Особенности водного режима и оценка жаростойкости груши уссурийской при использовании биоудобрений

Автор: Федорова Д.Г., Галактионова Л.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 8, 2024 года.
Бесплатный доступ
Цель исследования - определение показателей засухоустойчивости и жаростойкости груши уссурийской, произрастающей в семиаридном климате, с использованием биоудобрений (на примере города Оренбурга). Засухоустойчивость растений является одним из важных факторов успешного культивирования новых таксонов в климатических условиях при наличии лимитирующих факторов. Чрезмерная высокая температура вызывает быстрое повреждение клеток или их гибель, что приводит к катастрофическим изменениям во всем организме растений. Использование определенных видов удобрений может повысить устойчивость культурных растений к различным видам стресса, включая воздействие высоких температур. Жаростойкость, общую оводненность, водный дефицит, водоудерживающую способность и уровень подвижной влаги в листовых пластинках определяли по общепринятым методикам в лабораторных условиях. Для этого осуществляли взвешивание листьев: свежесобранные, через 24 часа, после полного насыщения, абсолютно сухие. Устойчивость образцов к воздействию высоких температур оценивали с использованием водяной бани. Установлено, что у опытных растений, с внесением исследуемого удобрения значительно увеличивается степень жаростойкостьи и они принадлежат к группе высокотермоустойчивых растений. Степень засухоустойчивости также выше у опытных образцов, по сравнению с контролем. Оценив степень водного режима с использованием модифицированной методики для климата Оренбуржья, авторы делают вывод, что опытные образцы относятся к группе растений с высокой засухоустойчивостью, контрольные - к группе среднезасухоустойчивых растений.
Засухоустойчивость, жаростойкость, водный режим, груша уссурийская, биоудобрения
Короткий адрес: https://sciup.org/140307633
IDR: 140307633 | УДК: 631.671.3 | DOI: 10.36718/1819-4036-2024-8-65-72
Peculiarities of water regime and evaluation of heat resistance of Ussuri pear under biofertilizer use
The aim of the study is to determine the indicators of drought resistance and heat resistance of Ussuri pear growing in a semiarid climate using biofertilizers (using the city of Orenburg as an example). Drought resistance of plants is one of the important factors for the successful cultivation of new taxa in climatic conditions in the presence of limiting factors. Excessive high temperature causes rapid cell damage or death, which leads to catastrophic changes in the entire plant organism. The use of certain types of fertilizers can increase the resistance of cultivated plants to various types of stress, including exposure to high temperatures. Heat resistance, total water content, water deficit, water-holding capacity and the level of mobile moisture in leaf blades were determined using generally accepted methods in laboratory conditions. For this purpose, the leaves were weighed: freshly picked, after 24 hours, after complete saturation, absolutely dry. The resistance of the samples to high temperatures was assessed using a water bath. It was established that the experimental plants, with the introduction of the studied fertilizer, significantly increase their heat resistance and they belong to the group of highly heat-resistant plants. The degree of drought resistance is also higher in experimental samples, compared to the control. Having assessed the degree of water regime using a modified technique for the climate of the Orenburg Region, the authors conclude that the experimental samples belong to the group of plants with high drought resistance, the control ones - to the group of moderately drought-resistant plants.
Текст научной статьи Особенности водного режима и оценка жаростойкости груши уссурийской при использовании биоудобрений
Введение. Экстремальные изменения климата создают трудности сельскому хозяйству и продовольственной безопасности. Стресс от высокой температуры ограничивает рост и продуктивность, а в экстремальных условиях приводит к гибели растений. Высокие температуры влияют на урожайность и качество плодовых культур. В отличие от термочувствительных термотолерантные растения обладают хорошими системами противостояния тепловому стрессу. Физические изменения и преобразования метаболизма, включая изменение ориентации листьев, снижение потерь воды, изменение липидного состава мембран и увеличение размеров сосудов ксилемы, являются общими механизмами ответа растений на воздействие высоких температур [1, 2]. В свою очередь, повышение температурного режима индуцирует выработку активных форм кислорода, что вызывает перекисное окисление липидов и пигментов, приводящее к потере проницаемости мембраны [3]. Кроме того, стресс от засухи изменяет состав хлоропластов и метаболитов листьев, тем самым снижая скорость фотосинтеза и приводя к укорочению жизненного цикла и снижению продуктивности растений [4].
