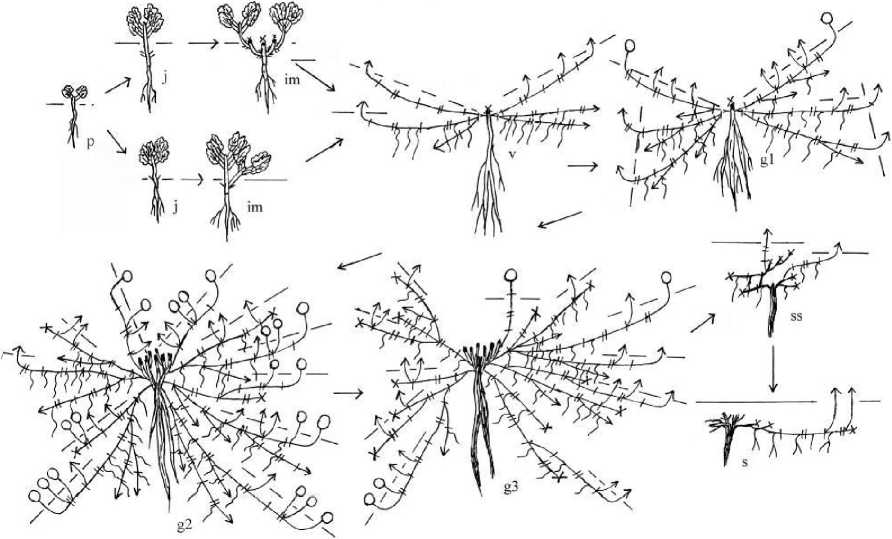
Рис. 2 . Онтоморфогенез семенных особей D. fragile на галечнике Онтогенетические состояния: j - ювенильное; im- имматурное; v -виргинильное; gl - молодое генеративное; g2 - зрелое генеративное; g3-старое генеративное; ss - субсенильное; s - сенильное. (Условные т ?
обозначения: • - вегетативный верхнерозеточный побег; ' - генеративный
побег;

- длиннокорневищный побег; - отмерший побег;
годичный прирост;
- сближенное междоузлие;
- удлиненное
междоузлие;
- уровень почвы;
- придаточный корень;
- узел с чешуевидными листьями;
- узел с листьями
переходного типа;
- узел с настоящими листьями.
При весеннем прорастании семян на галечнике ювенильное состояние наступает в этот же год. Особи нарастают моноподиально. Первый метамер вытягивается и розеточная часть побега выносится над поверхностью почвы. Первичный побег становится верхнерозеточным высотой до 3,1 см. При осеннем прорастании семян ювенильные особи зимуют в виде розеточного побега. Весной следующего года у таких растений третий или четвертый метамер вытягивается, и главный побег становится розеточно-верхнерозеточным. За счет контрактильной деятельности главного корня эпикотиль и гипокотиль побега ювенильных особей втягиваются в субстрат. Главный корень ветвится, образуются боковые корни I II порядка.
В имматурном состоянии у всех особей формируется первичный куст. У большинства растений за счет полегания главного побега и его засыпания верхушечная почка не погибает. Трогаются в рост боковые почки в пазухах семядольных и первой пары настоящих листьев. Первичный куст образован главным и 1-4 боковыми верхнерозеточными побегами 11 порядка, которые состоят из двух укороченных метамеров с чешуевидными листьями, одного, реже двух, удлиненных и 2-4 укороченных метамеров с настоящими листьями. К осени надземная часть главного побега отмирает, боковые побеги полегают, но не укореняются. Корневая система состоит из главного и боковых корней 111-IV порядка. Длительность состояния один год.
У некоторых особей главный побег отмирает в начале вегетации до удлиненного метамера. Трогаются в рост две почки, образуются верхнерозеточные побеги 11 порядка. После вегетации побеги часто отмирают до первого метамера их удлиненной части, их базальные части уже втянуты в почву. Почками возобновления в таком случае становятся любые почки сохранившегося участка побега. Подземная часть первичного побега сильно утолщается. Такие особи в имматурном состоянии могут находиться до двух лет.
