Особенности водного режима некоторых видов растений в тропическом лесу на юге Вьетнама во влажный сезон

Автор: Евдокимова Елена Валентиновна, Новичонок Артем Олегович, Марковская Евгения Федоровна, Курбатова Юлия Александровна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 4 (125), 2012 года.
Бесплатный доступ
Рассматриваются основные параметры водного обмена (интенсивность транспирации, водный дефицит, насыщающее содержание воды) 6 видов растений подлеска и подроста муссонного тропического леса во влажный сезон. Выделены 2 группы видов: первая, хорошо адаптированная, характеризуется относительной стабильностью водного режима; вторая - не имеет стабильного водного режима в связи с конкуренцией с другими видами этого сообщества.
Транспирация, водный дефицит, насыщающее содержание воды, подлесок, подрост, муссонный тропический лес
Короткий адрес: https://sciup.org/14750148
IDR: 14750148 | УДК: 581.116
Water status of some plant species in tropical forest of South Vietnam during wet season
Basic parameters of the water balance (the rate of transpiration, water deficit, and saturating water content), of the six understory and young undergrowth species of the tropical monsoonal forest during wet seasons were studied. We defined two groups of species. The first well-adapted group is characterized by the stability of the water balance, while the second group has unstable water status due to the competition with other species of this plant association.
Текст научной статьи Особенности водного режима некоторых видов растений в тропическом лесу на юге Вьетнама во влажный сезон
Древостой влажного тропического леса имеет сложную структуру. В вертикальном строении прослеживаются несколько подъярусов, что приводит к сильной трансформации факторов внешней среды. Прежде всего это касается потоков солнечной радиации, а также температуры и влажности воздуха.
Наиболее характерной экологической чертой растений подлеска в тропическом лесу являются адаптационные особенности, связанные с существованием в условиях слабой освещенности, – изменения в содержании пигментов и СО2-газообмена [2]. Кроме недостатка света лимитирующую роль в развитии растений играет избыточное содержание влаги в почве во влажный период. Избыток влаги в почве вызывает изменение устьичной проводимости [15] и приводит к снижению ассимиляции углерода [16], [17]. Вопрос о влиянии лимитирующих факторов на развитие растений остается в настоящее время по-прежнему дискуссионным. На основании большого системного исследования, проведенного С. Н. Шереметьевым на широком диапазоне градиента влажности почв, был сделан вывод, что нет местообитаний, которые принято называть стрессовыми. Виды в каждом местообитании полностью соответствуют эко- логической и фитоценотической обстановке [12]. Выводы были получены для травяного покрова. Исследование не включало зону влажных тропических лесов.
Цель настоящего исследования состояла в оценке основных параметров водного обмена растений сезонно-влажного тропического леса, произрастающих в условиях избыточного увлажнения, что можно рассматривать как вариант гумидных условий.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование было проведено в Национальном парке Кат Тиен, расположенном на юге Вьетнама (11° с. ш, 107° в. д.). Климат на территории национального парка относится к мягкому муссонному тропическому типу [13]. Среднегодовая температура воздуха составляет 25,4 °С, колебания между самыми жаркими и самыми прохладными месяцами не превышают 3–4 °С. В то же время суточная амплитуда температуры воздуха может составлять более 10 °С. Во время влажного сезона (май – октябрь) выпадает до 90 % годовой нормы осадков (~ 2000 мм) [13]. В вертикальном строении древостоя парка прослеживается от трех до пяти ярусов. В качестве объектов исследования были выбраны 6 видов
растений разных экологических групп, имеющих разные стратегии развития и произрастающих на данной стадии онтогенетического развития в нижнем ярусе сезонно-влажного тропического леса: Lagerstremia calyculata Kurz – подрост доминанта верхнего яруса, Hibiscus sp. , Dalbergia sp. , – подрост второго-третьего ярусов, Phyllan-thus sp. – вид нижнего яруса, Ancistrocladus tec-torius Lour. (Merr.), Calamus sp. – лианы.
