Особенности влияния инсулиноподобного фактора роста 1 и фактора роста фибробластов 2 на качество созревающих ооцитов и их способность к эмбриональному развитию in vitro у крупного рогатого скота (Bos taurus taurus)
")
Автор: Сингина Г. Н., Шедова Е. Н., Лопухов А. В., Жукова А. С.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Репродуктивные технологии
Статья в выпуске: 6 т.60, 2025 года.
Бесплатный доступ
Экстракорпоральное созревание (in vitro maturation, IVM) - неотъемлемый этап получения эмбрионов in vitro (in vitro embryo production, IVP). Условия IVM критически влияют на качество яйцеклеток и развившихся из них IVP эмбрионов, но в настоящее время остаются субоптимальными и требуют усовершенствования. В настоящей работе мы впервые установили, что коровьи ооциты, созревшие in vitro при воздействии IGF1 и FGF2, а затем стареющие при пролонгированном культивировании в их отсутствие, различаются по способности развиваться до стадии бластоцисты после экстракорпорального оплодотворения. Цель работы заключалась в сравнительном изучении влияния двух клеточных цитокинов (инсулиноподобного фактора роста 1 и фактора роста фибробластов 2) в среде in vitro созревания коровьих ооцитов на их качество, способность к эмбриональному развитию, а также устойчивость к возрастным трансформациям. Яичники половозрелых коров ( Bos taurus taurus ), собранные после убоя животных, были доставлены в лабораторию в физиологическом растворе в течение 4-6 ч при температуре не ниже 28 °С. Ооциты в составе ооцит-кумулюсных комплексов (ОКК) выделяли из фолликулов яичников механическим способом: рассекали стенки видимых фолликулов лезвием, после чего проводили поиск и промыв извлеченных ОКК. Группы из отобранных 20-25 ОКК созревали in vitro в контрольной среде ТС-199С, содержащей HEPES (25 мМ), Na-пируват (0,5 мМ), фолликулостимулирующий и лютеинизирующий гормоны (каждый по 10 мкг/мл), эпидермальный фактор роста (20 нг/мл), а также фетальную бычью сыворотку (ФБС, 10 %) и антибиотик гентамицин (50 мкг/мл) в течение 20 ч. В опытных группах в среде IVM аналогичного состава дополнительно присутствовал IGF1 («Thermo Fisher Scientific, Inc.», США) или FGF2 («Thermo Fisher Scientific, Inc.», США) в концентрации 20 и 40 нг/мл, соответственно. После IVM часть созревших ОКК переносили в среду старения (ТС-199 c 10 % ФБС) и культивировали дополнительно в течение следующих 12 или 24 ч. Непосредственно после IVM и через 24 ч старения в ооцитах оценивали степень апоптоза (метод TUNEL), а в первом случае дополнительно проводили анализ стадий мейоза. Процесс ядерного созревания оценивали по доле ооцитов на различных стадиях мейоза к общему числу культивируемых ооцитов, апоптоз - по доле TUNEL-позитивных ооцитов на стадии метафазы II от общего числа МII-ооцитов. Часть созревших ооцитов, а также ооциты, стареющие в течение 12 ч, подвергали экстракорпоральному оплодотворению (ЭКО) и культивированию для эмбрионального развития. Для оценки качества полученных бластоцист готовили и анализировали их цитологические препараты. Степень созревания ооцитов - достижение стадии метафазы II (MII) мейоза - значимо не различалась между группами и составила 76,5-82,5 %. В то же время воздействие IGF1 и FGF2 на ооциты приводило к снижению частоты апоптотических изменений MII хромосом: через 20 ч IVM - с 22,5±1,4 в контроле до 7,6±1,4 и 12,3±0,2 %; после 24 ч старения - с 40,8±1,8 в контроле до 23,2±1,0 и 19,8±0,78 % (p function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Инсулиноподобный фактор роста 1, фактор роста фибробластов 2, in vitro созревание, старение, эмбриональное развитие, апоптоз, крупный рогатый скот
Короткий адрес: https://sciup.org/142247712
IDR: 142247712 | УДК: 636.2: | DOI: 10.15389/agrobiology.2025.6.1085rus
IGF1 and FGF2 effects on the quality of maturing cattle (Bos taurus taurus) oocytes and their competence for embryonic development in vitro
