Отбор в гетерогенных популяциях сои: источники вариации (сообщение 1)
")
Автор: Розенцвейг В.Е., Голоенко Д.В., Давыденко О.Г.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (166), 2016 года.
Бесплатный доступ
Изучалась структура дисперсионного комплекса продуктивности растений сои для определения направления дальнейших исследований по минимизации селекционно -бесполезных компонентов вариации. Вариация продуктивности изучалась на модельных популяциях, представляющих смеси сортов 00-000 групп спелости и имитирующих разнообразие реальных гибридных популяций, при двух плотностях стеблестоя: 40 и 20 раст./м2 Наследуемость составляла 10-13 %, что говорит о неэффективности прямого отбора по продуктивности. В гетерогенных популяциях наблюдалась смена рангов генотипов по сравнению с однородными популяциями тех же сортов, обусловленная генотипической конкуренцией; варианса взаимодействия была равна 14-17 %. Наибольший вклад в дисперсию вносила экологическая конкуренция, варианса которой составляла 70-76 %. В разреженном ценозе селекционно-полезная и генотипическая кокурентная вариансы несколько увеличивались, тогда как экологическая конкурентная - снижалась. Таким образом, снижение основной компоненты селекционнобесполезной вариации - внутригенотипической дисперсии - будет более эффективным в разреженном ценозе, но при этом необходим анализ генотипических конкурентных отношений.
Селекция сои, структура вариансы продуктивности, бесполезные компоненты, конкурентная варианса
Короткий адрес: https://sciup.org/142151300
IDR: 142151300 | УДК: 631.523.4:633.853.52
Selection in heterogeneous soybean populations: 1. Variation sources
Soybean seed yield variation complex was studied in order to define further research directions aimed at minimizing of variation components useless for breeding. Individual plant seed yield variation was studied on model populations consisting of 000-00 maturity group cultivars and simulating diversity in real hybrid populations, at two plant populations of 40 and 20 plants per sq. m. Heritability was equal to 10-13%; thus, direct selection for seed yield is ineffective. Genotype ranks changed in heterogeneous populations compared to the same cultivars grown in pure line populations due to genotypic competition; interaction variance was equal to 14-17%. Ecological competition reaching 70-76% contributed most into the overall variance. In low plant population, genetically useful and genotypic competitive variances were increased a little while ecological competitive variance decreased. Thus, decreasing of the main variance component useless for breeding, i.e. intragenotypic variance, should be more effective at low plant population; however, analysis of intergenotypic competition is required.
Текст научной статьи Отбор в гетерогенных популяциях сои: источники вариации (сообщение 1)
Введение. В популяциях растений селекционно - полезная генетическая дисперсия маскируется рядом средовых «шумов», которые могут значительно превышать ее. Фенотипическую дисперсию можно разложить на следующие основные компоненты [4 ; 6; 16]:
σ = σ + σ + σ + σ + σеп (1), где σ – генотипическая селекционно полезная;
σ – генотипическая конкурентная;
σ – онтогенетическая (обусловленная различиями вегетационных периодов и соотношением фенофаз);
σ – экологическая конкурентная ;
σеп – экологическая почвенная (пара-типическая) вариансы.
Генотипическая конкурентная вариан-са обусловлена различиями генотипов по конкурентоспособности, которые не идентифицируются в однородном ценозе, но проявляются в гетерогенном. По различным оценкам [5 ; 6], она может превышать селекционно - полезную диспер сию в 3–4 раза. При наличии значимой σ невозможно корректно определить наследуемость, поскольку генотипическая конкуренция вызывает взаимодействие генотипа и среды (ВГС), выражающееся в смене рангов генотипов в однородной и гетерогенной популяциях. Предлагалось оценивать σ путем учета продуктивности растений, корневые системы которых изолированы в трубках [5 ; 15], но эта методика связана с большими трудозатратами и не устраняет конкуренции надземных частей растений.
