Палеофенетический анализ современного и древнего населения Чукотки

Автор: Мовсесян А.А.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология
Статья в выпуске: 3 (51), 2012 года.
Бесплатный доступ
По данным о 30 дискретно-варьирующих признаках изучено 58 черепов из Эквенского и 107 из Уэленского могильников, относящихся к древнеберингоморской культуре Чукотки. Обнаружено сходство древнеберингоморских популяций не только с современными арктическими, но и с байкальскими группами. Это может объясняться как контактами континентальных и арктических монголоидов Сибири в древности, так и общностью их генетических корней. Результаты сравнения древних популяций Чукотки с неолитическим населением Прибайкалья и южными монголоидами позволяют предположить, что образование протоэскимосоалеутской этнокультурной общности могло происходить на территории Чукотки в процессе культурной и биологической адаптации к экстремальным условиям Берингии переместившихся сюда групп как внутриконтинентального населения Сибири, так и мигрантов из Южной Азии и Тихоокеанского региона.
Антропология, дискретно-варьирующие признаки, древнее население чукотки, чукчи, эскимосы
Короткий адрес: https://sciup.org/14522932
IDR: 14522932 | УДК: 572.9
Текст научной статьи Палеофенетический анализ современного и древнего населения Чукотки
При сравнении современных и древних популяций по краниометрическим данным мы зачастую сталкиваемся с явлением эпохальной изменчивости признаков, искажающим реальную картину генетических взаимоотношений между этими популяциями. Более четкую информацию о генетической преемственности можно получить при использовании признаков, отражающих генетическую структуру популяций и не подверженных изменчивости во времени. Данная статья посвящена сравнительному анализу древнеберингоморских и современных североазиатских популяций с помощью дискретно-варьирующих признаков на черепе. Их использование в качестве «генетических маркеров» вполне оправданно, поскольку дискретные вариации в строении черепа относятся к особому классу морфологических признаков – фенов. Фенетика популяций предполагает, что фены отражают генетическую конституцию особи, а частота их встречаемости – генетические особенности группы особей. В силу этого появляется возможность распространения генетических подходов и принципов на популяции, генетическое изучение которых затруднено или невозможно [Тимофеев-Ресовский, Яблоков, 1973; Тимофеев-Ресовский, Яблоков, Глотов, 1973].
Многие косвенные данные говорят о том, что вариации в строении черепа появляются в процессе нормального развития и в определенной степени детерминированы генетически [Cheverud, Buikstra, 1981; Sjøvold, 1984; Lane, 1977]. Кроме того, многочисленными исследованиями показано, что генетически отдаленные друг от друга группы отчетливо различаются по частотам этих признаков, а генетически близкие обнаруживают значительное сходство. Таким образом, привлечение популяционно-генетических методов и подходов к исследованию фенофондов популяций, охарактеризованных по частотам дискретных признаков, весьма актуально для изучения ископаемых популяций человека. Подробные программа и методика определения дискретно-варьирующих признаков на черепе были даны нами ранее [Мовсесян, Мамонова, Рычков, 1975; Мовсесян, 2005].
Изучено 58 черепов из Эквенского и 107 из Уэлен-ского могильников, относящихся к древнеберинго-морской культуре Чукотки, культуре пережиточного неолита. Для сравнительного анализа были использованы опубликованные ранее данные о современных и древних сибирских народностях [Мамонова, Мовсесян, 1998; Мовсесян, 2005]. Важно отметить, что час-
Археология, этнография и антропология Евразии 3 (51) 2012
тота каждого признака в популяции вычислялась как отношение числа его встречаемости к количеству черепов, на которых было возможно определение признака. Статистическая обработка данных производилась с помощью следующих методов: 1) вычисления обобщенных генетических расстояний ( d ) по методу М. Нея [Nei, 1972]; 3) классификационного анализа в рамках пакета PHYLIP (Phylogeny Inference Package) [Felsenstein, 1989]; 2) канонического анализа (программа «Каноклас», разработанная В.Е. Дерябиным).
Впервые черепа из Уэленского и Эквенского могильников были изучены М.Г. Левиным [1962], который определил их как типично эскимосские, сделав на этом основании вывод о глубокой древности эскимосского типа. К такому же заключению пришел и
Г.Ф. Дебец, изучавший вопрос о формировании антропологического типа эскимосов [1975]. Одонтологический анализ древнеберингоморских серий показал их сходство друг с другом и с группами арктической расы, а также сохранение в обеих сериях архаических признаков строения зубной системы, характерных для эпох мезолита – бронзы [Зубов, 1969].
