Paleoenvironmental Conditions of Neanderthal Habitation in the Altai: Chagyrskaya and Okladnikov Caves

Author: Serdyuk N.V., Zazhigin V.S., Markin S.V., Kolobova K.A.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Paleoenvironment, the stone age
Article in issue: 1 т.50, 2022.
Free access
We explore the environments of the Sibiryachikha Neanderthals, who had migrated to the Altai at the end of MIS 4. Given that the territory was already populated by Denisovans, the key question is whether the choice of habitat was random (i.e., the immigrants occupied vacant ecological niches) or motivated by other factors. On the basis of published results relating to the study of small-mammal fauna and pollen analysis, the environments of Chagyrskaya and Okladnikov Caves during the Neanderthal habitation are reconstructed. Species of small mammals are viewed as biome members. To reconstruct the episodic transfer of mammalian remains between stratigraphic units, we used ordination statistics and compared the results with those of micromorphological and stratigraphic analyses of Chagyrskaya Cave. It was found that late Neanderthals of the Altai lived in similar environments, dominated by steppe and forest steppe landscapes. The choice of caves for habitation depended on several factors, the key ones being the availability of game and high-quality raw material for manufacturing tools. On the basis of the statistical analysis of small-mammal fauna and the stratigraphic and micromorphological analyses, we conclude that post-sedimentation processes in caves can include vertical transfer of animal remains, affecting environmental reconstructions.
Altai, Paleolithic, statistical analysis, small mammals, environmental reconstruction
Short address: https://sciup.org/145146839
IDR: 145146839 | DOI: 10.17746/1563-0110.2022.50.1.014-028
Text of the scientific article Paleoenvironmental Conditions of Neanderthal Habitation in the Altai: Chagyrskaya and Okladnikov Caves
In recent years, it has been proven that there were several waves of Neanderthal migration to the Altai in the Pleistocene. The late European Neanderthals migrated to the area at the end of MIS 4 and probably did not contact with first-wave Neanderthals. The late European Neanderthals practiced original material culture, which underwent almost no changes during the transcontinental migration (Slon et al., 2018;
Kolobova et al., 2020b; Vernot et al., 2021). New evidence suggests that the second-wave Neanderthals arrived to the territory inhabited by Denisovans, and occupied a certain ecological niche for about 20,000 years. In this context, issues concerning the subsistence strategies of new hominins in the settled area are of great relevance. Of particular importance are the issues of adaptation of the late European Neanderthals to the mosaic landscape and paleoecological conditions of the Altai Mountains.
Today, only two cave sites belonging to the Sibiryachikha techno-complex, which is the easternmost manifestation of the Micoquian, are known: Chagyrskaya and Okladnikov (Derevianko et al., 2013). Paleoecological reconstructions have already been made on the basis of the data on the Neanderthal environmental conditions in Chagyrskaya Cave; such data on the population from Okladnikov Cave are scarce.
This paper proposes comprehensive paleoreconstructions of the habitats of the late Altai Neanderthals from both caves, based on the data on small-mammal faunal communities and published results of biological analyses. Both caves are located in the low Altai Mountains. The same altitudinal belts are characterized by similar vegetation and faunal complexes, suggesting exploitation of the same range of natural resources by the late Neanderthals. Small mammals, in turn, reflect the paleoenvironments in the immediate vicinity of the sites; this provides the good ground for direct correlations between the sites under study.
The new results, on the one hand, complement the published data on the pollen analysis and the large fauna of Chagyrskaya Cave (Rudaya et al., 2017), but, on the other hand, reveal certain contradictions. For example, according to pollen analysis, landscapes near Chagyrskaya Cave, during the accumulation of layers 5 and 6, were characterized by formation of the steppe ecozone, while during the accumulation of layer 7, they were dominated by taiga. However, reconstructions based on theriological data do not confirm this. The ambiguity of the conclusions derived from various biological methods in the study of Chagyrskaya Cave requires a complex interpretation; so we involved data from detailed stratigraphic and micro-morphological analyses of the soft sediments of Chagyrskaya Cave, which showed the complexity of sedimentation and post-depositional processes in karst cavities, undoubtedly affecting the results of employed biological methods.
Material and methods
The osteological collection of Chagyrskaya Cave has been obtained using traditional methods. After being washed on sieves and dried, the bone remains were picked out from the small rock fraction. The material was identified with the aid of an MBS-10 binocular magnifying glass, using a micrometer eyepiece. For comparisons, collections of small mammals from the Borissiak Paleontological Institute of the Russian Academy of Sciences and the Zoological Museum of the Lomonosov Moscow State University were used. The derived data were processed in Excel. When analyzing the faunal composition, the percentage ratio for each species in the layer of the total amount of identifiable remains was determined; the minimum number of individuals was not determined, because it had been previously shown that when using these two techniques, the quantitative ratio between species in the layer is constant (Ivleva, 1990). In order to compare the fossil communities of small mammals from the two caves, ecological groups were identified: inhabitants of the open spaces— steppes, forest-steppes, semi-deserts; and inhabitants of closed spaces—forests; inter-zonal, semi-aquatic, and mountain-steppe species. The bats were not considered in this analysis by N.G. Ivleva. In order to make a correct comparison, they were also not taken into account in the paleoreconstructions.
The composition of small-mammal communities from various stratigraphic units was studied using multivariate ordination methods. In cases where a significant number of variables were analyzed across multiple samples, non-metric multidimensional scaling (NMDS) was applied. In order to compare paleoecological conditions, biome compositions by biotope were determined through discriminant analysis (LDA). All statistical tests were performed in the PAST software (Hammer, Harper, Ryan, 2001).
Description of sites
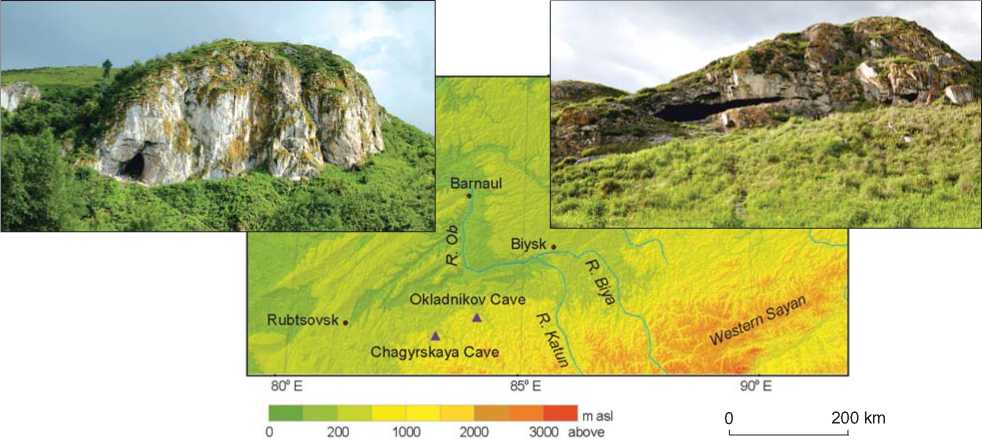
Chagyrskaya Cave is situated on the left bank of the Charysh River, in the the Tigirek Ridge branch, in the northwestern Altai (Fig. 1, 1 ). The cave faces north and is located 353 m above sea level and 19 m above the river. The cave is a key-site of the Sibiryachikha variant in the Altai Middle Paleolithic, which also includes Okladnikov Cave (Kolobova et al., 2019a). Chagyrskaya Cave is
Okladnikov Cave
Chagyrskaya Cave
ЭСТЕ
80“E
85“ E
200 km
Barnaul
Biysk
Rubtsovsk
3000 above
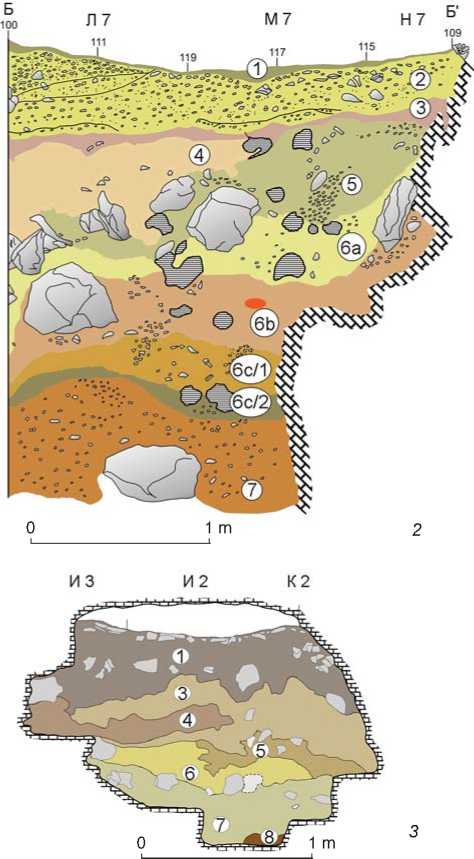
Fig. 1. Location ( 1 ) and stratigraphy of Chagyrskaya ( 2 ) and Okladnikov ( 3 ) Caves.