Необходимо также устранение различий по конкурентоспособности, обусловленных вегетационным периодом [6]. Для этого можно: а) проводить отбор раздельно в пределах узких групп спелости; б) использовать вместо абсолютной урожайности «относительную», т.е. отнесенную к продолжительности вегетационного периода; или в) величину отклонения от регрессии урожайности на вегетационный период.
Экологическая конкурентная варианса существует и в однородной популяции. Она вызывается неравномерностью заделки (и, как следствие, всходов) и разно-качественностью посевного материала: различиями по выполненности, количеству запасных веществ и энергии прораста- ния семян. В силу неравенства начальных условий, растения идентичного генотипа, вступая в автоконкуренцию, осваивают неодинаковую площадь питания. По существующим оценкам, σ может достигать 50 и даже 70–83 % [8; 15] σ . Предложено оценивать эту вариансу путем подсева части семян позже основного срока сева [8]. Однако этот способ неприемлем для сои, поскольку манипулирование сроком сева вызывает ВГС посредством фотопериодической реакции, зависящей от генотипа. В среднем каждый день задержки посева сои вызывает снижение продуктивности на 0,5 %, высоты – на 0,6 см, сокращение вегетационного периода – на 0,3 сут. [20].
Экологическую конкуренцию можно снизить путем точного высева и снижения плотности стеблестоя [11], однако последнее должно приводить к повышению генотипической конкуренции из - за различий генотипов по способности осваивать площадь питания за счет соседей. Эффекты ВГС вызывают при этом смену рангов, в результате чего могут получать преимущество морфотипы, неспособные эффективно использовать ограниченные ресурсы в производственном посеве [12: с. 169]. Таким образом, представляется необходимым установить, при какой схеме посева гибридных питомников сумма селекционно - бесполезных варианс будет минимизирована.
Паратипическая варианса, обусловленная пестротой плодородия почвы, может составлять порядка 20 % σ [10] и устраняется увеличением размера делянки и числа повторностей. Поскольку отбор в гибридных популяциях проводится среди ближайшего окружения, то этот источник варьирования не рассматривается в настоящей работе. Оптимизация площади делянки и числа повторностей рассмотрена в работе Бойко [2].
Наибольшие затруднения для селекционера представляет отбор элитных растений в гибридной популяции, поскольку на этом этапе селекции эффекты генотипа искажаются всеми средовыми «помехами», а наследуемость, судя по приведенным оценкам их уровней, составляет порядка 10 %. Целью настоящего исследования было изучение структуры вариации продуктивности в гетерогенных популяциях сои и оптимизация строения агроценоза для повышения эффективности отбора.
Материалы и методы. Для изучения структуры вариации и конкурентных процессов в гибридных питомниках сои нами были сформированы модельные популяции , представляющие собой смеси сортов 000 и ранней 00 групп спелости в равных пропорциях. Структура выборки сортов была близка к послойной [13], отражая разнообразие типов роста и ветвления, структуры продуктивности, аллелей фотопериодизма, которое может наблюдаться в реальных гибридных популяциях. Выборку производили в коллекционном питомнике, принимая во внимание рекомендации ряда авторов оценивать ВГС и индекс среды по разнообразным генотипам, а не по линиям КСИ, которые уже отселектированы для данных условий [14; 18; 21]. В учет были включены сорта, близкие по сроку созревания (113 ± 1 сут., CV g = 3 %). Для имитации реального генетического разнообразия в сортосмесь добавляли около 20 % генотипов 0 группы, которые не учитывались («браковались по позднеспелости») при уборке.
Модельные популяции выращивались при двух плотностях посева (20 и 40 раст./м 2 ) в трех повторностях. Опыты были заложены в 2010 и 2011 гг. в Луни-нецком р - не Беларуси, в 2012, 2014 и 2015 гг. – в Лельчицком р - не (обе точки на широте 52 о , с супесчаной почвой). При уборке растения разных сортов идентифицировались по апробационным признакам, убирались и анализировались отдельно. Этот же набор сортов параллельно оценивался на 3- рядных делянках в 5– 6 повторностях, при рекомендованных в производстве плотностях 40– 50 раст./м 2 .