Сходство между эквенской и уэленской популяциями выявляется и по частотам дискретно-варьирую-щих признаков. Обобщенное генетическое расстояние между ними достаточно мало ( d = 0,030). По-видимо-му, Уэленский и Эквенский могильники принадлежали локальным популяциям одной и той же этнической группы. При сравнении с современными арктическими популяциями (табл. 1) можно заметить, что древние бе-
Таблица 1. Распределение частот дискретно-варьирующих признаков в древних популяциях Чукотки и современных арктических группах
В то же время нельзя не отметить сходства древнего населения Чукотки с представителями байкальской расы – ульчами и негидальцами. Известно, что у современных оленных и береговых чукчей также выявляются черты, свидетельствующие, по мнению М.Г. Левина [1958], о примеси байкальского антропологического типа. Сравнивая современные арктические популяции с континентальными североазиатскими монголоидами по фенетическим данным, мы получа- ем аналогичные результаты (табл. 3). Обобщенные генетические расстояния показывают, что эскимосы в наибольшей степени приближаются к береговым чукчам. Чукчи береговые и оленные довольно схожи друг с другом. Алеуты занимают особое положение, о чем будет сказано ниже. Таким образом, чукчи и эскимосы образуют довольно изолированную общность, к которой относительно близки, кроме алеутов, негидальцы и ульчи. Еще более отчетливо эта картина проявляется на диаграмме, показывающей взаимное расположение современных сибирских популяций в пространстве канонических переменных (рис. 1). Представители байкальской расы – ульчи, негидальцы, орочи и эвенки, – образующие компактную группу, оказались вместе с эскимосами, чукчами и алеутами в области положительных значений первой переменной. Примечательно, что два независимых метода статистического анализа дали сходные результаты.
Следовательно, мы можем высказать предположение о генетических контактах арктических групп с популяциями байкальского расового типа, относящихся, по крайней мере, к эпохе древнеберингоморской культуры, а может быть, и к более раннему времени, тем более что наблюдается некоторый параллелизм в генетических и культурных связях этого региона на протяжении тысячелетий. Так, С.И. Руденко, исследовавший проблему происхождения древнеберингомор-
Таблица 2. Генетические расстояния между древним населением Чукотки и современными сибирскими этносами
|
Группа |
Уэлен |
Эквен |
Древняя Чукотка (средние) |
|
Эскимосы |
0,051 |
0,061 |
0,050 |
|
Чукчи береговые |
0,050 |
0,068 |
0,052 |
|
Чукчи оленные |
0,041 |
0,071 |
0,047 |
|
Алеуты |
0,044 |
0,059 |
0,046 |
|
Негидальцы |
0,044 |
0,051 |
0,041 |
|
Ульчи |
0,048 |
0,055 |
0,043 |
|
Эвенки |
0,056 |
0,075 |
0,060 |
|
Орочи |
0,071 |
0,074 |
0,069 |
|
Селькупы |
0,075 |
0,071 |
0,066 |
|
Бельтыры |
0,143 |
0,127 |
0,130 |
|
Сагайцы |
0,111 |
0,090 |
0,096 |
|
Шорцы |
0,098 |
0,077 |
0,079 |
|
Койбалы |
0,109 |
0,081 |
0,090 |
|
Качинцы |
0,076 |
0,085 |
0,073 |
|
Теленгеты |
0,101 |
0,106 |
0,095 |
|
Буряты |
0,099 |
0,115 |
0,100 |
|
Тувинцы |
0,068 |
0,093 |
0,075 |
Таблица 3. Генетические расстояния между арктическими и континентальными популяциями Сибири
Следует пояснить, что методология и принципы организации исследования, используемые нами в этногенетическом анализе, базируются на подходе к этническим группам как популяционным системам, обладающим «генетической памятью», т.е. сохраняющим исходные генетические характеристики. Это важное свойство популяционных систем было впервые обнаружено А.С. Серебровским [1935] и позднее аргументировано и экспериментально доказано Ю.Г. Рычковым и Ю.П. Алтуховым [Рычков, 1969, 1973; Алтухов, Рычков, 1970; Рычков, Мовсесян, 1972; Алтухов, Победоносцева, 1978]. С позиций такого подхода популяционная система представляется как подразделенная популяция, имеющая собственную микроэволюцион-ную историю и состоящая из субпопуляций, связанных единством происхождения. В вышеперечисленных работах убедительно показано, что она отличается генетической стабильностью во времени и пространстве. Благодаря исторической обусловленности этногенетических процессов возможно длительное существование этнических общностей и сохранение в генофонде современного человечества «памяти» о предшествовавших этапах развития на протяжении десятков, сотен и тысяч поколений. Следовательно, путем усреднения частот генов по популяциям, являющимся частью популяционной системы, можно реконструировать генофонд предковой популяции, что позволяет проводить сопоставления современных и древних групп не на диахронном, а на условно синхронном уровне.