In vitro maturation (IVM) is an essential component of in vitro embryo production (IVP). The conditions under which IVM is performed critically determine the quality of oocytes and the resulting IVP embryos; however, current maturation protocols remain suboptimal and require further improvement. In the present study, we demonstrate for the first time that bovine oocytes matured in vitro in the presence of IGF1 or FGF2, and subsequently subjected to prolonged cytokine-free culture, differ in their ability to develop to the blastocyst stage following in vitro fertilization (IVF). The aim of this work was to comparatively assess the effects of two cytokines, the insulin-like growth factor 1 (IGF1) and fibroblast growth factor 2 (FGF2) when added to the oocyte in vitro maturation medium, focusing on oocyte quality, embryonic developmental potential, and resistance to age-related alterations. Ovaries from sexually mature cows ( Bos taurus taurus ) were collected post-mortem and transported to the laboratory in physiological saline within 4-6 h at a temperature not below 28 °C. Cumulus-oocyte complexes (COCs) were mechanically isolated from visible follicles by dissecting follicular walls with a scalpel blade, followed by retrieval and washing of the collected complexes. Groups of 20-25 selected COCs were matured in vitro for 20 h in control TC-199C medium supplemented with HEPES (25 mM), sodium pyruvate (0.5 mM), follicle-stimulating and luteinizing hormones (10 mg/ml each), epidermal growth factor (20 ng/ml), fetal bovine serum (10 %), and gentamicin (50 mg/ml). Experimental groups received identical medium supplemented with IGF1 (20 ng/ml) or FGF2 (40 ng/ml). After IVM, a subset of matured COCs was transferred into an aging medium (TC-199 with 10 % serum) and cultured for an additional 12 or 24 h. Immediately after IVM and after 24 h of aging, apoptosis was evaluated using the TUNEL assay; in the former case, meiotic staging was additionally performed. Nuclear maturation was quantified as the rate of oocytes at specific meiotic stages, while apoptosis was assessed as the rate of TUNEL-positive metaphase II (MII) oocytes among the total MII population. A portion of matured oocytes and oocytes aged for 12 h underwent IVF and subsequent embryo culture. Blastocyst quality was assessed using cytological preparations. Oocyte maturation rate (progression to meiosis II metaphase, MII) did not differ between groups, ranging from 76.5 % to 85.2 %. However, IGF1 and FGF2 treatments reduced apoptotic degeneration in MII oocytes: from 22.5±1.4 % in control to 7.6±1.4 % and 12.3±0.2 % after 20 h IVM, respectively (p function show_eabstract() { $('#eabstract1').hide(); $('#eabstract2').show(); $('#eabstract_expand').hide(); }
Текст научной статьи Особенности влияния инсулиноподобного фактора роста 1 и фактора роста фибробластов 2 на качество созревающих ооцитов и их способность к эмбриональному развитию in vitro у крупного рогатого скота (Bos taurus taurus)
Эффективность вспомогательных репродуктивных технологий (ВРТ), направленных на получение эмбрионов вне организма, во многом зависит
Работа выполнена при финансовой поддержке Российского научного фонда (проект ¹ 24-16-00261).
от качества ооцитов, которое они приобретают в процессе созревания in vitro (in vitro maturation, IVM) (1, 2). Аналогично созреванию in vivo, в выделенных из фолликулов и инкубируемых вне организма ооцитах возобновляется мейоз и происходят изменения хромосом от стадии диплотены 1-го деления мейоза до метафазы II деления мейоза (МII). Тем не менее созревание цитоплазмы ооцитов, когда в ней наблюдаются необходимые для их последующего оплодотворения ультраструктурные и молекулярные перестройки, оказывается недостаточным, и полноценность яйцеклеток снижается (1, 3). Причина — неадекватность условий экстракорпорального созревания женских половых клеток, что требует совершенствования соответствующих методик (4). При этом воспроизведение физиологической среды, характерной для овариальных фолликулов, где происходит созревание ооцитов in vivo, представляется наиболее логичным и перспективным подходом (2, 4, 5).
Чтобы имитировать внутрифолликулярные условия, в среды созревания ооцитов добавляют гормоны и различные метаболиты, к которым, в частности, относится инсулиноподобный фактор роста 1 (insulin-like growth factor 1, IGF1) (6). Показано, что IGF1 задействован в аутокринной и паракринной регуляции функции яичников самок и может влиять на гранулезные и половые клетки, контролируя пролиферацию, дифференцировку и стероидогенез а также созревание ооцитов (7, 8). Рецепторы IGF1 выявлены в ооцитах на всех стадиях их развития, и этот фактор роста в большом количестве секретируется окружающим ооциты соматическим компартмен-том, в частности клетками кумулюса (9, 10). В условиях in vitro на различных видах животных показано, что присутствие IGF1 в среде IVM ускоряет мейоз и снижает степень апоптоза в ооцитах, а также способствует увеличению выхода эмбрионов доимплантационных стадий развития (10, 11).
