Особенности состава микробных сообществ низкотемпературных минеральных источников Хонгор-Уула (Тункинская долина, Республика Бурятия)
")
Автор: Кашкак Елена Сергеевна, Белькова Наталья Леонидовна, Дагурова Ольга Павловна, Матюгина Евгения Борисовна, Данилова Эржена Викторовна
Журнал: Вестник Бурятского государственного университета. Биология, география @vestnik-bsu-biology-geography
Рубрика: Биология
Статья в выпуске: 2, 2020 года.
Бесплатный доступ
В данной статье представлены результаты гидрохимических и микробиологических исследований воды, осаждений и микробных матов минеральных источников Хонгор- Уула. Вода источников относится к холодным железистым сульфатно-карбонатным, натриево-кальциево-магниевым водам с невысокой минерализацией и слабощелочными значениями рН. В нативных образцах выявлена ферментативная протеолитическая активность, указывающая на деструкцию легкоокисляемого органического вещества микробными сообществами. Наибольшие значения внеклеточной пептидазной активности на изученных субстратах обнаружена в «желудочном» источнике. В микробных сообществах исследуемого источника Хонгор-Уула методами молекулярно-генетического анализа определено разнообразие доминирующих генотипов. Обнаружены представители различных филогенетических линий с доминированием филума Proteobacteria.
Низкотемпературные источники, микробное сообщество, протеолитическая активность, хонгор-уула, республика бурятия
Короткий адрес: https://sciup.org/148316942
IDR: 148316942 | УДК: 579.26 | DOI: 10.18101/2587-7148-2020-2-21-29
Features of the composition of microbial communities of low-temperature mineral springs Khongor-Uula (Tunka valley, Republic of Buryatia)
This article presents the results of hydrochemical and microbiological studies of water, sedimentations and microbial mats of Khongor-Uula mineral springs. The waters of the springs belongs to cold ferrous sulfate-carbonate, sodium-calcium-magnesium type of waters. It characterized by low salinity and slightly alkaline pH values. In native samples enzymatic proteolytic activity was detected.It indicate the destruction of labile organic matter by microbial communities. The high values of extracellular peptidase activity in the “gastric” spring weremeasured. The diversity of dominant genotypes was determined using molecular methods in microbial communities of samples from the Khongor-Uula. In the microbial community, the representatives of various phylogenetic lines were found, the representatives of Proteobacteriadominate.
Текст научной статьи Особенности состава микробных сообществ низкотемпературных минеральных источников Хонгор-Уула (Тункинская долина, Республика Бурятия)
Кашкак Е. С., Белькова Н. Л., Дагурова О. П., Матюгина Е. Б., Данилова Э. В. Особенности cостава микробных сообществ низкотемпературных минеральных источников Хонгор-Уула (Тункинская долина, Республика Бурятия) // Вестник Бурятского государственного университета. Биология, география. 2020. № 2. С. 21–29.
Холодные железистые источники Хонгор-Уула расположены в долине р. Харагун в предгорной части Гурби-Дабанского хребта, окаймляющего с юга Тункинскую котловину (Восточный Саян). В данной местности насчитывается 10 выходов минеральных вод. Выходы железистых вод выявлены на террасах по обоим берегам реки (рис. 1).
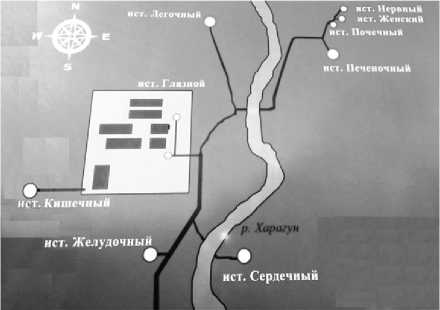
Рис. 1. Карта-схема расположения выходов источника Хонгор-Уула
Нижняя группа выходов воды находится на правобережной пойменной террасе в пределах расширения долины, восточной и западной границей которой являются крутые, местами отвесно падающие, скальные склоны хребта. С южной стороны по центру террасы, занимая примерно до 1/3 ее площади, углом надвинут отрог хребта. Вторая группа источников вытекает из-под края склона хребта на левом берегу реки в 1,5 км от подвесного моста (Геохимическая …, 2011).