Изложенный подход позволяет устранить взаимодействие «генотип–год», которое накладывается на результаты оценки элитных растений по потомству в реальных гибридных популяциях, когда отбор и испытание потомств проходят в разные годы.
Эффекты генотипов и их зависимость от типа популяции (однородная / гетерогенная) и ее плотности оценивались путем дисперсионного анализа. Из трёх повторностей модельной (гетерогенной) популяции и деляночного опыта (однородная популяция) отбиралось по 10 случайных растений каждого сорта. Эффекты фактора «тип популяции» считались фиксированными, а его градации сравнивались попарно: однородная 40 раст./м 2 с гетерогенной 40 раст./м 2 ; однородная 40 раст./м 2 с гетерогенной 20 раст./м 2 .
Эффекты генотипов принимались случайными, в соответствии с послойной случайной выборкой. Кроме того, согласно Драгавцеву [4], генотипическая конкурентная варианса о| с0m не аддитивна к о | , а включается в нее за счет ВГС. Поскольку в модели со случайными эффектами вари-анса взаимодействия включается в дисперсии факторов (а при фиксированных эффектах – нет), то случайность эффектов генотипов в данном случае более соответствует биологической природе варьирования. Таким образом, дисперсионный анализ проводился по смешанной модели III : эффекты плотностей фиксированные, эффекты генотипов случайные (табл. 1).
Таблица 1
Смешанная модель дисперсионного анализа «генотип – тип популяции»
|
Источник вариации |
Структура MS |
Интерпретация σ 2 |
|
Популяция |
о е + псо р |
– |
|
Генотип |
о е + со | хр + mco | |
= О |и |
|
Взаимодействие |
О е + СО | хр |
= о | со m |
|
Случайная |
σ е |
= о е со m |
|
Общая |
_ 2 — ° р Һ |
Примечание: n – число генотипов ; m – популяций, c – растений в выборке
Для устранения эффекта шкалы, возникающего вследствие различной площади питания, масса семян с растения в дисперсионном анализе была выражена в процентах (%) от центральной средней в каждом из типов популяции.
Результаты и обсуждение. Структура дисперсионного комплекса. Продуктивность генотипов в трех типах популяций приведена в таблице 2. При сопоставлении урожайности сортов и их средней продуктивности (массы семян с растения) в гетерогенных популяциях наблюдалась смена рангов, обусловленная различиями по конкурентоспособности.
Таблица 2
Продуктивность генотипов в однородном ценозе и модельных популяциях (средние значения за 2010-2015 гг.)
|
Сорт |
Урожайность, % 1 |
Средняя продуктивность в гетерогенной популяции, % от центральной средней |
|
|
40 раст./м 2 |
20 раст./м 2 |
||
|
Припять |
105 |
103 |
93 |
|
Оресса |
1 0 0 |
108 |
1 12 |
|
Грациja |
98 |
96 |
11 1 |
|
McCa l l |
96 |
130 |
1 6 6 |
|
Щара |
92 |
93 |
82 |
|
Gentleman |
90 |
109 |
120 |
|
Золотиста |
89 |
101 |
85 |
|
Major |
86 |
1 3 3 |
1 2 2 |
|
Бiлявка |
85 |
75 |
64 |
|
Снежок |
81 |
9 9 |
120 |
|
OT 94-47 |
75 |
1 17 |
128 |
|
Соер 5 |
72 |
5 5 |
52 |
|
Малахит |
63 |
50 |
47 |
Примечание: 1 – 3-рядковые делянки в 5–6 повтор- ностях. 100 % = 2,52 т/га
Компоненты вариации приведены в таблице 3. Эффекты популяций отсутствовали, поскольку продуктивность растений была выражена в процентах (%) от центральной средней по варианту опыта (а не в г/раст.; см. выше).