interpreted as a long-term camp of the Neanderthals for the consumption of hunting prey (Kolobova et al., 2019b). The hunting objects were bison and horse, which probably constituted the basis of the protein diet of the Neanderthals (Salazar-García et al., 2021). The artifacts found in the cave represent a practically complete cycle of lithic processing (Shalagina et al., 2020), as well as a complete chain of manufacturing various bone tools (Baumann et al., 2020); among which bone retouchers prevail (Kolobova et al., 2020a: 89).
Optically stimulated luminescence (OSL) dating technique, with measurement of single feldspar grains, allowed us to establish that the Neanderthals lived in the cave (layers 5–6d) for a relatively short period ranging from 59 to 49 ka BP. Layer 7, which shows no traces of ancient human habitation, is dated to 314 ka BP (Kolobova et al., 2020b).
Several stratigraphic units of loose sediments have been established at Chagyrskaya Cave. The layers are numbered from top to bottom of the profile, from the youngest to the oldest. The stratigraphy (Fig. 1, 2 ) was described by M. Krajcarz, and is based on the published data (Kolobova et al., 2019a, 2020b). The stratigraphic sequence of Chagyrskaya Cave consists of eight layers, where layer 8 is red clay accumulated in depressions in the bed rock. Layers 1 to 4 have been attributed to the Holocene, layer 5 and below to the Pleistocene.
Layer 1 is gray and dark gray non-carbonate loamy sand, slightly compacted, with considerable inclusions of small river-pebbles. This layer is the top of layer 2, altered as a result of organic-matter influx and human trampling.
Layer 2 is yellowish-brown loamy sand, similar to layer 3; it contains large quantities of pebble material. Solifluction was the main sedimentation process.
Layer 3 is grayish-brown loamy sand with a large quantity of river-pebbles. Pebbles and sand were most likely shifted into the cavity through karst chimneys in the ceiling of the back chamber from ancient riverterraces on the slope above the cave by colluvial processes. Abundant Bronze Age artifacts and the remains of fireplaces in this layer indicate the cultural features of the deposits. Numerous rodent holes were noted during excavation; the holes contain sediments from layer 3, penetrating the underlying deposits (layers 4, 5) and the upper part of layer 6a. The sediments in the holes and the soft deposits of the layers containing these holes were excavated separately.
Layer 4 is a variation of layer 5a, with a more grayish color.
Layer 5 is yellowish carbonaceous silt-deposits. This is a complex of strata comprising two types of deposit: 5a is a colluvial filling consisting of loesslike silt, with occasional rounded pebbles and angular limestone rubble, deposited in erosional channels; 5b is coarse limestone debris, usually without loose filling, which indicates very rapid accumulation. Type 5b sediments are the result of seismic events preceded by intense mechanical weathering (frost impact).
Layers 6b and 6a are brown and orange carbonaceous silt with occasional coarse limestone fragments, bone fragments, stone artifacts, and river-pebbles. The layers were established during archaeological excavations in 2007–2015. These sediments form a complex colluvial series of more than two intervening sedimentary units. These should rather be regarded as lithological types within this series. Type 6a is more clayey, orange, similar to layer 7. Type 6b is more silty, slightly denser, less porous, grayish brown, and similar to sub-layer 6c/1. The lower boundary of the series is erosional.
Layer 6c is gray carbonaceous silty loam with occasional small fluvial rounded pebbles, numerous bone fragments, Middle Paleolithic artifacts, and few limestone fragments. The sub-layer has a complex structure, and can be subdivided into two smaller units, 6c/1 and 6c/2. The lower subdivision 6c/2 is loess-like sediment with traces of pedogenesis. The layer is cryoturbated. Cryoturbation occurred after the deposition of layer 6a (i.e. much later than occupation of the cave by the Neanderthals). The deposits of 6c and underlying layers were not mixed with those of the overlying layers. Sub-layer 6c/2 contains the greatest amount of remains of the Neanderthal material culture in Chagyrskaya Cave.
Layer 6d is reddish-brown loam with fine weathered limestone debris, few bones, and small river-pebbles. It contains sediments from layer 7 mixed with soil from layer 6c redeposited vertically due to permafrost processes.
Layer 7 is red-brown clayey loam with quartz grains and small, chemically weathered fragments of limestone and river-pebbles. The lenses of greenish silt have been noted. The red clay is typical residual sediment (terra rossa type) accumulated as a result of limestone-karst dissolution.
Remains of the Neanderthal material culture have been noted in the bottom of layer 5, as well as in layers 6a, 6b, 6c/1, and 6c/2. According to stratigraphic and micromorphological analyses, layers 6c/1 and 6c/2 are undisturbed. In the other layers, archaeological material has been redeposited as a result of colluvium shifting from sub-layers 6c/1 and 6c/2 in the rear part of the cave (Kolobova et al., 2020b).
Okladnikov Cave is located in the Anui River valley, on the left bank of the Sibiryachikha River (see Figure 1, 1 ). It is located at an altitude of 368 m above sea level, and is a karst cavity with a system of branching galleries, grottoes, and chambers; stratigraphic sequences are specific for each area (Derevianko, Markin, 1992). All radiometric dates available for Okladnikov Cave (including dates of the youngest layer 1) are in the age range 37– 44 ka BP, which corresponds to the Karga interstadial, or to MIS 3 according to the SPECMAP scale (Imbrie et al., 1984) and the refined stack scale from low latitude oceanic wells ODP 677+MD900963 (Bassinot et al., 1994).
The site was discovered by A.P. Derevianko and V.I. Molodin in 1984, and has been excavated by A.P. Derevianko and S.V. Markin for four years. The archaeological material was associated with stratigraphic units 1–3, 6, 7.
Nine layers have been identified in the cave, but the layers are discontinuous and have not been established in all galleries and chambers (Fig. 1, 3). Layer 1 is loam saturated with limestone debris and solitary shale and sandstone pieces. Layer 2 under the rock-shelter includes single fragments of limestone, in gallery 2 fine debris. Layer 3 is brownish gray loam with varied clastic material in different parts of the cave. Layer 4 contains solitary rounded fragments of limestone. Layer 5 contains large fragments of limestone. Layer 6 is dark brown loam; it was noted only in the grotto. Layer 7 is reddish brown loam with weathered and soft shale and sandstone. This layer contains rounded pieces of limestone, which were possibly transported with water during the sediment formation. Layer 8 consists of reddish-yellow clays; in the galleries it contains coarse pieces of shale, sandstone, and small quartz fragments. Layer 9 is a thin stratified stratum of sandy loam recorded over a small area under a rock-shelter.
There are no data on post-depositional processes in the cave, as no detailed stratigraphic and micromorphological studies have been carried out there.