Другим известным цитокином, способным влиять на эффективность получения эмбрионов in vitro (in vitro embryo production, IVP), служит фактор роста фибробластов 2 (fibroblast growth factor 2, FGF2) (12, 13). Установлено, что FGF2 действует как ангиогенный стимулятор, регулирующий стероидогенез в гранулезных клетках, повышая их выживаемость, и он усиливает активность тканевого активатора плазминогена, тем самым улучшая созревание ооцитов (14, 15). Кроме того, FGF2 и его рецептор FGFR2 экспрессируются как в незрелых, так и зрелых ооцитах (16, 17).
У млекопитающих ооциты, достигнув стадии МII, вступают в период ожидания оплодотворения. Если последнее не происходит в течение определенного времени, то в созревших яйцеклетках инициируется постовуляторное старение, что негативно влияет на их качество (18). В таких ооцитах происходят различные морфологические и функциональные изменения, в первую очередь деструктивные изменения хромосом, спонтанная партеногенетическая активация, апоптоз и аномальные преобразования органелл (19, 20), что приводит к нарушению развития эмбрионов и здоровья потомства (21). Показано также, что быстрая потеря полноценности ооцитов на стадии МII служит одним из факторов, ограничивающих эффективность ВРТ у разных видов, включая человека (22, 23).
В условиях in vitro используемые для культивирования гаметы, изначально неоднородны по потенциалу развития. Часть ооцитов может завершать ядерное созревание уже к 16 ч IVM (24), то есть значительно раньше, чем их подвергают процедуре экстракорпорального оплодотворения (ЭКО), которое проводят, как правило, через 24 ч IVM. Как следствие, в этих яйцеклетках к моменту ЭКО возникают возрастные изменения, которые приводят к потере ими качества по сравнению клетками, достигшими
МII позднее (19). В этой связи при оптимизации условий IVM следует учитывать не только качественные изменения в созревающих ооцитах, но и характер влияния тестируемых факторов на устойчивость яйцеклеток к возрастным трансформациям (25).
В настоящей работе мы впервые установили, что коровьи ооциты, созревшие in vitro при воздействии IGF1 и FGF2, а затем стареющие при пролонгированном культивировании в их отсутствии, различаются по способности развиваться до стадии бластоцисты после экстракорпорального оплодотворения.
Цель работы заключалась в сравнительном изучении влияния двух клеточных цитокинов (инсулиноподобного фактора роста 1 и фактор роста фибробластов 2) в среде in vitro созревания коровьих ооцитов на их качество, способность к эмбриональному развитию, а также устойчивость к возрастным трансформациям.
Методика. В экспериментах, выполненных в 2024 году, использовали реагенты фирмы «Sigma-Aldrich» (США) (за исключением отдельно отмеченных случаев).
Культивирование проводили в четырехлуночных планшетах (ООО «Биомедикал», Россия) в каплях среды объемом 500 мкл, покрытых равным объемом минерального масла. Для ооцитов это происходило в условиях инкубатора MCO-18AIC («Sanyo», Япония) при 38,5 ° С и 5 % СО 2 в атмосфере, для эмбрионов — в планшетном инкубаторе BT-37 («Planer Limited», Великобритания) при аналогичной температуре, но содержащем в атмосфере 5 % СО 2 , 5 % О 2 и 90 % N 2 .
Яичники половозрелых коров ( Bos taurus taurus ) неизвестной породной принадлежности, собранные после убоя животных, были доставлены в лабораторию в физиологическом растворе (ФЗ) в течение 4-6 ч при температуре не ниже 28 ° С. После доставки их освобождали от лишних тканей и многократно промывали в стерильном подогретом до 37 ° С ФЗ с антибиотиками, чтобы удалить компоненты крови и микробную контаминацию. Ооциты в составе ооцит-кумулюсных комплексов (ОКК) выделяли из фолликулов яичников механическим способом: рассекали стенки видимых фолликулов лезвием, после чего проводили поиск и промывку извлеченных ОКК, как было описано ранее (26). При этом в качестве рабочего раствора применяли среду ТС-199M с буфером HEPES (25 мМ), 2 % фетальной бычьей сыворотки (ФБС; ООО «БиолоТ», Россия), 10 мкг/мл гепарина и 50 мкг/мл гентамицина. На завершающем этапе промывки выполняли морфологическую оценку выделенных ОКК, отбирая ооциты с гомогенной цитоплазмой и окруженные многослойным компактным кумулюсом.