В виргинильном состоянии формируется куртина. На начальных этапах куртина состоит из первичного куста и 2-4 полициклических длиннокорневищно-верхнерозеточных побегов 111 порядка. Первичный куст образован 2-4 ди-трициклическими среднерозеточными вегетативными побегами 11 порядка. Все побеги нарастают моноподиально. Первые годичные приросты побегов 11 порядка располагаются плагиотропно, ортотропной остается прирост текущего года. Они ветвятся, образуя побеги 111 порядка. Из верхней пары почек базальной и удлиненной частей второго годичного прироста развертываются 1-3 дициклических среднерозеточных вегетативных побега с длинночерешковыми ланцетовидными листьями длиной 1,52,5 см, шириной 0,4-0,8 см. Полициклические длиннокорневищноверхнерозеточные побеги развиваются из нижних пар почек базальной части первого годичного прироста побегов 11 порядка. Первый год или два, они нарастают под землей, образуя гипогеогенное корневище. За год у таких побегов формируется 7-9 междоузлий, первые 2-3 укороченные, остальные 5-6 удлиненные. На второй или третий год побег развивается как надземный годичный верхнерозеточный. Длина плагиотропной части длинокорневищно-верхнерозеточных побегов может достигать 8 см. Они могут ветвиться. Из нижней пары почек базальной части первого и второго годичных приростов развиваются новые длиннокорневищно-верхнерозеточные побеги IV порядка. Из верхней пары почек базальной и удлиненной частей второго годичного прироста развертываются анизотропные дициклические среднерозеточные вегетативные побеги. Корневая система смешанная. Большое число придаточных корней образуется в области укороченных междоузлий гипогеогенного корневища. Придаточные корни развиваются как в узлах, так на междоузлиях.
На 5-6 год развития семенные особи зацветают, и растения переходят в молодое генеративное состояние. К цветению переходят побеги первичного куста, зацветает 2-3 среднерозеточных побега II порядка. В средней части годичных ортотропных генеративных побегов развивается 2-3 вегетативных розеточных побега обогащения. После цветения их ортотропная часть отмирает. Соцветие - колосовидный тирс, состоящий из 4-6 дихазиев. Длина соцветия не превышает 8 см. Увеличение числа длиннокорневищно-среднерозеточных побегов до 57 и длины их геофильных участков до 9,5 см приводит к разрастанию куртины, которая в диаметре может достигать 15-20 см. Длиннокорнсвищныс побеги продолжают нарастать моноподиально. За счет интенсивного ветвления всех типов побегов увеличивается число растущих корневищ до 10 на особь. Корневая система смешанная. В этом состоянии растения находятся 2-3 года.
Куртина зрелых генеративных растений сильно разрастается и может достигать в диаметре 75-100 см. Она состоит из слабо кустящегося первичного куста и 7-46 разветвленных полициклических длиннокорневищно- среднерозеточных побегов п, и+1, и+2 порядков, структура которых описана выше. Основу побеговой системы первичного куста составляют 1-3 дициклических среднерозеточных побега IV-порядка, которые сформировались в результате ветвления дициклических среднерозеточных побегов 111 порядка. В первый год жизни они развиваются как верхнерозеточные, осенью полегают, погружаются в субстрат, но не укореняется. На следующий год их верхушечная почка трогается в рост, и образуется удлиненный ортотропный годичный генеративный побег. Ветвление дициклического среднерозеточного побега начинается на второй год его развития: трогаются в рост почки удлиненной части первого годичного прироста и формируются 1-2 моноциклических удлиненных генеративных и 1-4 верхнерозеточных вегетативных побега.
Разветвленные полициклические длиннокорневищносреднерозеточные побеги зацветают. После их цветения гипогеогенное корневище начинает нарастать симподиально. Длина геофильных участков полициклических длиннокорневищно-среднерозеточных побегов от 8 до 50 см, толщина до 0,5 см. За счет развертывания большого числа почек на дициклических среднерозеточных и разветвленных полициклических длиннокорневищносреднерозеточных побегах число генеративных побегов увеличивается до 150-177. Также увеличивается длина соцветия до 12 см и число дихазиев на нем до 8. Корневая система смешанная. Хорошо развита система главного, боковых и придаточных корней. На гипогеогенных корневищах в зоне формирования растущих корневищ образуются участки скопления придаточных корней. У некоторых особей коммуникационные корневища разрушаются, начинается вегетативное размножение, но семенное остается доминирующим.