Both caves were inhabited during the period from the late MIS 4 to the early MIS 3 by the Neanderthals genetically close to late European Neanderthals (Veront et al., 2021). This Neanderthal population produced the Micoquian lithic industry characterized by radial and orthogonal core reduction and plano-convex bifaces. The toolkit is dominated by side-scrapers including simple and convergent forms, retouched points, plano-convex bifacial side-scrapers, backed knives (Keilmesser), and truncated-faceted tools. The Neanderthals from both caves used almost identical sets of raw material (about 25 types), of which jasperoids of the Zasurye Formation were of the highest quality— they were used in manufacturing formal tools of the plano-convex biface type and convergent scrapers (Derevianko et al., 2015; Shalagina, Krivoshapkin, Kolobova, 2015).
Previous studies of Chagyrskaya and Okladnikov Caves make it possible to reconstruct the landscapes in this part of the Altai during the Pleistocene.
The Chagyrskaya fossil faunal collection, including bones of large (Vasiliev, 2013) and small mammals (Derevianko et al., 2013), has been analyzed; pollen analysis has been conducted (Rudaya et al., 2017). On the basis of the derived information, a detailed reconstruction of Pleistocene environments was proposed. Layer 5 was accumulated under the conditions of the aridization of climate; forest and forest-steppe stations with tundra elements prevailed. Judging by the reduction in the number of forest species and the increase in forest-steppe and steppe species, layer 6a was transitional between layers 5 and 6b, the latter being dominated exclusively by steppe taxons. Layer 6c was formed in an environment dominated by open biotopes, although arboreal vegetation occurred during the early periods of its accumulation. All subdivisions of layer 6 correspond to a moderately arid climate. The accumulation of layer 7 is associated with the spread of periglacial landscapes. Cold and severe climate is reconstructed for this layer.
The pollen, theriological, and malacological analytical data suggest the following paleoreconstruction for Okladnikov Cave. Layers 4 and 5 were not considered in the reconstruction, as they produce no biological objects. Layer 1 was formed in a humid and cold climate, with foreststeppe zones predominating among the landscapes. Layer 2 was characterized by a drier and warmer climate than during the formation of layer 1, and mountain-steppe and forest-steppe stations were common. For layer 3, warm and dry conditions were common, with steppe dominating everywhere. Layer 6 was characterized by steppe with forbs-wormwood vegetation, and the climate was dry and warm. Layer 7 was accumulated during the spread of mixed-grassmeadow steppe and forest-steppe ecozones under humid and warm climatic conditions.
Results
Over several field seasons (2008, 2009, 2015– 2019, 2021), a rich collection of bones of the small vertebrates of Chagyrskaya Cave was assembled. In total, over 14,000 bone elements were identified, of which over 8000 fossils were identifiable as to genus and species. The osteological remains discovered in 2015–2021 do not contradict the previous data; they complement and confirm the assumptions made before.
Bones from Chagyrskaya Cave show varying degrees of preservation. Holocene remains are white or cream-colored, Pleistocene remains are light yellow and light brown. Among the Pleistocene finds, isolated light-colored bones and teeth occur; this was noted by S.K. Vasiliev (2013). Some of the bones show traces of gastric juice, indicating that it was included in the taphocoenosis composition from the pellets.
38 small mammals belonging to four orders (Chiroptera, Lipotiphla, Lagomorpha, and Rodentia) were identified up to species. In general, the composition of the Chagyrskaya small-mammal faunal remains is similar to that of the fossil faunal assemblages from other sites in the northwestern Altai.
A half of the small-mammal bones were recovered from the Holocene layers. Among the Pleistocene deposits, the greatest amount of bone remains were associated with layers 5 and 6a. Layers 6b and 6c
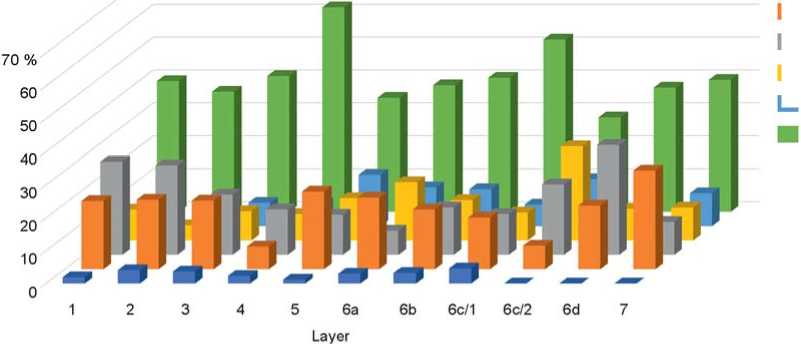
Fig. 2. Ratio of ecological groups of small mammals of Chagyrskaya Cave by layers. 1 – semi-aquatic; 2 – mountain-steppe; 3 – forest; 4 – steppe; 5 – semi-desert; 6 – forest-steppe.
produced ten times less bones, which may be due to the periodicity of human habitation in the cave. Layers 6b and 6c yielded the greatest amount of stone tools and Neanderthal fossils (Vernot et al., 2021). The large-mammal fossil collections show differences in the taphonomy of these layers (Vasiliev, 2013; Mezhdistsiplinarnyie issledovaniya…, 2018; Rudaya et al., 2017). The distribution of materials by ecological groups shows that the fauna of Chagyrskaya Cave is dominated by species typical for open spaces—steppe, forest-steppe, and meadows, while the number of forest species is insignificant (Fig. 2, 3). Occurrences of mountain-steppe species (rock-voles and pikas) indicate the formation of a specific mosaic fauna associated with latitudinal zones and vertical zonality.
The analysis of the fossils from Chagyrskaya Cave was made earlier (Ivleva, 1990). It has been shown that open spaces expanded upwards the profile, from layer 7 to layer 1 (Fig. 4). Throughout the whole sequence, a mosaic type of landscape with a decrease in the afforested area and a predominance of the forest-steppe zone is observed (Ibid.: 92). The semi-aquatic species (beaver and water vole) indicate the humidity level of climate; fluctuations of their population point to climatic changes. Layers 2 and 6 were the “driest” (Ibid.).
In the Pleistocene layers of Chagyrskaya Cave, few identifiable bone remains were found. According to stratigraphic and
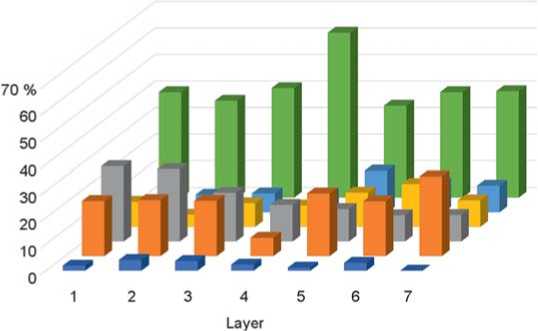
Fig. 3. The ratio of ecological groups of small mammals of Chagyrskaya Cave by layers (layers 6a–6c/2 combined into one). Legend same as on Fig. 2.
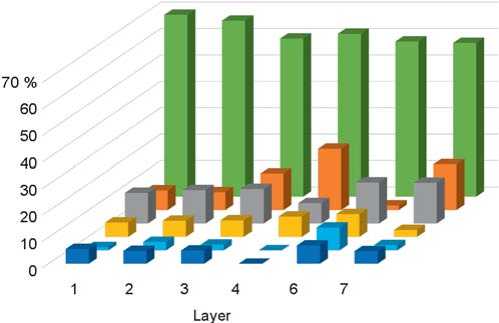
Fig. 4. Ratio of ecological groups of small mammals of Okladnikov Cave by layers. Legend same as on Fig. 2.
micromorphological analytical data, layer 6c/1 is considered undisturbed and contains faunal material; hence, its animal fossil composition was regarded as a reference for reconstructing the paleoecological conditions and the impact of post-sedimentary processes.