Группы из отобранных 20-25 ОКК созревали in vitro в контрольной среде ТС-199С, содержащей HEPES (25 мМ), Na-пируват (0,5 мМ), фолликулостимулирующий и лютеинизирующий гормоны (каждый по 10 мкг/мл), эпидермальный фактор роста (20 нг/мл), а также ФБС (10 %) и антибиотик гентамицин (50 мкг/мл) в течение 20 ч. В опытных группах в среде IVM аналогичного состава дополнительно присутствовал IGF1 («Thermo Fisher Scientific, Inc.», США) или FGF2 («Thermo Fisher Scientific, Inc.», США) в концентрации соответственно 20 и 40 нг/мл. Действие каждого цитокина было исследовано индивидуально. При выборе концентраций руководствовались ранее проведенными исследованиями, показавшими их эффективность в среде созревания ооцитов при получении партеногенетических эмбрионов КРС (27). После IVM часть созревших ОКК переносили в среду старения и культивировали дополнительно в течение следующих 12 или 24 ч. При пролонгированном культивировании применяли среду ТС-199С с исключением из ее состава гормонов.
В первом эксперименте по завершении периодов созревания (20 ч IVM) и старения (20 ч IVM + 24 ч пролонгированного культивирования) ооциты освобождали от окружающих клеток кумулюса посредством кратковременной обработки ОКК в 0,1 % растворе гиалуронидазы и последующего пипетирования комплексов через капилляр с внутренним диаметром 135 мкм в ТС-199М. Изолированные ооциты промывали в фосфатно-солевом буфере (ФСБ), содержащем 0,1 % бычьего сывороточного альбумина (ФСБ-БСА), и фиксировали 4 % раствором параформальдегида в ФСБ в течение 1 ч при комнатной температуре.
После промывания в ФСБ-БСА образцы подвергали процедуре пер-меабилизации в 0,5 % растворе Triton Х-100 в течение 30 мин. Наличие признаков апоптоза ядерного материала в ооцитах оценивали методом TUNEL с использованием набора In Situ Cell Death Detection Kit, fluorescein («Roche Diagnostics», Швейцария) согласно инструкции компании-производителя. Затем ооциты обрабатывали в течение 20 мин раствором DAPI (1 мкг/мл) для окрашивания хромосом и переносили на предметное стекло с последующим заключением в среду Vectashield («Vector Laboratories», Великобритания).
Микрофотографирование и анализ препаратов выполняли под микроскопом Axio Imager M2, оснащенным флуоресцентной приставкой, с использованием программы Zen pro («Carl Zeiss AG», Германия). На препаратах исследовали состояние хромосом, относя их к одной из следующих стадий развития: стадии зародышевого пузырька (ЗП); стадиям диакинез/мета-фазы (Дк-MI), анафазы (AI) и телофазы (TI) 1-го мейотического деления и стадии метафазы 2-го мейотического деления (MII). Также определяли наличие или отсутствие TUNEL-позитивного окрашивания хромосом на стадии метафазы II.
Процесс ядерного созревания оценивали по доле ооцитов на различных стадиях мейоза к общему числу культивируемых ооцитов, апоптоз — по доле TUNEL-позитивных ооцитов на стадии метафазы II от общего числа МII-ооцитов.
Во втором эксперименте по завершении периодов созревания (20 ч IVM) и старения (20 ч IVM + 12 ч пролонгированного культивирования) ооциты подвергали процедуре ЭКО с целью получения эмбрионов in vitro. Экстракорпоральное оплодотворение проводили в среде BO-IVF («IVF Bioscience», Великобритания). Бычью сперму размораживали, и подвергали процедуре swim-up c использованием среды Sperm-TALP (28). Полученную фракцию активных сперматозоидов, вносили в среду ЭКО с предварительно перенесенными туда ОКК до конечной концентрации 1,5½10 6 сперматозоидов на 1 мл. Продолжительность совместной инкубации гамет составляла 15-16 ч, после чего освобожденные от кумулюсных клеток и ненужных сперматозоидов ооциты переносили в среду BO-IVC («IVF Bioscience», Великобритания) и инкубировали в ней для эмбрионального развития. Через 3 сут культивирования in vitro (IVC) оценивали число раздробившихся ооцитов, на 7-е сут инкубации определяли число ооцитов, развившихся до стадии бластоцисты.
Для оценки качества полученных бластоцист готовили и анализировали их цитологические препараты, используя протокол, описанный выше для ооцитов, с одним отличием — продолжительность фиксирования в параформальдегиде была снижена до 30 мин.
Статистическую обработку данных проводили при помощи лицензионного программного пакета SigmaStat 2.03.0 («Systat Software, Inc.», США). 1088
Для каждой группы выполняли по 4 независимых опыта. Результаты выражали как средние значения ( M) и стандартные ошибки средних (±SEM). Для оценки статистической значимости различий при анализе двух групп (влияние старения) использовали t -критерий Стьюдента. Для множественных сравнений между группами (влияние факторов) применяли однофакторный дисперсионный анализ (ANOVA) с поправкой Тьюки. Различия считали достоверными при p < 0,05.