В старом генеративном состоянии в результате перегнивания коммуникационных корневищ образуются дочерние особи разных онтогенетических состояний: от имматурного до субсенильного. Вегетативное размножение становится доминирующим способом воспроизведения особей. Куртина уменьшается в диаметре. Центральная часть первичного куста разрушена с большим числом отмерших дициклических среднерозеточных побегов, в живых остается один неветвящийся дициклический среднерозеточный побег. Сокращается число коммуникационных корневищ до 4-9. Покровные ткани коммуникационных корневищ разрушаются и слущиваются. Длина корневища варьирует от 35 до 42 см, толщина до 0,6 см. Побегообразование у особей осуществляется за счет развертывания спящих почек, сохранившихся на старых коммуникационных корневищах. В куртине формируется всего 1-2 разветвленных полициклических длиннокорневищно-среднерозеточных побега. Цветут полициклические длиннокорневищно-среднерозеточные и дициклические среднерозеточные побеги. Моноциклические удлиненные генеративные побеги в старом генеративном состоянии не развиваются. Длина соцветия не превышает 3,5 см. Число дихазиев в соцветии уменьшается до 4. Растущие корневища старых генеративных особей в базальной части первого годичного прироста состоят из 4-6 укороченных метамеров с почками. Они имеют хорошо развитую придаточную корневую систему и при отделении могут существовать самостоятельно. Главный корень разрушен по центральному цилиндру и остается целым лишь в проксимальной части.
При полной партикуляции главного корня, особи переходят в субсенильное или сенильное состояние.
Кустящиеся партикулы субсенильных особей представляют собой разрушенный участок главного корня, от которого отходят 1-2 коммуникационных корневища. Корневище сформировано одним, реже двумя годичными приростами, из спящих почек которого развиваются 1-3 дициклических среднерозеточных и 1-2 длиннокорневищноверхнерозеточных вегетативных побегов. В результате повторной партикуляции главного корня образуются кустящиеся партикулы, находящиеся в том же или в сенильном онтогенетическом состояниях.
У сенильных особей из спящих почек на живых участках коммуникационного корневища образуется 1-3 ортотропных верхнерозеточных побега, которые после вегетации полностью отмирают.
Онтоморфогенез особей D. fragile вегетативного происхождения на галечнике
Особи вегетативного происхождения хорошо отличаются по сохранившимся остаткам геофильных частей побегов (рис. 3).
Имматурные особи представлены моноподиально нарастающим полициклическим длиннокорневищно-верхнерозеточным побегом, подземная сфера которого образованна 1-2 годичными приростами корневища (8-21 см) с большим числом почек и придаточных корней. Надземная часть побега может ветвиться: трогается в рост одна из пазушных почек удлиненной части и формируется розеточный побег с 3-4 парами настоящих листьев. К осени годичный верхнерозеточный побег полегает, но нс укореняется.
В момент, когда трогаются в рост почки геофильной части длиннокорневищно-верхнерозеточного побега, особи переходят в виргинильное состояние. Из почек развиваются 2-6 полициклических длиннокорневищно-верхнерозеточных побегов. Характер их ветвления не отличается от ветвления семенных растений. Геофильная часть растущих длиннокорневищно-верхнерозеточных побегов в год может увеличиться до 10 см. Прегенеративный период непродолжительный и длится не более двух лет.
В молодом генеративном состоянии особь представлена 8-10 разветвленными полициклическими длиннокорневищными побегами, большинство из которых верхнерозеточные вегетативные. Растущих корневищ 12-16 на особь. Из верхушечной почки 1-2 полициклических длиннокорневищных побегов развертывается годичный генеративный удлиненный побег, и побеги становятся среднерозеточными. К цветению также могут перейти дициклические среднерозеточные побеги.
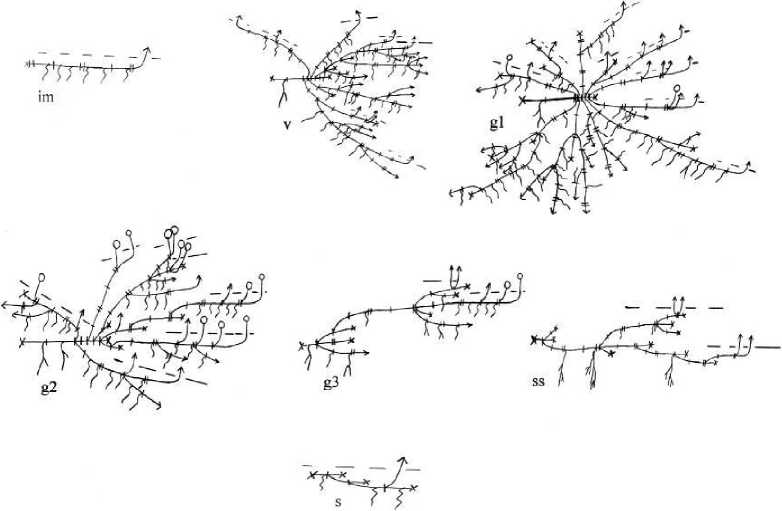
Рис. 3 . Онтоморфогенез особей D. fragile вегетативного происхождения на галечнике. Условные обозначения на рис. 2.