Таким образом, средние характеристики сибирских расовых групп, равно как и древнеберингоморских по-
1,0
0,8 "
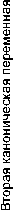
0,6 -
0,4 -
0,2 "
-1,0
Бельтыры
Сагайцы
Шорцы
Теленгеты
Чукчи береговые
БУРЯТЫ Тувинцы
Койбалы А
Алеуты
Эскимосы
Качинцы
Монголы Эвенки
Ханты
Ульчи
Чукчи оленные Орочи Негидальцы
Манси
’-1,0 -0,8 -0,6 -0,4 -0,2
О 0,2 0,4 0,6 0,8 1,0 1,2
Первая каноническая переменная
Рис. 1. Результаты канонического анализа современных сибирских популяций.
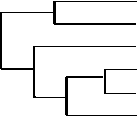
пуляций, должны в какой-то степени отражать, согласно изложенному выше подходу, генетическую структуру гипотетических предковых общностей. В этом случае сравнение средних характеристик равнозначно сопоставлению самих предковых общностей для выявления древних генетических связей между ними. Такой сравнительный анализ показал практически одинаковую степень близости древнеберингоморских популяций к арктической и байкальской группам (табл. 4). Это свидетельствует, по-видимому, в пользу предположения о генетических связях древних берингоморцев с более южными регионами. В то же время континентальные группы, включая байкальскую, значительно ближе к неолитическому населению Прибайкалья. При построении обобщенного классификационного древа древние берингоморцы объединяются с арктическими популяциями (с позиций предложенного выше подхода – с их предковой общностью), континентальные же группы тяготеют к прибайкальским неолитическим (рис. 2). Особенно наглядно сходство современного и древнего арктического населения проявляется на диаграммах, представляющих распределение частот 15 наиболее полиморфных признаков в отдельных группах и реконструированных предковых общностях (рис. 3).
Итак, судя по полученным результатам, палеофе-нетические данные свидетельствуют об уходящем корнями в далекое прошлое генетическом единстве популяций арктической группы и о сохранении в их фенофонде «памяти» о носителях древнеберингомор-ской культуры. Последняя, в свою очередь, являлась одним из этапов древнеэскимосской культуры морских зверобоев, возникшей во II тыс. до н.э. на берингомор-ском побережье Чукотки в процессе адаптации к специфическим природным условиям. Предполагается, что создатели этой культуры были потомками позднепалеолитических протоэскимосоалеутов, в конце
Арктическая раса
Древняя Чукотка Байкальская раса Уральская раса Центрально-азиатская раса Неолит Прибайкалья
Таблица 4. Генетические расстояния между древним населением Чукотки и современными сибирскими расовыми группами
|
Группа |
Арктическая |
Байкальская |
Уральская |
Центрально-азиатская |
|
Уэлен |
0,035 |
0,037 |
0,065 |
0,061 |
|
Эквен |
0,048 |
0,046 |
0,053 |
0,074 |
|
Древняя Чукотка (средние) |
0,034 |
0,036 |
0,052 |
0,062 |
|
Неолит Прибайкалья |
0,038 |
0,028 |
0,024 |
0,025 |
Рис. 2. Обобщенное классификационное древо современных и древних сибирских монголоидов.
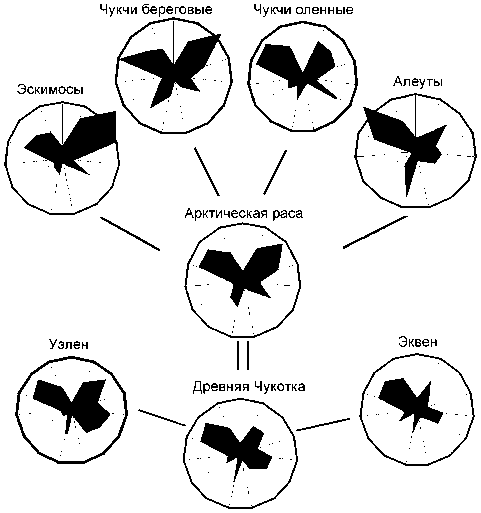
Рис. 3. Сравнение фенетической структуры современных и древних арктических популяций.
Частота 0 – в центре круга, 0,25 – по окружности.
плейстоцена и начале голоцена распространившихся по берингийской суше из Азии в Северо-Западную Америку [Диков, 1974].