Результаты. На основании оценки цитологических препаратов были получены данные о влиянии IGF1 и FGF2 на характер ядерных преобразований в ооцитах, созревающих in vitro (рис. 1). В контроле после 20 ч созревания in vitro 76,5±1,19 % ооцитов находились на стадии метафазы II. Введение тестируемых цитокинов в среду IVM не повлияло на завершение ядерного созревания: доля MII ооцитов в группе IGF1 составила 82,5±2,5 %, FGF2 — 81,7±1,2 %. Тем не менее были получены данные, свидетельствующие об ускоряющем воздействии на мейотический процесс одного из факторов. В присутствии FGF2 доля ооцитов на стадии АI составила 0 % и была значимо ниже, чем в контроле (р < 0,05). Наблюдалось также отсутствие ооцитов на стадии ТI, но без достоверных различий.
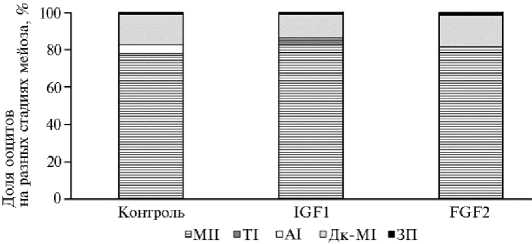
Рис. 1. Ядерные преобразования в ооцитах коров ( Bos taurus taurus ) после созревания в течение 20 ч в отсутствие (контроль) и присутствии инсулиноподобного фактора роста (IGF1, 20 нг/мл) и фактора роста фибробластов (FGF, 40 нг/мл): Дк-MI — диакинез/метафаза I, AI — анафаза I, TI — телофаза I, MII — метафаза II, ЗП — ооциты на стадии зародышевого пузырька (ФГБНУ ФИЦ животноводства — ВИЖ им. академика Л.К. Эрнста, 2024 год).
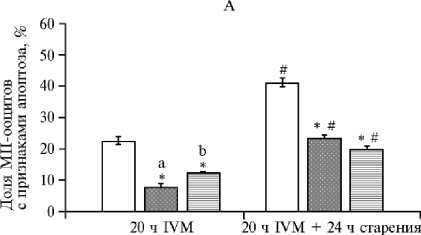
Выявлено положительное воздействие IGF1 и FGF2 на качество MII хромосом (рис. 2).
Через 20 ч IVM в контроле доля ооцитов с признаками апоптоза составила 22,5±1,44 %. Присутствие IGF1 и FGF2 с высокой достоверностью снижало этот показатель — соответственно до 7,55±1,42 и 12,3±0,17 % (р < 0,001). При этом между опытными вариантами наблюдались различия в пользу IGF1 (р < 0,05).
После пролонгированного культивирования (старения in vitro) в контроле доля зрелых ооцитов с апоптотическими изменениями MII хромосом также была наибольшей и составила 40,8±1,80 % (см. рис. 2). Кроме того, сохранялся положительный эффект тестируемых факторов аналогичный наблюдавшемуся после 20-часового IVM. По сравнению с контролем как IGF1, так и FGF2 существенно снижали степень апоптоза в ооцитах соответственно до 23,2±1,03 и 19,8±0,78 % (р < 0,001).
В целом, вне зависимости от условий созревания 24-часовое пролонгированное культивирование зрелых ооцитов приводило к значимому повышению апоптотической дегенерации MII хромосом во всех экспериментальных группах (р < 0,001). Тем не менее для опытных групп (IGF1 и FGF2) этот показатель снижался до значений, сопоставимых с контролем до начала периода старения.
□ Контроль □ IGF1 В FGF2
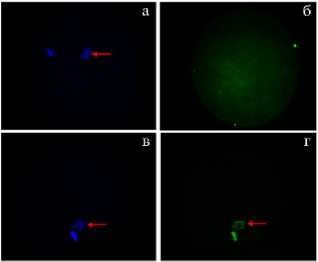
Рис. 2. Апоптотическая дегенерация MII-хромосом в ооцитах коров ( Bos taurus taurus ) после 20часового созревания in vitro (IVM) в контроле, при воздействии инсулиноподобного фактора роста (IGF1) и фактора роста фибробластов (FGF2) , а также после 24-часового старения (А) ( n = 4, M ±SEM) ; репрезентативные микрофотографии ооцитов коров с признаками апоптоза (Б) : верхний ряд — ооцит без апоптоза, нижний ряд — TUNEL позитивный ооцит; а, в — DAPI окрашивание хромосом на стадии метафазы II (синий цвет, отмечены стрелкой); б, г — окрашивание ядерного материала ооцитов методом TUNEL (зеленый цвет), TUNEL-позитивные хромосомы на стадии MII отмечены стрелкой (увеличение 400½, микроскоп Axio Imager.M2, «Carl Zeiss AG», Германия) (ФГБНУ ФИЦ животноводства — ВИЖ им. академика Л.К. Эрнста, 2024 год).