Зрелые генеративные особи состоят из полициклических длиннокорневищно-среднерозеточных побегов разных порядков, на гипогеогенных корневищах которых развиваются придаточные корни. У особей насчитывается от 10 до 15 цветущих побегов. К цветению переходят полициклические длиннокорневищно-среднерозеточные, дициклические среднерозеточные и моноциклические удлиненные побеги. На полициклических длиннокорневищно-среднерозеточных побегах развивается до 20 растущих корневищ на особь. В конце зрелого генеративного состояния происходит перегнивание коммуникационных корневищ.
В старом генеративном состоянии особи представлены длинным коммуникационным корневищем, из спящих почек которого развиваются 1-2 полициклических длиннокорневищносреднерозеточных генеративных побега. Моноциклические генеративные побеги в этом состоянии не образуются. Придаточные корни формируются только на молодых растущих корневищах, число которых в этом состоянии сокращается до 1-3.
Для субсенильных растений характерно отсутствие растущих корневищ. Длина сильно разрушенных коммуникационных корневищ не превышает 20 - 22 см. Дициклические среднерозеточные вегетативные побеги в числе 2-4 развиваются из почек, расположенных в укороченной части предыдущего годичного прироста дициклических среднерозеточных.
У сенильных растений коммуникационное корневище большей частью отмершее. Живым остается участок одного годичного прироста (5-6 междоузлий), из спящих почек которого развивается один-два верхнерозеточных побега с четырьмя парами листьев, которые после вегетации отмирают. Постепенно почки коммуникационного корневища реализуются и особи отмирают.
Онтоморфогенез особей D. fragile на осыпях
Семенные особи D. fragile на осыпях развиваются подобно особям, произрастающим на галечнике, у которых смена нарастания происходит в начале имматурного состояния. Однако в рост трогается не две почки, как у особей, произрастающих на галечнике, а одна из почек, расположенная в семядольном узле или в пазухе настоящих листьев: образуется розеточный побег замещения 11 порядка. За счет постоянного засыпания побега верхушечная почка не погибает, и побег может нарастать до трех лет.
В виргинильном состоянии формируется куртина, которая в конце прегенеративного периода за счет партикуляции главного корня, а также травматического повреждения корневищ распадается на парциальные образования, которые находятся в имматурном или виргинильном состояниях. В дальнейшем особи развиваются подобно парциальным образованиям особей, произрастающих на галечнике. Однако существуют и отличия. Так, у вегетативных особей в побеговой системе в связи с постоянным засыпанием на осыпях увеличивается интенсивность образования побегов. В корневой системе, начиная с имматурного состояния, преобладает в развитии один из придаточных корней, что в последующем приведет к формированию вторичностержневого придаточного корня, который уже в молодом генеративном состоянии хорошо выражен и сохраняется до конца жизни растений. С молодого генеративного состояния особи развиваются подобно семенным растениям, произрастающим на галечнике. Но из-за увеличения числа и длины метамеров у полициклических длиннокорневищно-среднерозеточных побегов в каждом онтогенетическом состоянии заметно удлиняется их геофильная часть, что способствует разрастанию особей. Также отмечено небольшое число генеративных особей, у которых формируется второй центр разрастания. Он образован в результате многократного ветвления полициклического длиннокорневищносреднерозеточного побега. Хотя центры чаще всего находятся далеко друг от друга, связь между ними на какое-то время сохраняется. В последующем они развиваются самостоятельно, обеспечивая расселение и вегетативное размножение растений.