The small-mammal compositions of the Holocene, Late Pleistocene, and Middle Pleistocene stratigraphic units of Chagyrskaya (Table 1) and Okladnikov (Table 2) caves have been compared. First of all, the faunal composition from layers 6c/1 and 6c/2 of Chagyrskaya Cave was considered, as the least disturbed by post-sedimentation processes. In addition, the likelihood of shifting small-mammal bone remains from one layer to another as a result of colluvial and permafrost processes was assessed.
Statistical ordination methods were used to test the assumption that the composition of small mammals reflected the relevant paleoenviroments. Because the available data include a great number of variables (31) and a small number of samples (17), non-metric multidimensional scaling was conducted. The scaling results are shown on the diagram (Fig. 5, stress level 0.02).
Table 1. Bone remains of mammals of various species in the deposits of Chagyrskaya Cave, spec.
|
Biotopes |
Таксон |
Layer |
||||||||||
|
1 |
2 |
3 |
5 |
6 |
6a |
6b |
6c/1 |
6c/2 |
6d |
7 |
||
|
Semi-desert |
Alactagulus sp. |
– |
– |
– |
1 |
– |
1 |
– |
– |
– |
– |
– |
|
Allactaga sp. |
– |
1 |
2 |
1 |
– |
5 |
– |
– |
– |
– |
– |
|
|
Ellobius talpinus |
– |
– |
2 |
2 |
– |
2 |
– |
– |
– |
– |
1 |
|
|
Lagurus lagurus |
– |
51 |
83 |
184 |
1 |
112 |
3 |
– |
– |
7 |
1 |
|
|
Eolagurus luteus |
– |
7 |
8 |
19 |
– |
24 |
1 |
– |
– |
– |
– |
|
|
Steppe |
Lepus sp. |
1 |
4 |
33 |
20 |
– |
15 |
– |
– |
– |
– |
– |
|
Spermophilus sp. |
3 |
25 |
72 |
125 |
– |
190 |
6 |
– |
– |
7 |
2 |
|
|
Marmota sp. |
– |
1 |
2 |
8 |
– |
15 |
– |
– |
– |
– |
– |
|
|
Cricetulus migratorius |
– |
5 |
7 |
20 |
– |
20 |
– |
– |
– |
2 |
– |
|
|
Allocricetulus eversmanni |
1 |
6 |
5 |
4 |
1 |
1 |
– |
– |
– |
– |
– |
|
|
Forest-steppe |
Myospalax myospalax |
7 |
98 |
83 |
126 |
3 |
206 |
4 |
4 |
1 |
5 |
1 |
|
Cricetus cricetus |
3 |
51 |
48 |
3 |
– |
4 |
1 |
– |
– |
– |
– |
|
|
Lasiopodomys gregalis |
6 |
103 |
311 |
210 |
– |
242 |
11 |
8 |
– |
22 |
3 |
|
|
Microtus oeconomus |
2 |
45 |
76 |
72 |
– |
25 |
2 |
2 |
– |
7 |
1 |
|
|
Microtus arvalis |
3 |
20 |
19 |
4 |
– |
4 |
– |
– |
– |
1 |
1 |
|
|
Sicista sp. |
– |
7 |
2 |
1 |
– |
– |
– |
– |
– |
– |
– |
|
|
Forest |
Asioscalops altaica |
– |
13 |
29 |
28 |
1 |
65 |
5 |
– |
– |
10 |
1 |
|
Crocidura sp. |
1 |
7 |
1 |
– |
– |
– |
– |
– |
– |
– |
||
|
Sorex sp. |
2 |
32 |
17 |
10 |
– |
– |
1 |
– |
– |
1 |
1 |
|
|
Sciurus vulgaris |
– |
1 |
– |
4 |
– |
2 |
– |
– |
– |
– |
1 |
|
|
Eutamias sibiricus |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
|
Pteromys volans |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
– |
|
|
Apodemus sp. |
2 |
14 |
7 |
4 |
– |
– |
– |
– |
– |
– |
– |
|
|
Clethrionomys sp. |
9 |
162 |
176 |
116 |
– |
19 |
7 |
1 |
3 |
17 |
– |
|
|
Myopus / Lemmus |
– |
1 |
3 |
5 |
– |
1 |
– |
– |
– |
– |
1 |
|
|
Microtus agrestis |
– |
12 |
15 |
2 |
– |
– |
1 |
– |
– |
3 |
– |
|
|
Semi-aquatic |
Arvicola terrestris |
1 |
36 |
49 |
14 |
– |
36 |
2 |
1 |
– |
– |
1 |
|
Mountain-steppe |
Alticola sp. |
11 |
177 |
237 |
320 |
– |
267 |
15 |
4 |
– |
15 |
4 |
|
Ochotona sp. |
– |
11 |
47 |
15 |
– |
19 |
2 |
1 |
– |
3 |
– |
|
Table 2. Bone remains of mammals of various species in the deposits of Chagyrskaya Cave, spec. (Ivleva, 1990)
|
Biotopes |
Taxon |
Layer |
|||||
|
1 |
2 |
3 |
4 |
6 |
7 |
||
|
Semi-desert |
Erinacea sp. |
0 |
0 |
1 |
0 |
0 |
0 |
|
Allactaga sp. |
1 |
1 |
3 |
0 |
0 |
2 |
|
|
Ellobius sp. |
5 |
10 |
19 |
0 |
1 |
2 |
|
|
Eolagurus sp. |
3 |
4 |
10 |
0 |
4 |
9 |
|
|
Steppe |
Lepus tolai |
1 |
0 |
3 |
0 |
0 |
2 |
|
Spermophilus sp. |
16 |
16 |
43 |
0 |
3 |
9 |
|
|
Marmota sp. |
10 |
4 |
14 |
0 |
1 |
3 |
|
|
Cricetulus sp. |
14 |
9 |
37 |
1 |
1 |
3 |
|
|
Forest-steppe |
Myospalax myospalax |
197 |
44 |
136 |
4 |
12 |
31 |
|
Cricetus cricetus |
169 |
32 |
69 |
2 |
3 |
19 |
|
|
Lasiopodomys gregalis |
26 |
109 |
257 |
0 |
9 |
132 |
|
|
Microtus oeconomus |
40 |
44 |
218 |
2 |
7 |
137 |
|
|
Microtus arvalis-agrestis |
85 |
87 |
245 |
0 |
3 |
62 |
|
|
Forest |
Asioscalops altaica |
12 |
7 |
47 |
0 |
1 |
8 |
|
Sorex sp. |
15 |
10 |
15 |
0 |
3 |
9 |
|
|
Crocidura sp. |
14 |
7 |
22 |
0 |
0 |
4 |
|
|
Pteromys volans |
3 |
2 |
2 |
0 |
0 |
0 |
|
|
Clethrionomys |
24 |
20 |
90 |
1 |
4 |
52 |
|
|
Myopus sp. |
1 |
0 |
3 |
0 |
0 |
0 |
|
|
Microtus agrestis |
18 |
14 |
23 |
0 |
1 |
28 |
|
|
Inter-zonal |
Castor fiber |
2 |
3 |
1 |
0 |
0 |
1 |
|
Semi-aquatic |
Arvicola terrestris |
40 |
20 |
76 |
0 |
4 |
30 |
|
Mountain-steppe |
Alticola sp. |
42 |
25 |
169 |
2 |
0 |
99 |
|
Ochotona sp. |
14 |
7 |
45 |
1 |
1 |
14 |
|
The composition of small-mammal bones varies from layer to layer. The layers with the highest biodiversity are located in the right portion of the graph, the layers with the lowest biodiversity are in the left part. The latter cluster includes the smallest complexes from modern layer 1 and layer 4 of Chagyrskaya Cave. The composition of small mammals from layer 7 of this cave, corresponding to the Middle Pleistocene and tundra landscapes, almost completely coincides with that from overlying layers 6c/2 and 6c/1. This may be due to shifting the faunal remains through frost processes and the poor preservation of the material inside the layer. Excavations in layer 7 revealed only bones of large mammals from overlying layer 6c/2.