* Различия с контролем статистически значимы при р < 0,001.
# Различия внутри групп (контроль, IGF1 и FGF2) между показателями при 20 ч IVM и после дополнительного пролонгированного культивирования (24 ч) статистически значимы при р < 0,001.
а, b Различия между группами IGF1 и FGF2 статистически значимы при р < 0,001.
Данные сравнительного анализа влияния IGF1 и FGF2 в период созревания ооцитов на их эмбриональное развитие в условиях in vitro после экстракорпорального оплодотворения представлены в таблице.
Развитие эмбрионов, полученных по технологии IVP (in vitro production) после созревания ооцитов коров ( Bos taurus taurus ) в течение 20 ч при воздействии инсулиноподобного фактора роста (IGF1) и фактора роста фибробластов (FGF2) , а также последующего старения в течение 12 ч ( n = 4, M ±SEM; ФГБНУ ФИЦ животноводства — ВИЖ им. академика Л.К. Эрнста, 2024 год)
|
Показатель |
Контроль |
IGF1 |
FGF2 |
|
Оплодотворение |
через 20 ч созревания |
||
|
Число ооцитов, n |
91 |
87 |
82 |
|
Доля раздробившихся ооцитов, % |
72,6±4,31 |
75,6±6,36 |
76,5±1,30 |
|
Доля ооцитов, развившихся до бластоцисты, % |
21,3±2,52a |
37,0±5,50*c |
39,0±2,40*e |
|
Оплодотворение через 20 |
ч созревания |
и 12 ч старения |
|
|
Число ооцитов, n |
89 |
87 |
82 |
|
Доля раздробившихся ооцитов, % |
63,6±3,33 |
71,4±1,72* |
72,5±0,96* |
|
Доля ооцитов, развившихся до бластоцисты, % |
11,9±1,34b |
23,3±2,92*d |
14,5±0,96f |
* Различия с контролем статистически значимы при р < 0,05.
a-f Различия в группах между показателями при 20 ч созревания и после дополнительного пролонгированного культивирования в течение 12 ч статистически значимы: a, b — при < 0,01, c, d — при р = 0,057, e, f — при р < 0,001.
Когда экстракорпоральное оплодотворение ооцитов происходило сразу после их созревания, доля раздробившихся ооцитов была сходной в сравниваемых группах и варьировала от 72-76 %. Тем не менее созревание ОКК в присутствие IGF1 и FGF2 положительно повлияло на дальнейшее эмбриональное развитие in vitro, приводя в равной степени к достоверному повышению доли ооцитов, развившихся до стадии бластоцисты (см. табл., рис. 3) по сравнению с контролем (р < 0,05).
В то же время результаты оплодотворения ооцитов, которые сначала созревали при воздействии IGF1 и FGF2, а затем дополнительно культивировались (старели in vitro) в течение 12 ч в отсутствие этих факторов 1090
несколько отличались от тех, что представлены выше для ЭКО непосредственно после IVM. При старении в случаях использования обоих цитокинов наблюдалось повышение доли дробления по сравнению с контролем (р < 0,05). Кроме того, IGFI, но не FGF2 оказал позитивное воздействие на долю развития стареющих ооцитов до стадии бластоцисты, повысив этот показатель от значения контроля в 2 раза (р < 0,05).
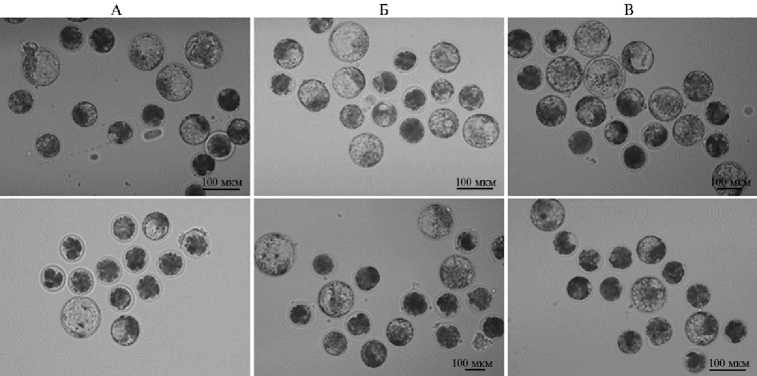
Рис. 3. Микрофотографии 7-суточных эмбрионов коров ( Bos taurus taurus ), развившихся до стадии бластоцисты после in vitro оплодотворения ооцитов, созревших в отсутствие (контроль) и в присутствии IGF1 или FGF2 (верхний ряд) , а также последующего старения в течение 12 ч (нижний ряд) (увеличение 100½, микроскоп Eclipse Ti-U, «Nikon», Япония; ФГБНУ ФИЦ животноводства — ВИЖ им. академика Л.К. Эрнста, 2024 год) .