Исследования были выполнены на 5–7 растениях каждого вида. Lagerstremia calyculata , Hibiscus sp. и Dalbergia sp. находились на вирги-нильном этапе развития, Phyllanthus sp. , Ancistro-cladus tectorius , Calamus sp. – на генеративном. Для определения интенсивности транспирации с каждого растения со средней части кроны отбирали по 2–5 листьев; для определения водного дефицита и степени оводненности – по 5 листьев для одной пробы. Все показатели определяли в 3–5-кратной повторности. Интенсивность транспирации листьев определяли методом быстрого взвешивания [5]. Для этого измеряли массу листьев сразу после сбора и через 3 минуты. Интенсивность транспирации ( E ) рассчитывали по формуле:
E = (Mo – M3 ) ⋅ 20/S (г/м2 ⋅ ч), где Mo и M3 – масса листьев сразу после сбора и через 3 минуты соответственно, S – площадь листьев, 20 – коэффициент пересчета минут в часы.
Одновременно с измерением транспирации собирали листья для определения дефицита водного насыщения и насыщающего содержания воды. Сразу после сбора измеряли свежую массу листьев. Для определения массы листьев при насыщении листья регидрировали во влажном поролоне в течение 24 часов. Затем листья высушивали в сушильном шкафу при 60 оС до постоянной массы и определяли сухую массу листьев. Дефицит водного насыщения ( WSD ) и насыщающее содержание воды ( WCs ) в листе рассчитывали по формулам:
WSD = (Ws – Wf ) / (Ws – Wd) ⋅ 100 (%), WCs = Ws – Wf / Wd (г/г), где Wf и Wd – свежая и сухая масса листьев, Ws – масса листьев при насыщении.
Исследования были проведены в июле – августе 2011 года в дневное время суток (6–18 ч.) с интервалом измерений в один час. Транспирационные циклы для каждого растения проводились в 3-кратной повторности.
Параллельно с измерением транспирации регистрировали метеорологические параметры: освещенность, влажность и температуру воздуха. Пространственная изменчивость метеорологических параметров, особенно освещенности, под пологом леса высокая, поэтому их регистрация проводилась непосредственно в месте измерения транспирации. Освещенность измеряли с помощью люксметра «ТКА-ЛЮКС». Для измерения температуры использовали цифровой термометр «Checktemp 1», относительной влажности воздуха – цифровую метеостанцию «Anymetre JR 900».
Статистическую обработку полученных результатов проводили с использованием программы Statgraphics 2.1.
РЕЗУЛЬТАТЫ
Основными экологическими факторами, оказывающими влияние на водный обмен растений, являются интенсивность фотосинтетически активной радиации, температура и относительная влажность воздуха [7]. Измерение освещенности ( J ) в период проведения исследований (влажный сезон) показало, что она имеет четкий суточный ход и достигает максимальных значений в 11 часов утра (рис. 1), восход солнца происходит в 05:40, заход – в 18:15 (1 августа). Максимальные значения относительной влажности воздуха ( H ) отмечаются в утренние и вечерние часы (рис. 1). Минимальная влажность воздуха на высоте 1 м наблюдается в 13 часов. Изменение влажности воздуха в течение суток незначительное и составляет 7–8 %. Перепад температуры воздуха ( T ) в течение суток составляет 3–4 °С (рис. 1). Интенсивность транспирации