Чрезмерная высокая температура вызывает быстрое повреждение клеток или их гибель, что приводит к катастрофическим изменениям во всем организме [2, 5]. Транскриптомный и про-теомный анализы показали, что высокая температура индуцирует различные физиологические реакции, связанные с биотическим и абиотическим стрессом в растениях. Такие физиологические реакции включают липидный и вторичный метаболизм, передачу сигналов кальция, фосфорилирование белка, передачу сигналов фитогормонов, метаболизм РНК и регуляцию транскрипции [6–10, 11].
Кроме того, различные реакции растений могут повышать их устойчивость к стрессу от засухи, включая закрытие устьиц, накопление пролина и индукцию других защитных механизмов [12, 13]. Например, Shourbalal et al. (2019) обнаружили, что можно изменять фенологические параметры и физиологические процессы растения пшеницы в условиях стресса от засухи, регулируя яровизацию растений с помощью регуляторов роста растений, включая гиббереллиновую кислоту, кинетин и 6-бензиладенин [14].
Засухоустойчивость растений можно разделить на четыре основных типа – предотвращение засухи, засухоустойчивость, избегание засухи и восстановление после засухи. В качестве индикаторов для оценки засухоустойчивости растений используются такие признаки, как особенности развития корней, листьев, способность к осмотической адаптации, параметры водного режима и стабильность клеточной мембраны [15].
До настоящего времени использовались различные методы смягчения воздействия высоких температур и низкой влажности на рост растений, включая использование засухоустойчивых сортов растений, стимулирование роста растений обработкой корневых систем ризобак-териями, биофизические воздействия и применение удобрений [16].
Исследования показали, что избирательное использование определенных видов удобрений может повысить устойчивость культурных растений к различным видам стресса, включая воздействие высоких температур [14, 17]. Элементы питания могут влиять на физиологию и морфологию растений в зависимости от его биохимических свойств. Так, например, калий выполняет важные функции в растениях, включая повышение устойчивости растений к абиотическим стрессовым факторам [18, 19].
Цель исследования – изучение влияния внесения биоудобрений на особенности жаро- и засухоустойчивости груши уссурийской ( Pyrus ussuriensis Maxim. ex Rupr), произрастающей в климатических условиях Оренбуржья.
Объекты и методы. Исследования проводились в течение вегетационного периода 2023 г. на территории Ботанического сада ФГБОУ ВО Оренбургский государственный университет. Почвенный покров участка исследования был представлен черноземом обыкновенным среднегумусным среднемощным тяжелосуглинистым, на котором произрастали деревья груши уссурийской.
При проведении эксперимента использовалось удобрение, созданное на основе растительных отходов переработки сельскохозяйственной продукции с добавлением биологически активных компонентов и содержанием органических веществ и азота не менее 75 и 4,5 % соответственно. Удобрение вносилось в осенний период (2022 г.) в дозе 5 л на 1 м2. Повторно, в аналогичной дозировке, удобрения вносили в почву ранней весной (2023 г.).
Объект исследования – груша уссурийская. Для каждого варианта опыта было исследовано 3–5 деревьев (выбор деревьев основывался исключительно по отбору деревьев с неповрежденными листьями):
№ 1 – образцы опытные (с внесением удобрения);
-
№ 2 – контрольные образцы (без удобрения).
Степень термоустойчивости растений исследовалась нами в засушливую погоду, когда наиболее активно работают механизмы адаптации к данному неблагоприятному фактору.