Целью работы было изучение физико-химических условий и состава микробного сообщества источников Хонгор-Уула.
Объекты и методы исследования
Объектами исследования являлись вода, донные отложения и микробные маты источников Хонгор-Уула. На месте отбора проб определяли температуру, рН и минерализацию воды портативными приборами. При исследовании физико-химического состава воды использовались потенциометрический, титриметрический и фотометрический методы анализа согласно нормативным документам. Всего было проанализировано 6 проб воды из шести разных источника (таблица 1).
Внеклеточную протеолитическую активность из полученных супернатантов определяли согласно методу Эрлангера (Erlanger et al., 1961). В работе использовали следующие паранитроанилидные субстраты, специфичные для эндопептидаз (рабочая концентрация 5 мМ): GlpAALpNA — субтилизин-подобных пептидаз; BzRpNA (BAPA) — трипсин-подобных пептидаз; GlpFpNA — химотрипсин-подобных пептидаз; LpNA (L-лейцил-паранитроанилид) и FpNA (L-фенилаланил-пара-нитроанилид) — аминопептидаз.
Выделение ДНК проводили коммерческими наборами ДНК-сорб-Б и РИБО-сорб по протоколам фирмы-производителя (ЦНИИ эпидемиологии Роспотребнадзора, Москва). ДНК, выделенную разными методами, использовали в качестве матрицы в полимеразной цепной реакции (ПЦР) с праймерами, комплементарными наиболее консервативным участкам гена 16S рРНК бактерий (в скобках дана нумерация нуклеотидов по Escherichia coli) 500L (514–533) и 1350R (1389–1407). Целевые ампликоны лигировали в векторе из коммерческого набора GeneJETTM PCR CloningKit (Fermentas, Литва), все реакции проводили по протоколу фирмы-производителя. Полученной лигазной смесью трансформировали компетентные клетки E. coli (штамм XL-1) по стандартной методике (Sambrook et al., 1989). Анализ выросших колоний проводили методом ПЦР на плазмидных праймерах с использованием грубого лизата бактериальных клеток в качестве матрицы. Нуклеотидные последовательности определяли на автоматическом секвенаторе ABI 3130×l (Genetic Analyzer, США). Сравнительный анализ нуклеотидных данных выполнен с использованием пакета программ FASTA .
Результаты исследования и их обсуждение
Источники Хонгор-Уулы — холодные железистые сульфатно-карбонатные, натриево-кальциево-магниевые с невысокой минерализацией. Вода источников имеет слабощелочные значения рН. Температура воды не поднимается выше 5,0ºС, суммарный дебит составляет 250 м3 в сутки (табл. 1). В водах источника Хонгор-Уулы обнаружено достаточно высокое содержание общего железа (11 мг/л).
Таблица 1
Физико-химические показатели воды минеральных источников Хонгор-Уулы
|
№ |
Обозначение пробы |
Наименование источника |
t, ºС |
рН |
Минерализация, мг/л |
|
1 |
ХУ1 |
Печеночный |
4,6 |
7,6 |
219 |
|
2 |
ХУ2 |
Почечный |
4,7 |
7,5 |
153 |
|
3 |
ХУ3 |
Нервный |
4,6 |
7,8 |
180 |
|
4 |
ХУ4 |
Женский |
4,7 |
7,7 |
181 |
|
5 |
ХУ5 |
Желудочный |
3,9 |
7,4 |
90 |
|
6 |
ХУ6 |
Глазной |
5,0 |
7,7 |
130 |
Внеклеточная протеолитическая активность в нативных образцах
Протеолитические бактерии являются инициаторами процесса разложения белоксодержащих веществ в микробных сообществах. В зависимости от экологических факторов активность протеолитических ферментов может проявляться в разной степени — активироваться, либо инактивироваться. В пробах микробных матов, ила и воды была изучена внеклеточная протеолитическая активность.