Анализ дневной динамики интенсивности транспирации исследуемых видов растений показал увеличение скорости транспирации с вос-
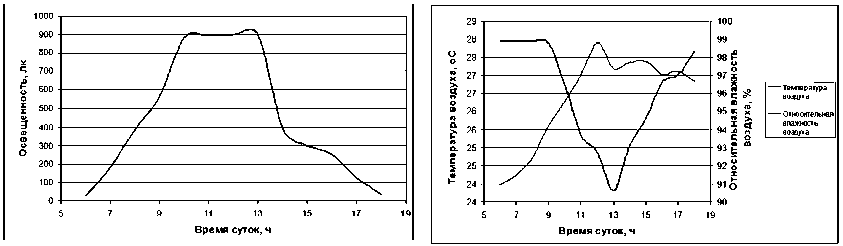
Рис. 1. Средняя суточная динамика освещенности, температуры и влажности воздуха под пологом леса на высоте 1 м в июле – сентябре 2011 года (Вьетнам, Национальный парк Кат Тиен)
ходом солнца. При этом у растений Ancistrocla-dus tectorius и Phyllanthus sp. почти сразу после восхода солнца (6 часов утра) отмечался высокий уровень транспирации, достигающий почти 80 % от максимума. Максимальная интенсивность транспирации (Emax) у растений Ancis-trocladus tectorius и Phyllanthus sp. наблюдалась в 11–13 и 9–12 часов соответственно (рис. 1). Сопоставление дневного хода интенсивности транспирации растений Phyllanthus sp. и освещенности показало несовпадение максимумов их фаз. Наибольшая интенсивность транспирации наблюдалась на 2 часа раньше, чем максимум освещенности.
Корреляционный анализ показал сильное влияние освещенности, температуры и относительной влажности воздуха на суточный ритм транспирации Ancistrocladus tectorius (табл. 1) и отсутствие зависимости между скоростью транспирации у Phyllanthus sp. и факторами внешней среды. Анализ дневной динамики скорости транспирации растений Dalbergia sp.
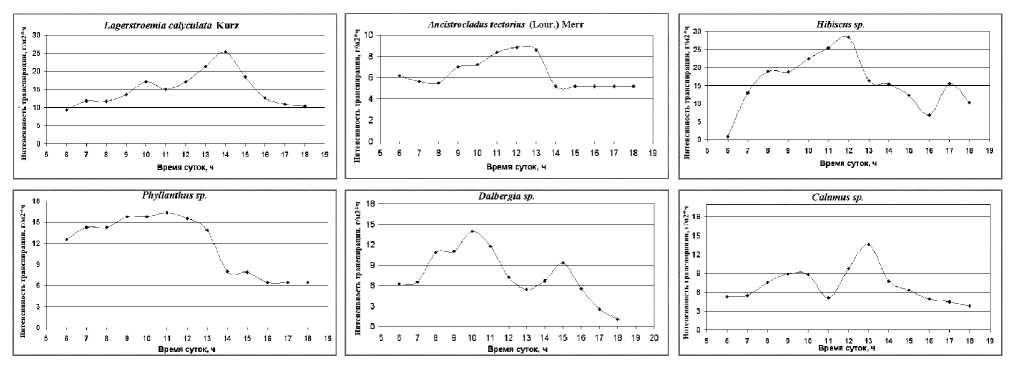
Рис. 2. Средняя суточная динамика интенсивности транспирации растений в июле – сентябре 2011 года (Вьетнам, Национальный парк Кат Тиен)
Таблица 1
Зависимость между интенсивностью транспирации и экологическими факторами
|
Вид |
Экологический фактор |
Коэффициент корреляции (r2) |
Коэффициент детерминации (R2) |
p-Value |
|
Ancistrocladus tectorius |
Температура воздуха |
0,66 |
43,20 |
0,0030 |
|
Относительная влажность воздуха |
-0,71 |
50,04 |
0,0010 |
|
|
Освещенность |
0,75 |
56,49 |
0,0003 |
|
|
Phyllanthus sp . |
Температура воздуха |
0,15 |
2,18 |
0,36 |
|
Относительная влажность воздуха |
-0,22 |
4,83 |
0,17 |
|
|
Освещенность |
0,31 |
9,54 |
0,05 |
|
|
Dalbergia sp. |
Температура воздуха |
0,003 |
0,0007 |
0,99 |
|
Относительная влажность воздуха |
0,01 |
0,02 |
0,97 |
|
|
Освещенность |
0,83 |
69,5 |
0,0004 |
|
|
Calamus sp. |
Температура воздуха |
0,30 |
4,4 |
0,24 |
|
Относительная влажность воздуха |
-0,26 |
6,9 |
0,15 |
|
|
Освещенность |
0,34 |
11,96 |
0,15 |
|
|
Hibiscus sp. |
Температура воздуха |
0,33 |
11,14 |
0,27 |
|
Относительная влажность воздуха |
-0,33 |
11,08 |
0,27 |
|
|
Освещенность |