Анализ дисперсионного комплекса продемонстрировал следующее. Селекционно-полезная генетическая дисперсия σ и наследуемость Н2 (= ηgu) были несколько выше при разреженном стеблестое. Но в целом, очевидно, что прямой отбор на продуктивность при таком уровне наследуемости неэффективен. Вариан-са взаимодействия, отражающая смену рангов генотипов вследствие различий по конкурентоспособности, была в среднем в 1,2 раза выше (на 3 абс. %) в разреженном ценозе, а экологическая конкуренция снизилась в 1,09 раза (на 6 абс. %) благодаря увеличенной площади питания. σ превышала σ в 1,3–1,4 раза, σ пре вышала σ в 5–7 раз. Таким образом, снижение внутригенотипической вариан-сы σ , как составляющей основную часть бесполезной дисперсии, представляется первоочередной задачей, решение которой необходимо для повышения эффективности отбора.
Таблица 3
Доли влияния п компонент вариации (%)
|
Плотность популяции |
Год |
Индекс среды I j , т/га 1 |
Факторы и вариансы |
||
|
генотип ( σ ) |
взаимодействие ( σ ) |
случайная (σ ) |
|||
|
40 раст./м 2 |
2010 |
3,2 |
12 * |
21 * * |
66 |
|
201 1 |
2,4 |
2 |
18 * * |
80 |
|
|
2012 |
1,9 |
1 1 * * |
13 * * |
76 |
|
|
2014 |
3,1 |
17 * * |
12 * * |
71 |
|
|
2015 |
1,7 |
9 * * |
4 * |
87 |
|
|
Среднее |
2,5 |
10 |
14 |
76 |
|
|
20 раст./м 2 |
2010 |
3,2 |
13 |
33 * * |
55 |
|
201 1 |
2,4 |
1 1 * |
8 * * |
81 |
|
|
2012 |
1,9 |
15 * * |
15 * * |
70 |
|
|
2014 |
3,1 |
17 * * |
17 * * |
66 |
|
|
2015 |
1,7 |
8 * * |
10 * * |
82 |
|
|
Среднее |
2,5 |
13 |
17 |
70 |
|
1 – Средняя урожайность в деляночном опыте с повторностями
* – Средний квадрат фактора значим на уровне α = 0,05
** – Средний квадрат фактора значим на уровне α = 0,001
Хотя параметр наследуемости подвергался обоснованной критике [16 ; 19], но, при всей его относительности и ограничениях, его можно считать применимым для сопоставления данных в пределах одного эксперимента.
Hawtin [18] отмечает, что при отборе в гибридных популяциях сои в Уганде наследуемость (в широком смысле) была выше при разреженном стеблестое (10 раст./м 2 ) по сравнению с плотным (40 раст./м 2 ), и, хо- 15
тя ее значение оказалось завышенным, больший генетический сдвиг наблюдался все же при отборе в разреженном ценозе, который, таким образом, являлся лучшим дифференцирующим фоном. При анализе данных сортоиспытания в США [17] было показано, что у сои и пшеницы σ и Н 2 положительно коррелировали с индексом среды, тогда как у ячменя, овса и масличного льна такой связи не было. Иными словами, продуктивные среды обладали в среднем лучшей дифференцирующей способностью для сои.
Межгенотипическая конкуренция.
Кроме дисперсионного анализа, о | с от можно оценить с помощью смены рангов генотипов при их выращивании в однородном ценозе и в сортосмеси (см. табл. 2):
о|сот = о| . (1 — RgXp), где а 1 - генетическая варианса в широком смысле' т-е- а1" + °|сот.