В результате анализа в исследуемых образцах была обнаружена различная внеклеточная активность фермента по всем субстратам (таблица 2). Максимальную активность наблюдали в нативном образце ХУ5 (источник «желудочный») по отношению к субстратам для аминопептидаз LpNa (0,742 ед.) и FpNA (0,681 ед.), а также для субтилизин-подобных пептидаз GlpAALpNA (0,530 ед.).
Таблица 2
Пептидазная активность в нативных образцах источника Хонгор-Уула
|
Обозначение пробы |
Наименование источника |
Субстраты |
||||
|
GlpAALpNA |
BzRpNA (BAPA) |
GlpFpNA |
LpNa |
FpNA |
||
|
ХУ1 |
печеночный |
0,094 |
0,062 |
0,049 |
0,198 |
0,206 |
|
ХУ2 |
почечный |
0,040 |
0,039 |
0,001 |
0,173 |
0,081 |
|
ХУ3 |
нервный |
0,056 |
0,028 |
0,009 |
0,091 |
0,036 |
|
ХУ4 |
женский |
0,040 |
0,037 |
0,013 |
0,089 |
0,070 |
|
ХУ5 |
желудочный |
0,530 |
0,111 |
0,006 |
0,742 |
0,681 |
Активность на трипсин подобном субстрате BzRpNA (BAPA) варьировала от 0,028 до 0,111 ед., при этом максимальная активность обнаружена в образце ХУ5. Химотрипсин-подобная активность (GlpFpNA) была достаточно низкая во всех исследуемых образцах и достигала 0,049 ед. в образце источника ХУ1. В нативных образцах ХУ2, ХУ3 и ХУ4 наблюдали низкие значения активности по всем субстратам.
Таким образом, в разных выходах низкотемпературного источника Хонгор-Уулы наблюдали протеолитическую активность, что указывает на деструкцию легко окисляемого органического вещества микробными сообществами. Отмечено, что наибольшая внеклеточная пептидазная активность по всем субстратам обнаружена в образце источника «желудочный» (ХУ5).
Идентификация доминирующих генотипов в микробных сообществах. В микробных сообществах воды, взвеси, осадка и микробного мата минерального источника Хонгор-Уулы было определено разнообразие доминирующих генотипов in situ методами молекулярно-генетического анализа.
В результате определения таксономического разнообразия прокариот в образцах воды, взвеси, микробного мата и ила из исследуемых источников Хонгор-Уулы были выявлены филотипы, принадлежащие домену Bacteria . В микробных сообществах обнаружены представители 5 филогенетических линий с доминированием Proteobacteria (таблица 3).
Таблица 3
Таксономическое разнообразие прокариот в низкотемпературных источниках Хонгор-Уулы
|
Идентификация на основании рибосомной филогении |
Легочный |
Желудочный |
Филогения |
||
|
вода |
взвесь |
осаждения |
микробный мат |
||
|
Proteobacteria |
|||||
|
Devosia sp. |
1 |
Alphaproteobacteria, Rhizobiales, Hyphomicro-biaceae |
|||
|
Phyllobacterium sp. |
1 |
Alphaproteobacteria, Rhizobiales, Phyllobacte-riaceae |
|||
|
Mesorhizobium sp. |
1 |
Alphaproteobacteria, Rhizobiales, Phyllobacte-riaceae |
|||
|
Sphingobium sp. |
2 |
Alphaproteobacteria, Sphingomonadales, Sphin-gomonadaceae |
|||
|
Sphingomonas sp. |
1 |
Alphaproteobacteria, Sphingomonadales, Sphin-gomonadaceae |
|||
|
Novosphingobium sp. |
1 |
Alphaproteobacteria, Sphingomonadales, Sphin-gomonadaceae |
|||
|
Sideroxydans sp. |
1 |
Betaproteobacteria, Gallio-nellales, Gallionellaceae |
|||
|
Gallionella sp. |
7 |
1 |
Betaproteobacteria, Gallio-nellales, Gallionellaceae |
||
|
Leptothrix sp. |
1 |
Betaproteobacteria, Burk-holderiales, Burkholderia-les genera incertaesedis |
|||
|
Acinetobacter sp. |
3 |
Gammaproteobacteria, Pseudomonadales, Mo-raxellaceae |
|||
|
Pseudomonas sp. |
1 |
1 |
Gammaproteobacteria, Pseudomonadales, Pseudo-monadaceae |
||
|
Geobacter sp. |