0,68 |
46,85 |
0,0099 |
|
|
Lagerstremia calyculata |
Температура воздуха |
0,65 |
41,9 |
0,0152 |
|
Относительная влажность воздуха |
-0,75 |
56,9 |
0,0032 |
|
|
Освещенность |
0,77 |
59,05 |
0,0021 |
и Calamus sp . показал, что у этих видов также отмечаются высокие значения транспирации утром сразу после восхода солнца, но они ниже и составляют около 40 % от суточного максимума. Вечером, после захода солнца, транспирация у Calamus sp. еще держится на относительно высоком уровне, а у Dalbergia sp. снижается до нуля. Кривые дневного хода транспирации у этих видов имеют два пика максимума, которые приходятся на разное время (у Calamus sp. на 9– 10 и 13 ч., а у Dalbergia sp. на 10 и 15 ч.). При этом стоит отметить, что у Calamus sp. не было выявлено зависимости между интенсивностью транспирации и факторами среды, а на транспирацию Dalbergia sp. оказывает влияние только освещенность (табл. 1). Из данных видов по интенсивности транспирации выделяются растения Phyllanthus sp. , которые показали максимальные значения – 16,3 г/м2ч, средние – 13,5 г/м2ч. У остальных видов данной группы максимальные и средние значения скорости транспирации ниже и соответствуют: Calamus sp. – 14,0 и 7,1 г/м2ч; Ancistrocladus tectorius – 9,0 и 6,9 г/м2ч; Dalber-gia sp. – 14,0 и 7,5 г/м2ч.
Существуют некоторые особенности дневной динамики транспирации у Hibiscus sp . Суточная кривая транспирации имеет один высокий пик в 12 ч. и небольшое увеличение в 17 ч. Интенсивность транспирации у Hibiscus sp. наибольшая из всех изученных видов: максимальные – 28,3 г/м2ч, средние – 15,7 г/м2ч. Для этого вида выявлена довольно сильная корреляционная связь между скоростью транспирации и освещенностью (табл. 1).
Суточный ход транспирации у подроста La-gerstroemia calyculata имеет один максимум в 14 ч. Уже с восходом солнца отмечаются высокие значения транспирации, которые после достижения максимума начинают снижаться, но и после захода солнца остаются на уровне, сходном с утренними значениями. Максимальные значения транспирации у Lagerstroemia calycu-lata достигают 25,3 г/м2ч, среднее значение – 14,9 г/м2ч. Наиболее сильное влияние на суточный ход транспирации Lagerstremia calyculata оказывают освещенность и влажность воздуха (табл. 1).
Дефицит водного насыщения
Дневная динамика дефицита водного насыщения у разных видов была сходной: наиболее низкие значения водного дефицита наблюдали в утренние и вечерние часы. В дневное время с понижением влажности воздуха и увеличением температуры водный дефицит увеличивался. Время формирования максимального водного дефицита различается у разных видов и варьирует между 11 и 13 ч. (табл. 2). Диапазон дефицита водного насыщения у всех изученных видов изменялся в пределах от 5 до 35 %. У Phyllanthus sp. , Lagerstroemia calyculata , Ancistrocladus sp.