Жаростойкость оценивали, используя общепринятую методику Ф.Ф Мацкова [20], с применением водяной бани UT-4300E. При этом оценивали степень побурения листьев при дозированной тепловой нагрузке. Использовались следующие диапазоны температур: 40; 50; 60 °С. После тепловой обработки листья охлаждали и помещали в 0,1 Н раствор соляной кислоты на 10 мин. Изменение зеленой окраски листьев на бурую объясняется преобразованием хлорофилла в феофетин при повреждении мембран клеток. Степень поражения листовых пластинок оценивалась нами визуально, чего вполне достаточно для дифференциации объектов исследования по термоустойчивости. Непораженная часть листа оставалась зеленого цвета, а чем больше площадь побурения листовой пластинки, тем меньше жароустойчивость. Показатель степени повреждения определяли по общепринятой шкале, при этом учитывали разделение объектов исследования на 5 качественных групп [20] (табл. 1).
Таблица 1
Общепринятая шкала оценки степени повреждений
|
Качественная оценка |
Характерные признаки повреждений |
|
Очень высокая |
При 60 ºС не отмечается повреждений листьев |
|
Высокая |
При 60 ºС повреждается до 50 % площади листовой пластинки |
|
Средняя |
Переносит 50 ºС (повреждается до 80 % площади листа), гибнет при 60 ºС |
|
Низкая |
Переносит 40 ºС, гибнет при 50 ºС |
|
Очень низкая |
Гибнет при 40 ºС |
Оценку водного режима производили по следующим параметрам: общая оводненность (ОВ), водный дефицит (ВД), количество подвижной влаги (ПВ), водоудерживающая способность (ВС). Оценивали данные параметры по стандартным методикам [20, 21]. Для этого осущест- вляли взвешивание листьев: свежесобранные, через 24 ч, после полного насыщения, абсолютно сухие. Взвешивания осуществляли с использованием лабораторных весов CAS MWP-300.
Оценку засухоустойчивости проводили по следующей схеме [22] (табл. 2).
Схема оценки засухоустойчивости
Таблица 2
|
Номер группы |
Степень засухоустойчивости |
Характерные признаки |
|
1 |
Высокая |
Высокие показатели общей оводненности в совокупности с высокой водоудерживающей способностью |
|
2 |
Средняя |
Отмечается при высокой водоудерживающей способности в совокупности с низкими показателями общей оводненности, либо низкая водоудерживающая способность при высокой оводненности |
|
3 |
Низкая |
Оба параметра имеют низкие значения |
На основе проведенной статистической обработки сделан анализ о жаро- и засухоустойчивости объектов исследования. Статистическую обработку данных проводили с применением программы MS Excel и Statistica 10.0.
Результаты и их обсуждение. Погодные условия в период проведения исследований характеризуются максимальной нестабильностью (табл. 3).
Таблица 3
Метеоданные по г. Оренбургу (2023 г.) (по данным Государственного учреждения «Оренбургский областной центр по гидрометеорологии и мониторингу окружающей среды»)
|
Показатель |
Июнь |
Июль |
Август |
|
|
Температура, °С |
средн. |
+20,3 |
+23,7 |
+21,8 |
|
max |
+35,2 |
+40,4 |
+34,3 |
|
|
min |
+5,0 |
+9,8 |
+8,1 |
|
|
Сумма активных температур, °С |
621,5 |
735,9 |
655,3 |
|
|
Относительная влажность воздуха, % |
средн. |
49 |
58 |
53 |
|
min |
16 |
9 |
15 |
|
|
Скорость ветра, м/с |
средн. |
3,6 |
3,0 |
3,0 |
|
max |
10 |
8,0 |
10 |
|
|
Осадки, мм |
сумма |
31 |
67,3 |
31,6 |
|
ГТК |
0,49 |
0,86 |
0,48 |
|
Гидротермический коэффициент за период июль – август, при сумме активных температур – 2 012,7 °С, равен 0,61, что определяет погодные условия, характерные для зон «сухого земледелия». Данный факт подтверждает актуальность исследований, посвященных проблемам повышения степени термоустойчивости растительных организмов в данных климатических условиях.