Сравнение результатов эмбрионального развития до и после пролонгированного культивирования (ПК) показало, что старение привело к значимому снижению выхода бластоцист как в контроле, так и при использовании FGF2 (соответственно р < 0,01 и р < 0,001), в то время как при использовании IGF1 наблюдалась только тенденция к снижению (р = 0,057) (см. табл., рис. 3). Кроме того, при воздействии этого цитокина показатель снижался до значений, сопоставимых с контролем до начала периода старения.
Использование IGF1 и FGF2 повлияло на качество IVP эмбрионов, которое оценивали по доле ядер в бластоцистах с признаками апоптоза относительно их общего числа (рис. 4). В контроле этот показатель до и после старения составил соответственно 5,39±0,34 и 9,80±0,33 %. Оба тестируемых фактора снижали степень апоптоза в эмбрионах, которые развились из ооцитов, оплодотворенных сразу после IVM (р < 0,05). IGF, кроме того, уменьшал долю апоптотических ядер в бластоцистах, развившихся из стареющих ооцитов (р < 0,05).
Качество ооцитов, приобретаемое ими в период экстракорпорального созревания, служит основным фактором, определяющим их последующее эмбриональное развитие (1, 2). В настоящем исследовании, чтобы повысить полноценность яйцеклеток, в среду IVM вносили инсулиноподобный фактор роста (IGF1) и фактор роста фибробластов (FGF2), которые играют важную роль в регуляции оогенеза в фолликуле в условиях in vivo (7, 14). Мы впервые обнаружили, что присутствие IGF1 в среде IVM не только снижает степень апоптоза в созревших ооцитах и увеличивает процент развившихся из них после ЭКО бластоцист, как это было продемонстрировано ранее (10, 11), но и повышает их устойчивость к возрастным трансформациям. Частично сходный, но менее долговременный эффект в отношении старение в аналогичных условиях обнаружен и для другого исследуемого цитокина — FGF2.
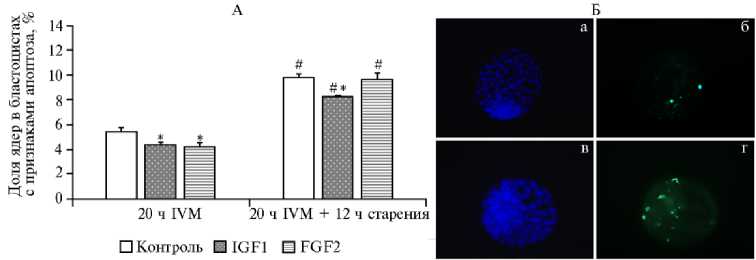
Рис. 4. Степень апоптотической дегенерации ядерного материала бластоцист, развившихся из ооцитов коров ( Bos taurus taurus ) после 20-часового созревания in vitro (IVM) в контроле, при воздействии инсулиноподобного фактора роста (IGF1) и фактора роста фибробластов (FGF2) , а также после 12-часового старения (А) ( n = 4, M ±SEM) ; цитологический анализ степени апоптоза в эмбрионах (Б) : верхний ряд — эмбрион, развившийся до стадии бластоцисты из ооцита до старения, нижний ряд — после старения; а, в — DAPI окрашивание ядер в бластоцисте (синий цвет); б, г — окрашивание ядерного материала бластоцисты методом TUNEL (зеленый цвет) (увеличение 400½, микроскоп Axio Imager.M2, «Carl Zeiss AG», Германия).
* Различия с контролем статистически значимы при р < 0,05.
# Различия внутри групп (контроль, IGF1 и FGF2) между значениями при 20 ч IVM и после дополнительного пролонгированного культивирования (12 ч) статистически значимы при р < 0,001.
Старение — сложный биологический процесс, который сопровождается рядом изменений в структуре и функциональном состоянии зрелого ооцита, негативно влияющих на его качество и возникающих, если яйцеклетка не активируется оплодотворением (24, 29-31). В условиях in vitro, как следует из источников литературы, возрастные трансформации могут происходить в ооцитах в период их созревания, снижая эффективность ВРТ (22, 23).