The composition of small mammals from layer 6d, which was formed due to freeze fracturing of the sediments in layers 6c/2 and 7, almost completely corresponds to that of layer 6c/2. This confirms the conclusions drawn from the composition of the bone material from layer 7: it was shifted from layer 6c/2 and contained poorly preserved remains. The sparse composition of small mammals from undisturbed layers 6c/2 and 6c/1 can be a result of the taphonomic processes in these stratigraphic units. Nevertheless, the similarities that have been identified between these layers suggest that the deposits were formed under similar paleoenvironmental conditions and that materials were transferred from these layers to neighboring ones.
The similarity in the composition of small mammals from layer 6b of colluvial genesis and from undisturbed layers 6c/2 and 6c/1 indicates that the remains of small mammals were transferred from these layers.
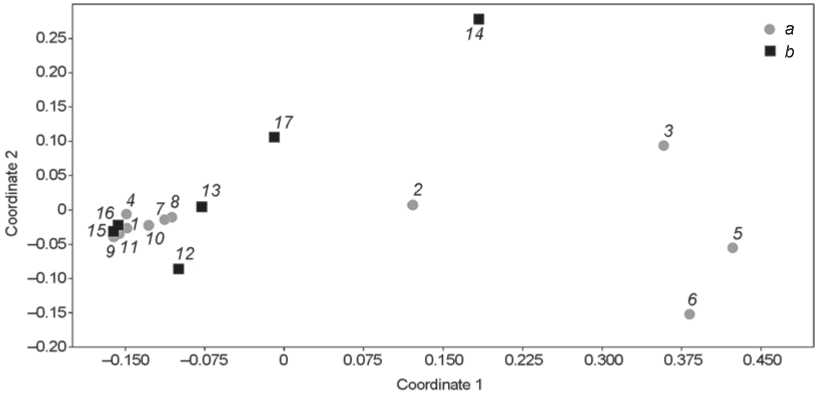
Fig. 5. Non-metric multidimensional scaling of the small fauna composition from stratigraphic sequences of Chagyrskaya ( a ) and Okladnikov ( b ) Caves.
1 – layer 1; 2 – layer 2; 3 – layer 3; 4 – layer 4; 5 – layer 5; 6 – layer 6а; 7 – layer 6b; 8 – layer 6c/1; 9 – layer 6c/2; 10 – layer 6d; 11 – layer 7; 12 – layer 1; 13 – layer 2; 14 – layer 3; 15 – layer 4; 16 – layer 6; 17 – layer 7.
The situation in Pleistocene layers 6a (colluvial genesis) and 5 (complex genesis including seismic and colluvial processes of varying degrees of intensity) is completely different. Forest-steppe environment was reconstructed for layer 5, and dry arid steppe landscapes were typical for layer 6a. The smallmammal faunal assemblages from these layers are similar in composition, but differ significantly from the bone collection of the underlying layers (6c/2, 6c/1, 6b). Layers 5 and 6a contain remains of great biodiversity, which brings them closer in composition to Holocene layer 3 containing material remains of the Bronze Age Afanasyevo culture.

The great part of small-mammal remains may have been transferred to layer 6a from overlying layers 3 and 5, rather than from underlying layers 6c/2, 6c/1, as compared to the case with layer 6b. A small proportion of small-mammal remains may nevertheless have been transferred from the underlying layers together with the remains of the artifacts of late Neanderthals. These assumptions are supported by the high degree of biodiversity of the mammal composition and the similarity of the composition of Holocene layer 3 with that of layers 5 and 6a.
Okladnikov Cave shows significant biodiversity in Pleistocene layers 2, 3, and 7. However, the composition of the small-mammal remains from layers 4 and 6 is almost identical to that from layers 6c/2 and 6c/1 of Chagyrskaya Cave.
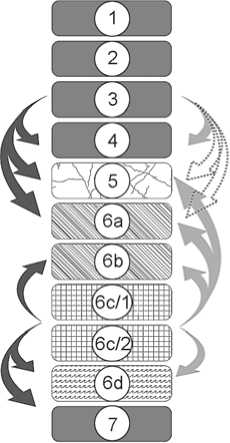
A diagram was constructed on the basis of the results of statistical analysis of the composition of small-mammal fauna from the stratigraphic units of Chagyrskaya Cave and the results of stratigraphic and micro-morphological analyses. The directions of transfer of bone remains correspond almost entirely with those of soft sediments (Fig. 6).
In order to ordinate and correlate the paleoenvironments of Altai Neanderthals, a
Fig. 6. Scheme of transfer of archaeological and osteological materials in the deposits of Chagyrskaya Cave.
1 – lithological layer; 2 – lithological layer of colluvial and seismic genesis; 3 – lithological layer of colluvial genesis; 4 – lithological layer in situ ; 5 – lithological layer of permafrost genesis; 6 – movement of small fauna; 7 – movement of Neanderthal material-culture remains;
8 – movement of loose deposits as a result of rodent activities.
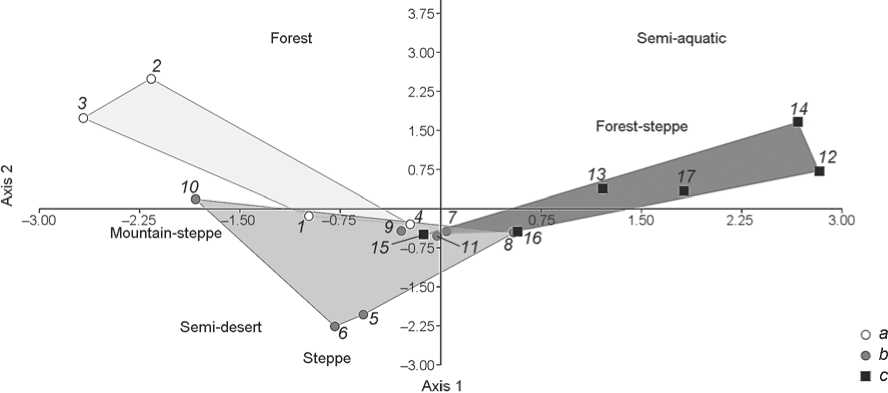
Fig. 7. Discriminant analysis of the small fauna composition in biotopes from the Holocene ( a ) and Pleistocene ( b ) deposits of Chagyrskaya Cave and Pleistocene deposits ( c ) of Okladnikov Cave.
discriminant analysis based on biotope data was carried out. To reduce variables, the remains of small mammals were grouped by biotopes (see Tables 1, 2). For the analysis, the samples were subdivided into three groups, which included the Pleistocene and Holocene layers of Chagyrskaya Cave and the lithological layers of Okladnikov Cave.
The discriminant graph, which establishes correlations of various Middle Paleolithic layers belonging to different biotopes and interrelations between layer complexes, shows that the Pleistocene deposits of Chagyrskaya Cave were accumulated under the conditions of predominance of steppeforest-steppe landscapes, and the Holocene deposits under the conditions dominated by forest-steppe-forest landscapes (Fig. 7). Soft sediments in Okladnikov Cave were formed under the conditions dominated by foreststeppe and semi-aquatic fauna.
Discussion
The Holocene layers of Chagyrskaya Cave are characterized by a high concentration of the remains of chiropterans. Six species were identified: Eptesicus nilssonii northern bat, Myotis brandtii Brandt’s bat, Myotis dasycneme pond bat, Myotis blythii lesser mouse-eared bat, Plecotus ognevi Siberian longeared bat, and Murina leucogaster greater tubenosed bat. In the Pleistocene layers, the amount of remains is significantly lower than in the Holocene layers. In layers 6b and 6c/1, only two teeth were found. This can be explained by the fact that bat bones are fragile and poorly preserved; besides, during the period of human occupation of the cave, the chiropterans may have not used the cave for denning or wintering (Agadjanian, Serdyuk, 2005). The fauna of chiropterans is not analyzed in this article, and its use for paleoreconstructions is difficult, because, first, bats represent a rather mobile group of small mammals, and, second, the number of their remains is small. Rodent remains are the most abundant in the cave deposits, with over 30 species. Among insectivores, at least five species have been identified. The remains of duplicidentates have also been identified. For each layer of Chagyrskaya Cave, the following composition of small mammals has been determined.