Оценка жаростойкости осуществлялась в середине сезона вегетации в трехкратной повторности. Минимальная температура воздействия на листья была 40 °С. Видимых повреждений на листовых пластинках нами не отмечено, поэтому можно с уверенностью утверждать, что все исследуемые образцы выдерживают данный температурный режим.
При температуре 50 °С у 1-го варианта опыта наблюдалось небольшое побурение, которое в наибольшей степени концентрировалось у центральной жилки. Повреждения составили не более 7–10 % от площади листовой пластинки. У контрольных образцов при этой же температуре отмечено значительное повреждение, которое в среднем составило 70 % от площади листа. Максимальных значений (до 90 %) при данном температурном режиме поражение достигало у контрольных образцов.
При максимальной температуре (60 °С) у образцов с внесением удобрения повреждения были более ярко выражены в сравнении с воздействием других температурных режимов. Поражения затрагивали до 50–60 % листовой пластинки. Участки, изменившие окраску, располагались в большей степени также вдоль главной жилки листа. У контрольных образцов повреждения достигали 80 % от общей площади листьев.
Таким образом, оценив по вышеописанной методике степень жаростойкости исследуемых образцов, можно сделать вывод, что образцы листовых пластинок деревьев, растущих под влиянием удобрения, обладают высокой степенью устойчивости, когда контрольные варианты опыта – средней термоустойчивостью.
Высокие температуры сказываются на изменении параметров водного режима растений. Так, оводненность образцов с внесением удобрений в среднем за период исследования сос- тавила почти 49 %, а контрольных – 44 % при водоудерживающей способности 2,7 %. При использовании биоудобрений этот показатель увеличился в 4 раза и составил 8,66 %. Стоит отметить, что водный дефицит у опытных образцов оказался хоть и не достоверно, но меньше контроля с разницей в 4,3 %. Содержание подвижной влаги в листьях исследуемых образцов в среднем практически одинаково, с незначительной разницей всего в 1 % (рис. 1).
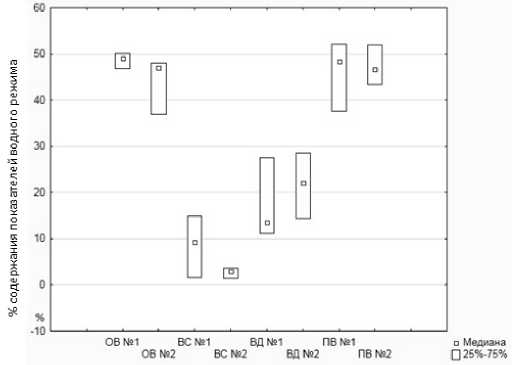
Рис. 1. Диаграмма размаха показателей водного режима исследуемых образцов
На графике (рис. 2) наглядно отображается динамика изменения исследуемых параметров в течение периода вегетации. Так, общая овод-ненность у листьев опытных образцов характеризуется наибольшей стабильностью, несмотря на изменчивость климатических факторов в период вегетации. У контрольных образцов наблюдается заметное снижение этого параметра в самый жаркий месяц (июль), а также снижение в конце летнего периода по сравнению с началом летнего сезона. Сходная тенденция прослеживается и по показателю водоудерживающей способности, однако у образцов первого варианта опыта в самый жаркий период этот параметр значительно выше, чем у контрольных растений.