2 |
Deltaproteobacteria, Desulfuromonadales, Geobacte-raceae |
|||
|
Bacteroidetes |
|||||
|
Flavobacterium sp. |
1 |
Flavobacteriia, Flavobacte-riales, Flavobacteriaceae |
|||
|
Actinobacteria |
|||||
|
Propionibacterium sp. |
1 |
Actinobacteridae, Actino-mycetales, Propionibacteri-neae, Propionibacteriaceae |
|||
|
Leifsonia sp. |
1 |
Actinobacteridae, Actino-mycetales, Micrococcineae, Microbacteriaceae |
|||
|
Verrucomicrobia |
|||||
|
Luteolibacter sp. |
1 |
Verrucomicrobiae, Verrucomicrobiales, Verrucomicrobiaceae |
|||
|
Opitutus sp. |
1 |
Opitutae, Opitutales, Opitutaceae |
|||
|
Cyanobacteria |
|||||
|
Symploca sp. |
7 |
Oscillatoriophycideae, Oscillatoriales |
|||
Тип и лечебные качества воды источника «желудочный» определяются доминирующим содержанием железа (более 10 мг/л). В «желудочном» источнике были обнаружены нити микробных матов светло-коричневого цвета. В микробном мате данного источника были выявлены представители цианобактерии рода Symploca , анаэробных бактерий рода Geobacter , окисляющих органические вещества и железо, а также бактерии рода Leptothrix , доминирующих во многих железоокисляющих пресноводных сообществах (Emerson, Ghiorse, 1992, Schmidt et al., 2014). Бактерии рода Geobacter в последнее время становятся объектом пристального внимания со стороны ученых. Данные бактерии способны строить биологические структуры, отличающиеся электропроводностью, производить «микробную нанопроволоку» (Sun et al., 2014).
В «легочном» источнике в воде была обнаружена видимая взвесь, которую отфильтровали на крупнопористый, поликарбонатный фильтр с диаметром пор 5 мкм и проанализировали отдельно — фильтрат в качестве фракции свободноживущих бактериальных клеток, взвесь — как ассоциированные с минеральными частицами бактериальные клетки, а также осаждения — сформированный минеральный осадок, так называемые современные минеральные осаждения. В осадках и взвеси легочного источника идентифицированы микроорганизмы, участвующие в окислении железа — Gallionella sp., Acinetobacter sp., Pseudomonas sp., Flavobacterim sp.
В составе микробных сообществ низкотемпературных источников Хонгор-Уула отмечены особенности, связанные с экологической специализацией близких видов для реализации сходных метаболических путей в одном месте обитания. Наиболее ярко это отмечено для представителей альфапротеобактерий — разные представители порядка Rhizobiales детектировались в воде и современных минеральных осаждениях. Они были представлены бактериями, относящимися к разным видам — Devosia sp.,
Phyllobacterium sp., Mesorhizobium sp., представители которых примечательны своей способностью устанавливать симбиотические отношения с бобовыми растениями (Mantelin et al., 2006, Balachandar et al., 2007, Yoon et al., 2007, Degefu et al., 2013). Еще один пример — в воде, взвеси и осаждениях обнаружены разные виды семейства Sphingomonadaceae .
Также экологическая специализация отмечена на уровне разных мест обитания. В современных минеральных осаждениях источника «легочный» и бактериальных матах источника «желудочный» идентифицированы бактерии, биогеохимическая деятельность которых способствует формированию разного вида современных минеральных осаждений. В источнике «желудочный» за счет деятельности Gallionella sp., Leptothrix sp., Geobacter sp. происходит трансформация растворимых соединений, получение взвешенного вещества и формирование минерального осадка.