Layer 1. Characterized by species occurring in the present-day Altai; no exotic species were found. The tooth morphology of the background vole species is close to that of the modern one. The overall faunal composition of this layer suggests the distribution of forest-steppe stations.
Layer 2. The faunal composition is the same as in layer 1, but includes Lagurus lagurus steppe lemming, Eolagurus luteus yellow steppe lemming, and a lemming of the Lemmus genus . The first two species inhabit steppes and semi-deserts, while the third is characteristic of tundra associations. All the three species are markers of “mixed” faunas (Vangengeim, 1977; Gromov, 1948; Markov et al., 1965; Chersky, 1891). Holocene refugia of large mammals are well known (Kovacs, 2011; Vartanyan, 2004; Stuart et al., 2004; Vereshchagin, 1988); small mammals, unlike large ones, are more resistant to environmental changes
(Popova, 2014). During the accumulation of layer 2, a tundra-steppe biocoenosis may have persisted for some time in the vicinity of the cave. The pollen spectrum is dominated by synanthropic plant species.
Layer 3. The faunal composition is the same as in layer 2. The forest-steppe formations dominate, but there are representatives of “mixed” periglacial faunas and wood-shrub biotopes ( Sicista birch mouse, and forest mice of Apodemus genus), and Allactaga jerboa, which prefers semi-desert habitats. According to our statistical calculations, layer 3 is close to layers 5 and 6a in terms of biodiversity. Partial transfer of materials between layers is possible. Large mammals are represented by bison, a typical inhabitant of vast open landscapes from forest-steppe to tundra steppe (Mezhdistsiplinarnyie issledovaniya…, 2018). The occurrence of remains of this artiodactyl in the area of Chagyrskaya Cave suggests that the area was a refugium for mammoth fauna in the Holocene. The pollen data for this layer indicate forest-steppe with arboreal components, which is consistent with the composition of small mammals.
Layer 4. The amount of bones is very low for the Holocene layer, the animal community is dominated by voles (narrow-skulled, common, and tundra one), while other species are rare. Judging by the composition of the small-mammal bone collection, the layer may be a buried rodent hole containing pellets of a predator (Shalabaev, 2011).
Layer 5. The small-mammal fauna is the most abundant for the Pleistocene deposits of Chagyrskaya Cave. It indicates the steppe and semi-desert biotopes during the formation of the layer. There are remains of Allactaga marmot and Alactagulus tarbagan inhabiting takyrs with dense clayey soil. Steppe lemming and Eolagurus luteus yellow steppe lemming are also common in such semi-deserts. Layer 5 was apparently formed under periglacial cold steppe conditions; this is evidenced by the remains of Lemmus sibiricus lemming. Pollen data indicate a taiga biome for this layer. Typical taiga small-mammal species are represented by few teeth and bones of squirrels; no bones of other typical forest-dwelling species were found. Squirrel remains may be associated with small forests along riverbanks, which did not play a significant role (Mezhdistsiplinarnyie issledovaniya…, 2018). The remains of forest mice attest to arboreal and shrub vegetation. Large mammals were dominated by steppe and forest-steppe species. As shown earlier, cave hyenas were involved in the formation of the large-mammal taphocoenosis (Kolobova et al., 2019b); these animals are rightfully considered unique collectors, they pick up the remains of almost the full range of medium and large-mammal fauna in their hunting territory. It is likely that humans visited the cave infrequently at this time; hence, it was also attractive to predators gathering small fauna, such as birds of prey or small polecats. In the context of our study, it is important that large predators probably dug the cave soil while making dens and thus caused some movement of the archaeological material, which may partially penetrated into the overlying or underlying layers.
Layer 6a. Like the previous layer, it reveals a large amount of bone remains. The layer is characterized by a greater number of desert and semi-desert smallmammal species than layer 5. The Siberian lemming was noted. Layers 5 and 6a were formed under different conditions. In the fauna of large mammals, the number of forest-steppe species in layer 6a is lower than in layer 5. The pollen data for this layer suggest a transition from “taiga” (layer 5) to “steppe” (layer 6b). The small-mammal fauna does not show a clear “transition”. As no obvious taiga fauna was recorded in layer 5, layer 6a shows neither a reduction in the number of forest species, nor an increase in the number of steppe species. In general, the period of formation of layers 6 coincided with the period of human habitation in the cave. This explains the small number of small mammals remains: wild animals and birds avoided close proximity to humans.
Layer 6b. The amount of small-mammal remains is sparse. Neither jerboa nor wood-shrub dweller remains occurred in the layer. The fauna includes representatives of meadow and meadow-steppe biotopes (gopher, mole rat, common vole, and root vole), while the number of steppe species is insignificant. The large-mammal fauna is dominated by steppe species. The pollen data indicate an increase in cereal pollen, which is inconsistent with the data on small mammals. This can be explained by the small amount of material or other taphonomic features.
Layers 6c/1 and 6c/2. The amount of small-mammal remains is also low. The species composition is almost identical to that of layer 6b. According to statistical and micro-morphological analyses, layer 6c is the source of the formation of the taphocoenosis of layer 6b. A partial transfer of archaeological materials from the layer is possible owing to human economic activity in the cave at that time. The fauna is dominated by the remains of meadow-steppe stations; few osteological remains have been associated with the rocky habitats. Large-mammal community is dominated by steppe species. The pollen data show the reduction in the number of cereals.
Layer 6d. Small-mammal community of this layer is dominated by grey voles. The composition of the microfauna, almost entirely corresponding to that of layer 6c/2, supports geological definition that the layer was formed due to permafrost processes.
Layer 7. The amount of materials for biological analysis from this layer is negligible. Among small mammals, representatives of “mixed” periglacial fauna are noted; remains of mole vole avoiding feathergrass and sagebrush steppe were recorded. There are also species of mountain-steppe and grassland habitats. According to the mentioned data, the climate was arid. Large-mammal community is dominated by steppe species. Pollen samples attest to the taiga, steppe, and tundra biomes; these data do not correlate with the paleotheriological facts. This discrepancy is explainable by ingression of the overlying sediments, which does not contradict the results of the statistical analysis.
Data of the microtheriological analysis of finds from Okladnikov Cave indicate a predominance of forest-steppe biotopes in the cave area and warming of the climate closely before the Holocene. Remains of inhabitants of the “mixed” periglacial landscapes were also found in the cave. Four teeth of the forest lemming of Myopus genus were discovered (Ivleva, 1990). These lemmings inhabited various forests— spruce, fir, cedar, and mixed forests, i.e. their biotopic allocation was not the same as that of the tundra lemming. According to recent data, forest lemmings differ from the tundra ones in the third upper molar and the morphometric characteristics of other teeth (Ponomarev et al., 2011). Probably, a revision of the lemming material from Okladnikov Cave is needed to confirm or refute the occurrences of tundra components in the Okladnikov Cave area in the Pleistocene.
Discriminant analysis confirmed the results of both microtheriological analysis and multidimensional scaling, and revealed statistically significant similarities in the composition of the small-mammal fauna (Chagyrskaya Cave – layers 6c/1, 6c/2, 6b, Okladnikov Cave – layer 6). According to the results of discriminant analysis, the late Altai Neanderthals, whose traces of habitation were found in both caves, lived predominantly in forest-steppe landscapes. The significant presence of semi-aquatic species in the sediments of Okladnikov Cave may be due to the shorter distance to the river as compared to the distance from Chagyrskaya Cave to the river.