и Calamus sp. водный дефицит практически не формируется, и его величина составляет от 5 до 12–15 % (табл. 2). Для двух видов, являющихся подростом второго-третьего ярусов ( Dalbergia sp. , Hibiscus sp. ), отмечены высокие значения водного дефицита. У Hibiscus sp. наблюдаются наибольшие по сравнению с остальными изученными видами значения водного дефицита (36 %).
Таблица 2
Средние ( WSD ) и максимальные ( WSDmax ) значения водного дефицита листьев растений сезонно-влажного тропического леса во влажный период
|
Вид |
WSD , % |
WSDmax , % |
Время наступления WSDmax , часы |
|
Hibiscus sp. |
20,6 ± 2,57 |
36,3 |
12 |
|
Lagerstroemia calyculata |
8,0 ± 0,68 |
12,2 |
12 |
|
Phyllanthus sp. |
4,2 ± 0,38 |
5,6 |
11 |
|
Ancistrocladus tectorius |
10,8 ± 1,05 |
14,4 |
13 |
|
Calamus sp. |
5,2 ± 0,55 |
8,2 |
13 |
|
Dalbergia sp. |
19,2 ± 1,64 |
24,4 |
13 |
Насыщающее содержание воды
Максимальные значения насыщающего содержания воды наблюдаются у Phyllanthus sp. (3,4 г/г) и Hibiscus sp. (2,7 г/г), а минимальные – у Calamus sp . (1,00 г/г). У остальных видов ( Ancis-trocladus tectorius , Dalbergia sp. и Lagerstroemia calyculata ) значения насыщающего содержания воды находятся примерно на одном уровне – около 2 г/г (табл. 3). По этому показателю не выявлено отличий между разными группами растений. Значения насыщающего содержания воды зависят от многих факторов, в том числе от биологических особенностей видов и, в частности, от анатомо-морфологической структуры.
Таблица 3
Средние значения насыщающего содержания воды в листьях растений сезонно-влажного тропического леса во влажный период
|
Вид |
Насыщающее содержание воды, г /г |
|
Hibiscus sp. |
2,71 ± 0,09 |
|
Lagerstroemia calyculata |
2,20 ± 0,04 |
|
Phyllanthus sp. |
3,39 ± 0,06 |
|
Ancistrocladus tectorius |
1,90 ± 0,10 |
|
Calamus sp. |
1,00 ± 0,04 |
|
Dalbergia sp. |
2,00 ± 0,14 |
ОБСУЖДЕНИЕ
В работе исследованы виды, которые различаются по жизненной стратегии: Phyllanthus sp. является типичным видом подлеска и постоянно растет в условиях очень низкой освещенности и высокой влажности воздуха и почвы, особенно во влажный сезон. Три других древесных вида
( Lagerstroemia calyculata , Hibiscus sp. , Dalber-gia sp. ) произрастают в этих условиях только на ранних этапах онтогенеза, а затем они должны достичь верхнего ( Lagerstroemia calyculata ) или второго-третьего ( Hibiscus sp. , Dalbergia sp. ) ярусов. Лианы – Ancistrocladus tectorius и Calamus sp. могут достигать второго-третьего и верхнего ярусов соответственно.
Проведенная работа показала, что исследуемые виды включают виды с высокими – 25–30 ( Lagerstroemia calyculata , Hibiscus sp. ), средними – 14–16 ( Phyllanthus sp. , Dalbergia sp. , Calamus sp. ) и низкими – 9 г/м2ч ( Ancistrocladus tectorius ) значениями Emax . Эти данные поддерживают представления C. Н. Шереметьева о том, что в гумидных местообитаниях сосуществуют виды с разной скоростью транспирации при слабом экотопическом отборе, но при более конкурентных отношениях между видами [12].
Высокие значения водного дефицита были обнаружены у Hibiscus sp. и Dalbergia sp. Самые низкие значения водного дефицита были получены у Phyllanthus sp. и Calamus sp. Сильное варьирование показателя оводненности листьев в пределах одного типа леса на небольшой территории также согласуется с представлениями С. Н. Шереметьева о том, что в гумидных экото-пах снижается сходство видов и увеличивается разнообразие [12].