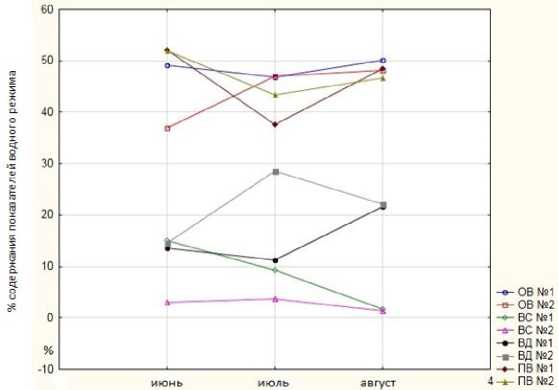
Рис. 2. Динамика показателей водного режима груши уссурийской
Характеризуется резкой противоположностью изменчивость водного дефицита у контрольного и опытного образцов. При практически одинаковых значениях в июне значения потребности в воде у образцов № 1 снижаются при увеличении засухи, у контроля – резко увеличиваются. Данный факт свидетельствует о лучшей адаптации к засушливым условиям образцов растений с внесением биоудобрения. Тенденция содержания подвижной влаги в листьях у обоих образов сходна; так, максимальное содержание подвижной влаги у всех образцов наблюдается в июне, а минимальное – в августе. Однако при воздействии высоких температур у опытных образцов наблюдается снижение уровня подвижной влаги в большей степени, чем у образцов № 2. Это означает, что образцы первого варианта в большей степени адаптированы к снижению влажности окружающей среды, уменьшая транспирацию и тем самым сохраняя водный баланс в листьях в норме.
Анализ параметров водного режима образцов груши показал, что использование биоудобрения значительно повышает степень засухоустойчивости, так как, согласно шкале, приведенной нами в методическом аппарате, опытные образцы относятся к первой группе засухоустойчивости, а контрольные – к средней.
Заключение. В ходе исследований выявлены общие тенденции динамики водных показателей в течение периода вегетации растений.
Полученные данные о вариации показателей общей оводненности, содержания подвижной формы влаги, водоудерживающей способности и водного дефицита частично являются механизмами адаптации к изменениям температуры и влажности окружающей среды.
Опытные образцы, выращиваемые с использованием биоудобрения, характеризуются более высокой засухоустойчивостью и жаростойкостью по сравнению с контрольными.
Таким образом, экспериментальные исследования с моделированием засухи и воздействия высоких температур на листовые пластины показали, что использование биоудобрения значительно повышает термоустойчивость груши уссурийской, позволяя тем самым активно использовать данное растение в насаждениях районов с семиаридным климатом.
Вестник КрасГАУ. 2024. № 8 (209) Список источников
-
1. Genetic and epigenetic control of plant heat responses / J. Liu [et al.] // Frontiers in plant science. 2015. Vol. 6. P. 267.
-
2. Molecular mechanisms governing plant responses to high temperatures / B.J. Li [et al.] // Journal of integrative plant biology. 2018. Vol. 60, № 9. P. 757–779.
-
3. Niu Y., Xiang Y. An overview of biomembrane functions in plant responses to high-temperature stress // Frontiers in plant science. 2018. Vol. 9. P. 915.
-
4. Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles / M. Djanaguiraman [et al.] // BMC plant biology. 2018. Vol. 18. P. 1–17.
-
5. Plant molecular stress responses face climate change / I. Ahuja [et al.] // Trends in plant science. 2010. Vol. 15, № 12. P. 664–674.
-
6. Overexpression of CsCaM3 improves high temperature tolerance in cucumber / B. Yu [et al.] // Frontiers in Plant Science. 2018. Vol. 9. P. 797.
-
7. Integrating omics and alternative splicing reveals insights into grape response to high temperature / J.F. Jiang [et al.] // Plant Physiology. 2017. Vol. 173. № 2. P. 1502–1518.
-
8. Transcriptional profiling and identification of heat-responsive genes in perennial ryegrass by RNA-sequencing / K.H. Wang [et al.] // Frontiers in Plant Science. 2017. Vol. 8. P. 1032.