В настоящем исследовании мы обнаружили, что при пролонгированном культивировании зрелых ооцитов контрольной группы в них значимо (с 22 до 40 %, р < 0,001) повышалась частота апоптотической дегенерации MII хромосом, что согласуется с результатами других исследователей (25), в то время как IGF1 и FGF2 подавляли это повышение (см. рис. 2). Степень апоптоза в ооцитах под воздействием этих факторов в среде IVM при последующем ПК также возрастала, но значения были сопоставимы с таковыми в контроле до ПК. Как и другие авторы (11), мы показали, что IGF1 в условиях IVM снижает апоптоз в созревших ооцитах (с 22 до 7 %), но также продемонстрировали, что это влияние сохраняется и при последующем их старении (23 % в группе IGF1 против 40 % в контроле). Выявленный эффект FGF2 на апоптоз в ооцитах коров описан нами впервые и может быть связан с его антиапоптотическим влиянием на клетки куму-люса, окружающие ооциты, о чем сообщалось ранее (32). Количественно влияние этого фактора было сходно с таковым для IGF1.
Немало экспериментальных данных, полученных на различных видах животных, свидетельствуют о положительном влиянии IGF1 и FGF2 в среде экстракорпорального созревания ооцитов на их последующую способность к эмбриональному развитию in vitro (11-13, 32). Наше исследование не было исключением: оба цитокина повышали выход бластоцист (см. табл.), а также их качество (см. рис. 4), когда ооциты подвергались ЭКО сразу после периода IVM. В то же время антивозрастной эффект этих 1092
факторов в отношении ооцитов ранее не изучался, и в настоящей работе в аналогичных условиях был не так однозначен. По завершении 20-часового созревания in vitro в контроле выход бластоцист составил 21,3 %, после пролонгированного культивирования зрелых ооцитов контрольной группы в течение 12 ч он снижался до 11,9 % (р < 0,05). Наблюдалось также повышение степени апоптоза в полученных IVP бластоцистах. IGFI, но не FGF2 оказал позитивное воздействие на долю развития стареющих ооцитов до стадии бластоцисты, повысив этот показатель с 11,9 % в контроле до 23,3 % (р < 0,05), а также улучшил их качество, снизив долю ядер с признаками апоптоза с 40,8 % в контроле до 23,2 %. Очевидно, что антивозрастной эффект IGFI, оказываемый на созревающие ооциты, имеет более выраженный и долговременный характер, чем в случае FGF2.
Следует отметить, что, несмотря на значимость проблемы старения ооцитов в условиях in vitro, исследований, посвященных поиску веществ, которые способны замедлять возрастные изменения в яйцеклетках, пока немного. Сообщается, что воздействие L-карнитина на коровьи ооциты может замедлять процессы их старения (25). Добавление этого антиоксиданта в среду IVM, кроме снижения окислительного стресса в зрелых ооцитах, увеличивало, как и в нашем исследовании при добавлении IGFI, долю их развития до стадии бластоцисты после ЭКО (с 20 до 29 %), а также снижало степень апоптоза в эмбрионах. Внеклеточные везикулы из фолликулярной жидкости яичников в среде IVM и флавоноидный гликозид растительного происхождения икариин в среде старения могут также повышать резистентность ооцитов к возрастным изменениям и улучшать развитие эмбрионов in vitro.
Таким образом, индивидуальное воздействие инсулиноподобного фактора роста 1 (IGFI) и фактора роста фибробластов 2 (FGF2) на ооциты коров во время созревания in vitro способствует повышению качества яйцеклеток, а также их способности к эмбриональному развитию после экстракорпорального оплодотворения. Первое проявляется в снижении степени апоптоза в МII хромосомах, второе — в увеличении процента развившихся из ооцитов бластоцист и подавлении в них апоптотических изменений. Для IGFI, кроме того, эти эффекты имеют долговременный характер и сохраняются после пролонгированного культивирования созревших клеток, вызывающего их старение, что указывает на повышение устойчивости ооцитов к возрастным трансформациям в присутствии указанного цитокина. Влияние FGF2 в среде IVM на возрастные изменения в созревающих ооцитах коров менее выражено и проявляется только в снижении степени апоптоза в МII хромосомах. Это свидетельствует о различии в предшествующих молекулярных изменениях при воздействии исследуемых цитокинов. Следовательно, несмотря на то, что и IGFI, и FGF2 положительно влияют на качество коровьих ооцитов, созревающих in vitro, для повышения их устойчивости к возрастным трансформациям в среду IVM следует добавлять IGF1.