Корреляция значений урожайности генотипов в многорядковых делянках с повторностями и их средней продуктивности в гетерогенной популяции с плотностью 40 раст./м 2 была равна 0,57; аналогичная корреляция для гетерогенной популяции с плотностью 20 раст./м 2 – 0,47 (5- летние данные). Отсюда :
– для гетерогенной популяции 40 раст./м 2 П 6 сот = 24 % . (1 — 0,57 2 ) = 16 %;
– для гетерогенной популяции 20 раст./м 2 n gc0т = 30 % . (1 - 0,47 2 ) = 23 %.
Эти значения и их соотношение близки к полученным из дисперсионных анализов (см. табл. 3): при 20 раст./м2 генетическая конкуренция и обусловленная ею смена рангов возрастают.
Внутригенотипическая конкуренция.
Для иллюстрации снижения Оес от в разреженном стеблестое приведем распреде ления продуктивности растений в однородных популяциях при 40 и 20 раст./М (рис. 1). Они были близки к асимметричному распределению Максвелла, которое используется для описания конкуренции в ценозе [9]. В 16
разреженном посеве растения были более выровненными: эксцесс распределения возрастал, частота класса угнетенных растений уменьшалась, CV e продуктивности снизился в среднем в 1,15 раза (с 41 до 35 %), а^ о т - в 1,16 раза (20102011 гг.). Можно предположить следующие механизмы снижения селекционно бесполезных компонент фенотипической дисперсии в разреженном посеве: п | сот снижается, так как позже наступает дефицитность и исчерпание ресурсов, а оеп - так как редкий посев более засухоустойчив [1 ; 3].
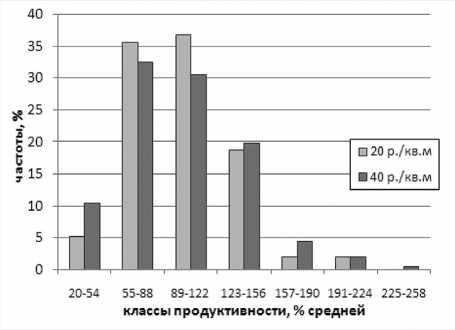
Рисунок 1 – Распределение продуктивности (% от средней по сорту) растений из однородных популяций с плотностью 40 и 20 раст./м 2 (объединенные данные по 4 сортам за 2010–2011 гг.).
Для сортов сои 0-II групп спелости было показано [3], что снижение плотности стеблестоя с 20 до 5 раст./м 2 снижает CV e продуктивности растений с 38–49 до 24 –30 % и, таким образом, позволяет частично устранить неэффективные отборы, обусловленные Ое сот , однако при этом не оценивалось изменение о| сот , которое характеризует ВГС.
Коэффициент вариации был использован для устранения эффекта шкалы, так как продуктивность при большей площади питания возрастает. Однако необходимо принять во внимание существующее возражение: снижение CVе может быть обусловлено тем, что он обратно пропорционален х [7]. В этом случае график σ / х не проходит через начало координат. В регрессионной модели σ / х, построенной путем объединения 83 выборок по 13 генотипам за 5 лет, свободный член был равен 0,26 ± 0,28 (α = 0,35), а значимая отрицательная корреляция между CVе и х отсутствовала (R2 = 0,014).
Выводы. 1. В гетерогенных популяциях сои прямой отбор по продуктивности неэффективен. Наследуемость составляет 10 – 13 %, варианса генотипической конкуренции – 14 – 17 %, варианса экологической конкуренции – 70 – 76 %.
-
2. При пониженной плотности популяции селекционно - полезная и генотипическая конкурентная вариансы несколько увеличиваются, экологическая конкурентная снижается.
Список литературы Отбор в гетерогенных популяциях сои: источники вариации (сообщение 1)
- Баранов В.Ф. Агрономические аспекты повышения засухоустойчивости ценозов сои//Повышение продуктивности сои: Сб. науч. тр. -Краснодар, 2000. -С. 71-76.