In general, the data on small mammals from the two caves indicate similar landscapes and conditions in the Karga period. According to the microtherological records, forest-steppe ecozone dominated in the low Altai Mountains for almost the entire period of sedimentation.
Conclusions
This study has shown that in the Altai in the final MIS 4–early MIS 3, late Neanderthals lived in similar paleoenvironments dominated by steppe-forest-steppe landscapes. The high proportion of semi-aquatic species in the faunal complexes of Okladnikov Cave probably attests to different hydrological regimes of the Sibiryachikha and the Charysh Rivers in the Pleistocene. Perhaps the Charysh was not as full then as it is now.
Despite the small sample-size of the two sites, it can be assumed that the choice of the late Altai Neanderthals (inhabitants of two caves in similar paleoecological conditions) could have hardly been accidental. The Neanderthals from the Micoquian populations of Central and Eastern Europe hunted a wide range of large mammals, which included almost all representatives of the mammoth fauna (Richter, 2006). In the complexes of the geographically closest Caucasus and Crimea, Neanderthals hunted predominantly large herbivores (bison, Pleistocene donkey, etc.) (Uthmeier, Chabai, 2010; Golovanova et al., 2018; Ramírez-Pedraza et al., 2020). The Sibiryachikha Neanderthals, who hunted bison and horse, likely occupied the most suitable ecological niches for this activity—caves in river valleys, which served as transit corridors for seasonal migrations of large herbivores. A similar dependence of ancient hominin adaptation strategies on habitats and migrations of Pleistocene animals has already been documented in several areas of Central and North Asia (Agadjanian, Shunkov, 2018; Khatsenovich et al., 2021; Zavala et al., 2021).
Another reason for settling in these karst cavities was probably the possibility of extracting jasperoids of the Zasurye formation and chalcedony—high-quality lithic raw materials used for making tools typical of the Sibiryachikha industries (plano-convex bifaces and convergent scrapers). The Chagyrskaya and Okladnikov Caves are the only Middle Paleolithic sites in the region that are located on outcrops of these rocks (Derevianko et al., 2015; Kolobova et al., 2019b). The dependence of choice of human settlement on the availability of high-quality lithic materials been noted exclusively for the Upper Paleolithic sites in North and Central Asia (Rybin et al., 2020).
The statistical study of the composition of the fauna of small vertebrates proved useful for comparing complexes containing dozens of species; statistical data make it possible to draw the conclusions about post-sedimentary processes, which do not contradict the stratigraphic and micromorphological data, but complement them. It has been determined that the faunal composition of the individual layers of Chagyrskaya Cave is influenced by the state preservation of organic materials within the layer. For example, in layer 7, to which pollen data suggest taiga/ tundra conditions during the sedimentation period, small mammals from the overlying stratigraphic units (layer 6c/2) were recorded, with compositions corresponding to steppe-forest-steppe landscapes. The fauna of small mammals from the colluvial layers (6b) largely coincides with that of the source layer of redeposition, confirming the assumption about transportation of not only Neanderthal cultural remains, but also fauna. The faunal composition also depends on various disturbances of the layers, especially rodent burrows from the overlying layers, even though the excavations were carried out in the up-to-day way. Layer 6a, for example, showed greater impact from the overlying layers (3 and 5) than from the undisturbed stratigraphic unit containing numerous archaeological materials and large animal bones.
Small-mammal fauna data combined with the results of stratigraphic, micromorphological, and pollen studies can be used in the reconstruction of postsedimentation processes in karst sites. In Chagyrskaya Cave, the recorded transfer of small-mammal remains between layers is almost entirely consistent with post-sedimentary processes.
Paleoreconstructions based on the small-mammal fauna and pollen are usually carried out apart from detailed stratigraphic reconstructions that take into account episodes of displacement or redeposition of stratigraphic units. Assemblages of each layer are considered as discrete units reflecting the sequential development of paleolandscapes from the bottom to the top of the sections, which may not completely correspond to the sedimentation processes. The study of Chagyrskaya Cave complexes revealed the complexity of stratigraphic processes in karst cavities, their impact on the composition of the small-mammal fauna and, ultimately, on paleoreconstructions.
Acknowledgements
The analysis of the small-mammal fauna was supported by the Russian Foundation for Basic Research, Project No. 1809-40070; the statistical study was supported by the Russian Science Foundation, Project No. 21-18-00376.
References Paleoenvironmental Conditions of Neanderthal Habitation in the Altai: Chagyrskaya and Okladnikov Caves
- Agadjanian A.K., Serdyuk N.V. 2005 History of mammalian communities and paleogeography of Altai Mountains in the Paleolithic. Paleontological Journal, vol. 39 (6): 645-820.
- Agadjanian A.K., Shunkov M.V. 2018 Late Pleistocene mammals of the Northwestern Altai : Report 1. Anui Basin. Paleontological Journal, vol. 52: 1450-1460.
- Bassinot F.C., Labeyrie L.D., Vincent E., Quidelleur X., Shackleton N.J., Lancelot Y. 1994 The astronomical theory of climate and the age of the Brunhes-Matuyama magnetic reversal. Earth and Planetary Science Letters, vol. 126: 91-108.
- Baumann M., Plisson H., Rendu W., Maury S., Kolobova K., Krivoshapkin A. 2020 Neandertalian bone industry at Chagyrskaya cave (Altai, Russia). Quaternary International, vol. 559: 68-88. Chersky I.D. 1891
- Opisaniye kollektsii posletretichnykh mlekopitayushchikh zhivotnykh, sobrannykh Novo-Sibirskoy ekspeditsiyey 1885-1886 gg. St. Petersburg: Imp. akad. nauk.
- Derevianko A.P., Markin S.V. 1992 Mustye Gornogo Altaya (po materialam peshchery im. Okladnikova). Novosibirsk: Nauka.
- Derevianko A.P., Markin S.V., Kulik N.A., Kolobova K.A. 2015 Lithic raw material exploitation in the Sibiryachikha facies, the Middle Paleolithic of Altai. Archaeology, Ethnology and Anthropology of Eurasia, vol. 43 (3): 3-16.
- Derevianko A.P., Markin S.V., Zykin V.S., Zykina V.S., Zazhigin V.S., Sizikova A.O., Solotchina E.P., Smolyaninova L.G., Antipov A.S. 2013 Chagyrskaya cave: A Middle Paleolithic site in the Altai. Archaeology, Ethnology and Anthropology of Eurasia, vol. 41 (1): 2-27.
- Golovanova L.V., Doronichev V.B., Aleksandrovsky A.A. 2018 Peshchery Lagonakskogo nagorya: Stratigrafiya i paleoekologiya arkheologicheskikh pamyatnikov kamennogo veka. Vopory stratigrafi i. Iss. 147: Speleologiya i karstovedeniye: 268-298.
- Gromov V.I. 1948 Paleontologicheskoye i arkheologicheskoye obosnovaniye stratigrafi i kontinentalnykh otlozheniy chetvertichnogo perioda na territorii SSSR. Moscow: Izd. AN SSSR.
- Hammer Ø., Harper D.A.T., Ryan P.D. 2001 PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, vol. 4 (1): 4.
- Imbrie J., Hays J.D., Martinson D.G., McIntyre A., Mix A.C., Morley J.J., Pisias N.G., Prell W.L., Shackleton N.J. 1984 The orbital theory of Pleistocene climate: Support from a revised chronology of the marine δ18O record. In Milankovitch and Climate, pt. 1. New York: Springer, pp. 269-305.
- Ivleva N.G. 1990 Mikroteriologicheskiye materialy iz peshcher im. Okladnikova i Denisova na Altaye. In Kompleksniye issledovaniya paleoliticheskikh obyektov basseina r. Anuy. Novosibirsk: IIFF SO RAN, pp. 82-104.