-
9. Tissue-specific transcriptomics reveals an important role of the unfolded protein response in maintaining fertility upon heat stress in Ara-bidopsis / S.S. Zhang [et al.] // The Plant Cell. 2017. Vol. 29, № 5. P. 1007–1023.
-
10. Maize WRKY transcription factor ZmWRKY106 confers drought and heat tolerance in transgenic plants / C.T. Wang [et al.] // International journal of molecular sciences. 2018. Vol. 19. № 10. P. 3046.
-
11. Proteome and transcriptome reveal involvement of heat shock proteins and indoleacetic acid metabolism process in Lentinula edodes thermotolerance / G.Z. Wang [et al.] // Cellular Physiology and Biochemistry. 2018. Vol. 50, № 5. P. 1617–1637.
-
12. Differential response of quinoa genotypes to drought and foliage-applied H 2 O 2 in relation to oxidative damage, osmotic adjustment and antioxidant capacity / H. Iqbal [et al.] // Ecotoxicology and environmental safety. 2018. Vol. 164. P. 344–354.
-
13. Enhanced drought tolerance of foxtail millet seedlings by sulfur dioxide fumigation / Y. Han [et al.] // Ecotoxicology and environmental safety. 2019.Vol. 178. P. 9–16.
-
14. The quantity/intensity relation is affected by chemical and organic P fertilization in calcareous soils / J. Shahabifar [et al.] // Ecotoxicology and Environmental Safety. 2019. Vol. 172. P. 144–151.
-
15. Fang Y., Xiong L. General mechanisms of drought response and their application in drought resistance improvement in plants // Cellular and molecular life sciences. 2015. Vol. 72. P. 673–689.
-
16. M Sunflower ( Helianthus annuus L.) biochemical properties and seed components affected by potassium fertilization under drought conditions / S. Zamani [et al.] // Ecotoxicology and environmental safety. 2020. Vol. 190. P. 110017.
-
17. Investigating the effect of biochar on the potential of increasing cotton yield, potassium efficiency and soil environment / X. Wu [et al.] // Ecotoxicology and environmental safety. 2019. Vol. 182. P. 109451.
-
18. Wang Y., Wu W. H. Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency // Current Opinion in Plant Biology. 2015. Vol. 25. P. 46–52.
-
19. He synergistic effects of sodium and potassium on the xerophyte Apocynum venetum in response to drought stress / Y.N. Cui [et al.] // Plant physiology and biochemistry. 2019. Vol. 135. T. 489–498.
-
20. Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур / под ред. Е.Н. Седова, Т.П. Огольцовой. Орел, 1999. С. 235–246.
-
21. Таренков В.А., Иванова Л.Н. Водоудерживающая способность листьев боярышника в связи с устойчивостью к засухе // Интродукция, акклиматизация, охрана и использование растений. Куйбышев, 1990. С. 3–9.
-
22. Федорова Д.Г., Назарова Н.М., Кухлев-ская Ю.Ф. Модификация методики оценки жизнеспособности интродуцентов в соот-
ветствии с условиями сухостепной зоны Оренбургского Предуралья // Вестник Нижневартовского государственного университета. 2021. № 2 (54). С. 57–62.
Список литературы Особенности водного режима и оценка жаростойкости груши уссурийской при использовании биоудобрений
- Genetic and epigenetic control of plant heat responses / J. Liu [et al.] // Frontiers in plant science. 2015. Vol. 6. P. 267.
- Molecular mechanisms governing plant responses to high temperatures / B.J. Li [et al.] // Journal of integrative plant biology. 2018. Vol. 60, № 9. P. 757–779.
- Niu Y., Xiang Y. An overview of biomembrane functions in plant responses to high-temperature stress // Frontiers in plant science. 2018. Vol. 9. P. 915.
- Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles / M. Djanaguiraman [et al.] // BMC plant biology. 2018. Vol. 18. P. 1–17.