- Бойко Ю.Г. Оптимизация методов испытаний генотипов и проведения отборов из популяций подсолнечника: автореф. дис.. канд. с.-х. наук. -Краснодар, 2009. -24 с.
- Гужов Ю.Л., Патирана Р. Закономерности изменчивости количественных признаков у сои и использование их в селекции//Сельскохозйственная биология. -1980. -Т.15.-№ 6.-С. 854-859.
- Драгавцев В.А. Новые принципы отбора генотипов по количественным признакам в селекции растений//Генетика количественных признаков сельскохозяйственных растений. -М.: Наука, 1978. -С. 5-9.
- Драгавцев В.А., Герасименко И.И. Об эффективности идентификации генотипов по фенотипам при визуальном отборе пшеницы в F2 по признаку «масса зерна с главного ко лоса»//Проблемы отбора и оценки селекционного материала. -Киев: Наукова думка, 1980.-С. 29-31.
- Драгавцев В.А., Шкель Н.М., Герасименко И.И., Дьяков А.Б. О подходах к оценке генетического потенциала урожайности яровых пшениц Западной Сибири//Сельскохозяйственная биология. -1980. -Т. 15. -№ 2. -С. 254-263.
- Дьяков А.Б. Влияние условий внешней среды на генотипическую и экологическую изменчивость продуктивности подсолнечника//Взаимодействие генотип -среда у растений и его роль в селекции (сб.). -Краснодар, 1988.-С. 61-93.
- Дьяков А.Б., Драгавцев В.А. Конкурентоспособность растений в связи с селекцией. I. Надежность оценки генотипов по фенотипам и способ ее повышения//Генетика. -1975. -Т. 11. -№ 5. -С. 11-22.
- Лакин В.Ф. Биометрия. -М.: Высшая школа, 1990. -352 с.
- Литун П.П. Приемы уменьшения фенотипической изменчивости и ее компонентов на разных этапах отбора в селекции//Генетика количественных признаков сельскохозяйственных растений. -М.: Наука, 1978.-С. 93-100.
- Мережко А.Ф., Капешинский A.M. Сотовый (гексагональный) способ размещения растений в селекционных посевах//Селекция и семеноводство. -1984, № 1. -С. 12-15.
- Селекция сортов сои северного экотипа/Под ред. А.П. Устюжанина, В.Е. Шевченко . -Воронеж-Белгород, 2007. -225 с.
- Снедекор Дж. У. Статистические методы в применении к исследованиям в сельском хозяйстве и биологии. -М., 1961. -503 с.
- Хотылева Л.В., Тарутина Л.А. Взаимодействие генотипа и среды. -Минск: Наука и техника, 1978. -109 с.
- Чекалин Н.М. Основные направления селекции зернобобовых культур//Сельскохозяйственная биология. -1979. -Т. 14. -№ 6. -С. 695-703.
- Шкель Н.М., Драгавцев B.A. Проблема идентификации генотипов растений по фенотипам и повышение эффективности селекционного процесса//Генетический анализ количественных признаков растений. -Уфа, 1979.-С. 40-48.
- Allen F.L., Comstock R.E., Rasmusson D.C. Optimal environments for yield testing//Crop Sci. -1978. -V. 18. -P. 747-751.
- Hawtin G.C. The exploitation of genetic variation in Glycine max (L.) Merrill: Ph.D. thesis. -Cambridge, 1974. -249 p.
- Johnson H.W., Robinson H.F., Comstock R.E. Estimates of genetic and environmental variability in soybeans//Agron. J. -1955. -V. 47. -P.314-318.
- OMAFRA -Ontario Ministry for Agriculture and Food. Soybean Production in Ontario. -. -URL: http://www.omafra.gov.on.ca/english/crops/-2011.
- Pedersen A.R., Everson E.H., Grafius J.E. The gene pool concept as a basis for cultivar selection and recommendation//Crop Sci. -1978. -V. 18. -P. 883-886.