- Khatsenovich A.M., Vishnevskaya I.A., Bazargur D., Volvakh A.O., Klementiev A.M., Ge J., Zhilich S.V., Marchenko D.V., Okuneva T.G., Kogai S.A., Gunchinsuren B., Rybin E.P., Olsen J.W., Derevianko A.P. 2021 Late Pleistocene paleoenvironments and episodic human occupations in the Orkhon valley of Central Mongolia. Archaeology, Ethnology and Anthropology of Eurasia, vol. 49 (2): 3-22.
- Kolobova K.A., Chabai V.P., Shalagina A.V., Krajcarz M.T., Krajcarz M., Rendu W., Vasiliev S.K., Markin S.V., Krivoshapkin A.I. 2019a Exploitation of the natural environment by Neanderthals from Chagyrskaya Cave (Altai). Quartär, vol. 66: 7-31.
- Kolobova K.A., Shalagina A.V., Chabai V.P., Markin S.V., Krivoshapkin A.I. 2019b Signifi cation des technologies bifaciales au Paléolithique moyen des montagnes de l’Altaï. L’Anthropologie, vol. 123 (2):276-288.
- Kolobova K., Rendu W., Shalagina A., Chistyakov P., Kovalev V., Baumann M., Koliasnikova A., Krivoshapkin A. 2020a The application of geometric-morphometric shape analysis to Middle Paleolithic bone retouchers from the Altai Mountains, Russia. Quaternary International, vol. 559: 89-96.
- Kolobova K., Roberts R., Chabai V., Jacobs Z., Krajcarz M., Shalagina A., Krivoshapkin A., Li B., Uthmeier T., Markin S., Morley M., O’Gorman K., Rudaya N., Talamo S., Viola B., Derevianko A. 2020b Archaeological evidence for two separate dispersals of Neanderthals into Southern Siberia. Proceedings of the National Academy of Sciences, vol. 117 (6): 2879-2885
- Kovacs J. 2011 Radiocarbon chronology of Late Pleistocene large mammal faunas from the Pannonian basin (Hungary). Bulletin of Geosciences, vol. 87: 13-19.
- Markov K.K., Lazukov G.I., Nikolaev V.A., Velichko A.A. 1965 Chetvertichniy period: (Lednikoviy period - antropogenoviy period), vol. I. Moscow: Izd. Mosk. Gos. Univ. Mezhdistsiplinarnyie issledovaniya Chagyrskoy peshchery - stoyanki srednego paleolita Altaya. 2018
- Derevianko A.P., Markin S.V., Kolobova K.A., Chabay V.O., Rudaya N.A., Viola B., Buzhilova A.P., Mednikova M.B., Vasiliev S.K., Zykin V.S., Zykina V.S., Zazhigin V.S., Volvakh A.O., Roberts R.G., Jacobs Z., Bo Li. Novosibirsk: Izd. IAET SO RAN.
- Ponomarev D.V., Isaiychev K.I., Bikbaeva G.V., Kryazheva I.V. 2011 Materialy k rodovoy diagnostike lemmingov Lemmus i Myopus. Vestnik Instituta geologii Komi Nauchnogo tsentra UrO, No. 11: 10-11.
- Popova L.V. 2014 Sudby mamontovoy fauny i fenomen Chernoy korolevy. Visnik Natsionalnogo naukovo-prirodnichogo muzeyu, No. 12: 20-32.
- Ramírez-Pedraza I., Rivals F., Uthmeier T., Chabai V.P. 2020 Palaeoenvironmental and seasonal context of the Late Middle and Early Upper Palaeolithic occupations in Crimea: An approach using dental wear patterns in ungulates. Archaeological and Anthropological Sciences, iss. 268.
- Richter J. 2006 Neanderthals in their landscape. In Neanderthals in Europe. Liège, Tongeren: ERAUL 117 & ATVAT VCA, pp. 17-32.
- Rudaya N., Vasiliev S., Viola B., Talamo S., Markin S. 2017 Palaeoenvironments during the period of the Neanderthals settlement in Chagyrskaya cave (Altai Mountains, Russia). Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 467: 265-276.
- Rybin E.P., Paine C.N., Khatsenovich A.M., Bolorbat T., Talamo S., Marchenko D.V., Rendu W., Klementiev A.M., Odsuren D., Gunchinsuren B., Zwyns N. 2020 A new Upper Paleolithic occupation at the site of Tolbor-21 (Mongolia): Site formation, human behavior and implications for the regional sequence. Quaternary International, vol. 559:133-149.
- Salazar-García D.C., Power R.C., Rudaya N., Kolobova K., Markin S., Krivoshapkin A., Henry A.G., Viola, B. 2021 Dietary evidence from Central Asian Neanderthals: A combined isotope and plant microremains approach at Chagyrskaya Cave (Altai, Russia). Journal of Human Evolution, vol. 156.
- Shalabaev R.N. 2011 Izbiratelnost khishchnykh ptits v pitanii Micromammalia na ostepenennoy territorii. Sovremenniye problemy nauki i obrazovaniya, No. 4: 51-56.
- Shalagina A.V., Kharevich V.M., Mori S., Bomann M., Krivoshapkin A.I., Kolobova K.A. 2020 Rekonstruktsiya tekhnologicheskikh tsepochek proizvodstva bifasialnykh orudiy v industrii Chagyrskoy peshchery. Sibirskiye istoricheskiye issledovaniya, No. 3: 130-151.
- Shalagina A.V., Krivoshapkin A.I., Kolobova K.A. 2015 Truncated-faceted pieces in the Paleolithic of Northern Asia. Archaeology, Ethnology and Anthropology of Eurasia, vol. 43 (4): 33-45.
- Slon V., Mafessoni F., Vernot B., Filippo C. de, Grote S., Viola B., Hajdinjak M., Peyrégne S., Nagel S., Brown S., Douka K., Higham T., Kozlikin M.B., Shunkov M.V., Derevianko A.P., Kelso J., Meyer M., Prüfer K., Pääbo S. 2018 The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature, iss. 561: 113-116.
- Stuart A.J., Kosintsev P.A., Higham T.F. G., Lister A.M. 2004 Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth. Nature, iss. 431: 684-689.
- Uthmeier T., Chabai V.P. 2010 Neanderthal subsistence tactics in the Crimean Micoquian. In Settlement dynamics of the Middle Paleolithic and Middle Stone Age, vol. III. Tübingen: Kerns Verlag, pp. 195-234.
- Vangengeim E.A. 1977 Paleontologicheskoye obosnovaniye stratigrafi i antropogena Severnoy Azii. Moscow: Nauka.
- Vartanyan S.L. 2004 Paleogeografiya pozdnego neoplestotsena i golotsena territorii ostrova Vrangelya. Cand. Sc. (Geography) Dissertation. St. Petersburg.
- Vasiliev S.K. 2013 Large mammal fauna from the Pleistocene deposits of Chagyrskaya Cave, Northwestern Altai (based on 2007-2011 excavations). Archaeology, Ethnology and Anthropology of Eurasia, vol. 41 (1): 28-44.
- Vereshchagin N.K. 1988 Paleogeografiya i paleoekologiya zverey mamontovoy fauny. In Obshchaya i regionalnaya teriogeografi ya. Moscow: Nauka, pp. 19-32.
- Vernot B., Zavala E.I., Gómez-Olivencia A., Jacobs Z., Slon V., Mafessoni F., Romagné F., Pearson A., Petr M., Sala N., Pablos A., Aranburu A., Castro J.M.B., Carbonell E., de, Li B., Krajcarz M.T., Krivoshapkin A.I., Kolobova K.A., Kozlikin M.B., Shunkov M.V, Derevianko A.P., Viola B., Grote S., Essel E., Herráez D.L., Nagel S., Nickel B., Richter J., Schmidt A., Peter B., Kelso J., Roberts R.G., Arsuaga J.-L., Meyer M. 2021 Unearthing Neanderthal population history using nuclear and mitochondrial DNA from cave sediments. Science, vol. 372 (6542).