Вопрос о связи дневных колебаний интенсивности транспирации с факторами среды активно изучается, но имеющиеся данные достаточно противоречивы. В ряде работ показана тесная связь скорости транспирации с температурой [6], [11] и другими факторами [9]. Однако имеются работы, в которых не отмечается связи между транспирацией и факторами среды [3], [4]. Высказывается мнение, что связь между дневным ходом транспирации и гидрометеорологическими параметрами обусловлена степенью увлажнения экотопа: при достаточном увлажнении эта связь есть, а при недостатке она нарушается [1], [7], [10]. Во влажный период в сезонно-влажном лесу растения произрастают в условиях высокой влажности воздуха и почвы. Однако даже в этих условиях выделяются три группы видов. Так, у Ancistrocladus tectorius и Lagerstroemia calycu-lata скорость транспирации зависит от всех изученных факторов среды. Однако у Phyllanthus sp. и Calamus sp. нет корреляции транспирации ни с одним из факторов среды. Эти виды имеют такие механизмы внутренней регуляции, которые, по-видимому, успешно работают и в условиях благоприятного увлажнения. У Dalbergia sp. и Hibiscus sp. отмечается зависимость транспирации только от освещенности, что может свидетельствовать о ведущей роли этого фактора в коррекции водного режима в условиях подлеска.
В литературе широко обсуждается вопрос о форме дневных колебаний транспирации . По мнению С. Н. Шереметьева, для растений гу-мидной зоны характерна одновершинная кривая с максимумом в околополуденное время, а для аридных условий наблюдаются многовершинные кривые [12]. В полученных нами данных имеются как одно-, так и двувершинные кривые. Последние получены у Calamus sp. и Dalbergia sp. – двух видов, которые различаются между собой по другим показателям водного обмена. По гипотезе Г. Ш. Нахуцришвили [9], двувер-шинность кривых суточного хода транспирации может свидетельствовать об активном регулировании водного режима указаннами видами. Этот тезис хорошо согласуется с данными по Calamus sp. – виду, интенсивность транспирации которого не зависит от факторов среды.
Проведенное исследование показало, что типичные для подлеска виды, такие как Ancis-trocladus tectorius , Phyllanthus sp. , хорошо адаптированы к условиям среды и занимают определенную экологическую нишу в подлеске. Они различаются по интенсивности водного режима: у Phyllanthus sp. сравнительно высокая интенсивность транспирации, отсутствует водный дефицит и максимальные значения насыщающего содержания воды, а у Ancistrocladus tectorius низкая скорость транспирации, отсутствует водный дефицит и средние значения насыщающего содержания воды. От этих видов отличаются два других вида подлеска, которые имеют двувершинные кривые суточного ритма скорости транспирации. Динамика этого процесса слабо связана с факторами среды: у Dalbergia sp. сравнительно высокие значения транспирации, присутствует водный дефицит, средние значения насыщающего содержания воды, у Calamus sp. сравнительно высокие значения транспирации, не формируется водный дефицит, наиболее низкие значения насыщающего содержания воды. Эти параметры водного обмена свидетельству-. ют о менее благоприятных для этих двух видов условиях произрастания. Что касается Hibiscus sp. , то высокая скорость транспирации, хорошо синхронизированная с динамикой освещенности, способна компенсировать высокие значения водного дефицита, которые отмечаются у этого вида. Это может свидетельствовать о высокой конкурентоспособности вида в борьбе за освещенность, что соответствует и его положению в подлеске. Особого внимания заслуживает подрост Lagerstroemia calyculata – растения, для которых характерны максимальные значения транспирации при отсутствии водного дефицита. Растения хорошо адаптированы к условиям произрастания, в том числе к световым, и не участвуют в конкуренции за свет. Высокий уровень зависимости от температуры и влажности воздуха подтверждает их высокий уровень адаптации.