Заключение. В условиях Тувы на северо-западной границе ареала у особей D. fragile формируется длиннокорневищно-стержнекорневая жизненная форма. Изучение особенностей развития особей показало, что они развиваются по симподиальной среднерозеточной модели побегообразования. Онтогенез семенных особей на галечнике проходит по следующим фазам морфогенеза: первичный побег (р, j) — первичный куст (im) — куртина (v, gl, g2, g3) - кустящаяся партикула (ss, s). Ход онтоморфогенеза семенных особей, произрастающих на осыпях, схож с онтогенезом особей, растущих на галечнике. Отличие заключается в длительности нахождения особей в фазе куртины, которая у них выделена только в виргинильном состоянии. Морфогенез особей вегетативного происхождения характеризуется следующими фазами: парциальный побег (im) -система парциальных кустов и побегов (v, gl, g2, g3, ss) - парциальный побег (s), где фаза «система парциальных кустов и побегов» является самой длительной. Онтогенез особей D. fragile - полный, сложный и состоит из онтогенеза семенной особи и сокращенных онтогенезов дочерних особей. Вегетативное размножение у особей, растущих на галечнике, начинается в зрелом генеративном состоянии, у особей, произрастающих на осыпи, в виргинильном состоянии, раметы глубоко омоложены до имматурного состояния.
Список литературы Особенности развития Dracocephalum fragile Turcz. ex Benth. (Lamiaceae) в Туве
- Бобров Ю. А. 2009. Грушанковые России. Киров: ВятГГУ. 130 с.
- Гаджиев И.М., Королюк А.Ю., Титлянова А.А. и др. 2002. Степи Центральной Азии. Новосибирск: Изд-во СО РАН. 299 с.
- Гатцук Л. Е. 1974. Геммаксиллярные растения и система соподчиненных единиц их побегового тела//Бюлл. МОИП. Отд. биол. Т. 79. Вып. 1. С. 100-112.
- Гатцук Л.Е. 2008. Растительный организм: опыт построения иерархической системы его структурно-биологических единиц//Современные подходы к описанию структуры растения. Киров: Лобань. С. 26-47.
- Колегова Е.Б., Черемушкина В.А. 2012. Структура побеговых систем видов рода Thymus (Lamiaceae) в Хакасии//Бот. журн. Т. 97. №2. С. 173-182.
- Малышев Л.И. 1965. Высокогорная флора Восточного Саяна (обзор сосудистых растений, особенности состава и флорогенезис). М.-Л.: Наука. 207 с.
- Нухимовский Е. Л. 1997. Основы биоморфологии семенных растений. М.: Недра. Т. 1. 630 с.
- Пешкова Г.А. 1997. Род Dracocephalum L.//Флора Сибири. Новосибирск: Наука. Т.11. 179 с.
- Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах//Труды БИН АН СССР. Сер. 3. Геоботаника. № 6. С. 179 -196.
- Савиных Н.П. 2012. Архитектура трав//Актуальные проблемы современной биоморфологии. Киров: Радуга-ПРЕСС. С. 342-354.
- Савиных Н.П. 2013. Сохранение биоразнообразия с позиций биоморфологии//Вестник Тверского государственного ун-та. Сер. Биология и экология. Вып. 32, № 23. С. 195-209.
- Савиных Н.П., Черёмушкина В.А. 2015. Биоморфология: современное состояние и перспективы//Сибирский экологический журнал. №5. С. 659-670.
- Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования//Учен. Зап. Моск. Гоc. пед. ин-та им. В. П. Потемкина. Вопросы биологии растений: Т. 100. № 5. С. 3-38.
- Серебряков И. Г. 1962. Экологическая морфология растений. М.: Высш. шк. 378 с.
- Серебрякова Т. И. 1980. Еще раз о понятии «жизненная форма» у растений//Бюлл. МОИП. Отд. биол. Т. 85. Вып. 6. С. 75-86.
- Смирнова О.В., Заугольнова Л.Б., Ермакова И.М. и др. 1976. Ценопопуляция растений: (основные понятия и структура). М.: Наука. 216 с.
- Телятников М.Ю. 2016. Растительность восточной части Восточного Саяна (высокогорный и горнолесной пояса). Новосибирск: Академиздат. 104 с.
- Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов//Биол. науки. № 2. С. 7-34.
- Чебакова Н.М., Парфенова Е.И., Бляхарчук Т.А. 2011. Экосистемы и виды//Изменение климата и его воздействие на экосистемы, население и хозяйство российской части Алтае-Саянского экорегиона. М. С. 80-99.
- Urgamal M., Oyuntsetseg B. et al. 2014. Conspectus of the vascular plants of Mongolia. Ulaanbaatar: Admon Printing. 334 p.