Таким образом, результаты нашего исследования выявили разнообразие и широкое распространение представителей 5 филогенетических линий с доминированием Proteobacteria. Идентифицированные протеобактерии в исследуемых образцах являются преимущественно гетеротрофами, которые проводят деструкцию органического вещества в кислородных или микроаэрофильных условиях.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 18–34–00552.
Список литературы Особенности состава микробных сообществ низкотемпературных минеральных источников Хонгор-Уула (Тункинская долина, Республика Бурятия)
- Геохимическая деятельность микроорганизмов гидротерм Байкальской рифтовой зоны / Б. Б. Намсараев, Д. Д. Бархутова, Э. В. Данилова и др.-Новосибирск: Академическое изд-во "Гео", 2011.- 302 с.
- Balachandar D., Raja P., Kumar K., Sundaram SP. Non-rhizobial nodulation in legumes // Biotechnology and Molecular Biology Review 2007. Vol. 2 (2). P. 49-57: DOI: 10.13140/2.1.1775.2960
- Degefu T., Wolde-meskel E., Liu B., Cleenwerck I., Willems A., Frostegard A. Mesorhizobium shonense sp. nov., Mesorhizobium hawassense sp. nov. and Mesorhizobium abyssinicae sp. nov., isolated from root nodules of different agroforestry legume trees Free // International Jurnal of Systematic and Evolutionary Microbiology. 2013. V63 (5). P. 1746-1753: https://doi. org/. DOI: 10.1099/ijs.0.044032-0
- Emerson D., Ghiorse W. C. Isolation, cultural maintenance and taxonomy of a sheath- forming strain of Leptothrixdiscophora and characterization of manganese-oxidizing activity associated with the sheath // Appl. Environ. Microbiol. 1992. V 58. P. 4001-4010.
- Erlanger B. F., Kokowsky N., Cohen W. The preparation and properties of two new chromogenic substrates of trypsin // Arch. Biochem. Biophys. 1961. V. 95. (2). P. 271-278.
- Mantelin S., Saux M. F., Zakhia F., Cleyet-Marel J. Emended description of the genus Phyllobacterium and description of four novel species associated with plant roots: Phyllobacterium bourgogense sp. nov., Phyllobacterium ifriqiyense sp. nov., Phyllobacterium leguminumsp. nov., and Phyllobacterium brassicacearum sp. nov. //International Journal of Systematic and Evolutionary Microbiology. 2006. Vol. 56. P. 827-839: DOI: 10.1099/ijs.0.63911-0
- Sambrook J., Fritsch E. F., Maniatis T. Molecular Coning. A Laboratory Manual // Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 1989. Vol. 1, 2, 3.
- Schmidt B., Sanchez L. A., Fretschner T., Kreps G., Ferrero M. A., Sineriz F., Szew- zyk U. Isolation of Sphaerotilus-Leptothrix strains from iron bacteria communities in Tierra del Fuego wetlands // FEMS Microbiology Ecology. 2014. Vol. 90(2). P. 454-466.https://doi. org/ DOI: 10.1111/1574-6941.12406
- Sun D., Wang A., Cheng S., Yetes M. D., Logan B. E. Geobacteranodireducens sp. nov., an exoelectrogenic microbe in bioelectrochemical systems // International Journal of Systematic and Evolutionary Microbiology. 2014. Vol. 64 (Pt. 10). P. 3485-3491. DOI: 10.1099/ijs.0.061598-0
- Yoon J.-H., Kang S.-J., Park S., Oh T.-K. Devosiainsulae sp. nov., isolated from soil, and emended description of the genus Devosia // International Journal of Systematic and Evolutionary Microbiology. 2007. Vol. 57 (Pt 6). P. 1310-1314: DOI: 10.1099/ijs.0.65028-0