Таким образом, в системе подлеска выявлены две группы видов. Одна группа (Ancistrocla-dus tectorius, Phyllanthus sp., Lagerstroemia ca-lyculata) хорошо адаптирована к этим условиям и занимает определенную экологическую нишу, что обеспечивает относительную стабильность водного режима. Виды второй группы (Dalber-gia sp., Hibiscus sp., Calamus sp.) не имеют стабильного водного режима в связи с их участием в конкурентных взаимоотношениях с другими видами этого растительного сообщества.
Список литературы Особенности водного режима некоторых видов растений в тропическом лесу на юге Вьетнама во влажный сезон
- Алексеенко Л. Н. Водный режим луговых растений в связи с условиями среды. Л.: ЛГУ, 1976. 198 с.
- Вальтер Г. Растительность земного шара: Пер. с нем. Т. 1: Тропические и субтропические зоны. М.: Прогресс, 1968. 550 с.
- Витко К. Р. Экологическая характеристика гырнецовой дубравы в южной Молдавии: Автореф. дис.... канд. биол. наук. Кишинев, 1963. 18 с.
- Горшкова А. А. Эколого-морфологические особенности и водный режим степных растений Забайкалья: Автореф. дис.... д-ра биол. наук. Новосибирск, 1970. 57 с.
- Иванов Л. А., Силина А. А., Цельникер Ю. Л. О методе быстрого взвешивания для определения транспирации в естественных условиях//Ботанический журнал. 1950. Т. 35. № 2. С. 171-185.
- Иванов Л. А., Силина А. А., Цельникер Ю. Л. О транспирации полезащитных полос в условиях Деркульской степи//Ботанический журнал. 1953. Т. 38. № 2. С. 167-184.
- Копытова Л. Д. Запас воды в степных сообществах Забайкалья и ее расход на транспирацию: Автореф. дис.... канд. биол. наук. Иркутск, 1974. 22 с.
- Лархер В. Экология растений: Пер. с нем. М.: Мир, 1978. 384 с.
- Нахуцришвили Г. Ш. Экология высокогорных травянистых растений и фитоценозов Центрального Кавказа: Автореф. дис.... д-ра биол. наук. Тбилиси, 1972. 52 с.
- Савинкин А. П. Водный режим пустынных пастбищ и богарных посевов в подзоне бурых почв Казахстана: Автореф. дис.... канд. биол. наук. Алма-Ата, 1962. 28 с.
- Свешникова В. М. Водный режим растений и почв высокогорных пустынь Памира//Труды Института ботаники АН Таджикской ССР. Душанбе, 1962.
- Шереметьев С. Н. Травы на градиенте влажности почвы (водный обмен и структурно-функциональная организация). М.: Товарищество научных изданий КМК, 2005. 271 с.
- Blanc L., Maury-Lechon G., Pascal J.-P. Structure, floristic composition and natural regeneration in the forests of Cat Tien National Park, Vietnam: an analysis of the successional trends//Journal of biogeography. 2000. Vol. 27 (1). P 141-157.
- Ehleringer J. R., Field C. B. Scaling physiological processes: Leaf to Globe. San Diego: Academic Press, 1993. 388 p.
- Herrera A., Tezara W., Marin O., Rengifo E. Stomatal and non-stomatal limitations of photosynthesis in trees of a tropical seasonally flooded forest//Physiologia Plantarum. 2008. Vol. 134 (1). P. 41-48.
- Kozlowski T. T. Responses of woody plants to flooding and salinity//Tree Physiology Monograph. 1997. Vol. 1. P. 1-29.
- Kreuzwieser J., Papadopoulou E., Rennenberg H. Interaction of flooding with carbon metabolism of forest trees//Plant Biology. 2004. Vol. 6 (3). P. 299-306.