- Plant molecular stress responses face climate change / I. Ahuja [et al.] // Trends in plant science. 2010. Vol. 15, № 12. P. 664–674.
- Overexpression of CsCaM3 improves high temperature tolerance in cucumber / B. Yu [et al.] // Frontiers in Plant Science. 2018. Vol. 9. P. 797.
- Integrating omics and alternative splicing reveals insights into grape response to high temperature / J.F. Jiang [et al.] // Plant Physio-logy. 2017. Vol. 173. № 2. P. 1502–1518.
- Transcriptional profiling and identification of heat-responsive genes in perennial ryegrass by RNA-sequencing / K.H. Wang [et al.] // Frontiers in Plant Science. 2017. Vol. 8. P. 1032.
- Tissue-specific transcriptomics reveals an im-portant role of the unfolded protein response in maintaining fertility upon heat stress in Ara-bidopsis / S.S. Zhang [et al.] // The Plant Cell. 2017. Vol. 29, № 5. P. 1007–1023.
- Maize WRKY transcription factor ZmWRKY106 confers drought and heat tolerance in transge-nic plants / C.T. Wang [et al.] // International journal of molecular sciences. 2018. Vol. 19. № 10. P. 3046.
- Proteome and transcriptome reveal involvement of heat shock proteins and indoleacetic acid metabolism process in Lentinula edodes thermotolerance / G.Z. Wang [et al.] // Cellular Physiology and Biochemistry. 2018. Vol. 50, № 5. P. 1617–1637.
- Differential response of quinoa genotypes to drought and foliage-applied H2O2 in relation to oxidative damage, osmotic adjustment and an-tioxidant capacity / H. Iqbal [et al.] // Ecotoxi-cology and environmental safety. 2018. Vol. 164. P. 344–354.
- Enhanced drought tolerance of foxtail millet seedlings by sulfur dioxide fumigation / Y. Han [et al.] // Ecotoxicology and environmental safety. 2019.Vol. 178. P. 9–16.
- The quantity/intensity relation is affected by chemical and organic P fertilization in calca-reous soils / J. Shahabifar [et al.] // Ecotoxi-cology and Environmental Safety. 2019. Vol. 172. P. 144–151.
- Fang Y., Xiong L. General mechanisms of drought response and their application in drought resistance improvement in plants // Cellular and molecular life sciences. 2015. Vol. 72. P. 673–689.
- M Sunflower (Helianthus annuus L.) bioche-mical properties and seed components affect-ed by potassium fertilization under drought conditions / S. Zamani [et al.] // Ecotoxicology and environmental safety. 2020. Vol. 190. P. 110017.
- Investigating the effect of biochar on the po-tential of increasing cotton yield, potassium ef-ficiency and soil environment / X. Wu [et al.] // Ecotoxicology and environmental safety. 2019. Vol. 182. P. 109451.
- Wang Y., Wu W. H. Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency // Current Opinion in Plant Biology. 2015. Vol. 25. P. 46–52.
- He synergistic effects of sodium and potas-sium on the xerophyte Apocynum venetum in response to drought stress / Y.N. Cui [et al.] // Plant physiology and biochemistry. 2019. Vol. 135. T. 489–498.
- Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур / под ред. Е.Н. Седова, Т.П. Огольцовой. Орел, 1999. С. 235–246.
- Таренков В.А., Иванова Л.Н. Водоудерживающая способность листьев боярышника в связи с устойчивостью к засухе // Интродукция, акклиматизация, охрана и использование растений. Куйбышев, 1990. С. 3–9.
- Федорова Д.Г., Назарова Н.М., Кухлев-ская Ю.Ф. Модификация методики оценки жизнеспособности интродуцентов в соответствии с условиями сухостепной зоны Оренбургского Предуралья // Вестник Ниж-невартовского государственного университета. 2021. № 2 (54). С. 57–62.