Однако остается открытым вопрос о генетических истоках древнего населения Берингии. Попробуем вна- чале вслед за А.М. Золотаревым [1937] найти генетические корни протоэскимосоалеутов во внутриконти-нентальных районах Северной Азии. К сожалению, мы не располагаем достаточными палеоантропологическими материалами с близких к Чукотке территорий Сибири. Однако представляется оправданным сравнение древнеберингоморской серии с черепами из неолитических погребений Прибайкалья, датируемых VII–II тыс. до н.э.
Сравнительный анализ показал, что фенетически «палеоэскимосы» отличаются от прибайкальского неолитического населения не более, чем от современных арктических групп, обнаруживая наибольшую близость к носителям китойской культуры (VII–VI тыс. до н.э.) (табл. 5). На диаграммах, представляющих распределение частот дискретных признаков в древнеберингомор-ских и прибайкальских популяциях, можно заметить сходство «фенетических узоров» серовской и глазков-ской групп и отличие от них хронологически более ранней китойской, которая в определенной степени близка к реконструированной древнечукотской предковой общности (рис. 4).
Результаты канонического анализа современных и древних популяций Сибири полностью согласуются с предыдущими и дополняют их (рис. 5). В области положительных значений второй переменной расположились популяции арктической и байкальской рас, а также уэленская, эквенская и китойская группы. Представители центрально-азиатской и уральской рас оказались вместе с серовцами и глазковцами в области отрицательных значений.
Довольно примечательно отделение алеутов от остальных арктических групп в пространстве канонических переменных и вхождение их в североазиатскую общность. Как известно, алеуты занимают особое место в системе арктических популяций. Лингвистически и этнографически они сближаются с эскимосами, но по морфологическим признакам довольно сильно от них отличаются. Морфологическое своеобразие алеутов было отмечено во многих работах [Токарева, 1937; Hrdlicka, 1944; Дебец, 1951; Рычков, Шереметьева, 1972; Шереметьева, Рычков, 1978; Алексеев, 1981]. Если Г.Ф. Дебец рассматривал алеутов как локальную форму арктической расы, то в классификации Н.Н. Че-боксарова [1947] они входят в одну группу с бурята-
Таблица 5. Генетические расстояния между древними популяциями Чукотки и неолита Прибайкалья
|
Группа |
Китой |
Серово |
Глазково |
Неолит Прибайкалья (средние) |
|
Уэлен |
0,035 |
0,047 |
0,051 |
0,037 |
|
Эквен |
0,041 |
0,050 |
0,038 |
0,036 |
|
Древняя Чукотка (средние) |
0,032 |
0,043 |
0,039 |
0,031 |
ми, тувинцами и монголами. Аналогичные особенности алеутов выявляются и по фенетическим данным. В пространстве канонических переменных они располагаются гораздо ближе к тувинцам и эвенкам, чем к чукчам и эскимосам (см. рис. 1, 5).
Если исходить из предположения о том, что заселение Чукотки шло из Центральной Сибири, то особенности алеутов можно объяснить сохранением в условиях изоляции характерных черт протоэскимо-со-алеутского типа, близкого к типу древних обитателей Центральной Сибири. По мнению Т.Я. Токаревой, отметившей сходство алеутов с эвенками и неолитическим населением Прибайкалья, «исходной стадией расового типа алеутов был расовый тип, известный в Прибайкальском неолите и распространенный в некую отдаленную эпоху на огромной территории, включавшей бассейн Берингова моря и, может быть, северо-западную Америку» [1937, с. 58]. Поддерживая эту точку зрения, В.П. Алексеев писал: «...есть все основания предполагать, что морфофизиологическое своеобразие алеутов может найти объяснение в рамках гипотезы консервации протомонголоидных особенностей» [Алексеев, Трубникова, 1984, с. 75]. Действительно, алеуты довольно близки к населению неолита Прибайкалья с учетом усредненных характеристик неолитических групп ( d = 0,036).
При включении в канонический анализ средних характеристик древних популяций неолитическое население Чукотки смещается в сторону арктических групп (рис. 6), что соответствует структуре классификационного древа (см. рис. 2). Неолитическое население Прибайкалья передвигается к центру ареала континентальных монголоидов. Алеуты же благодаря присутствию предковой общности неолита Чукотки оказываются вместе с остальными группами арктической расы в области положительных значений второй и отрицательных значений первой канонической переменной, располагаясь в то же время на равном расстоянии от неолитического населения Прибайкалья и центрально-азиатских популяций. Можно заключить, что средние характеристики древних популяций действительно в определенной степени отражают исходные генофонды: протоэскимосоалеутскую общность и предковую общность континентальных монголоидов Сибири. По-видимому, протоэскимосоалеуты,
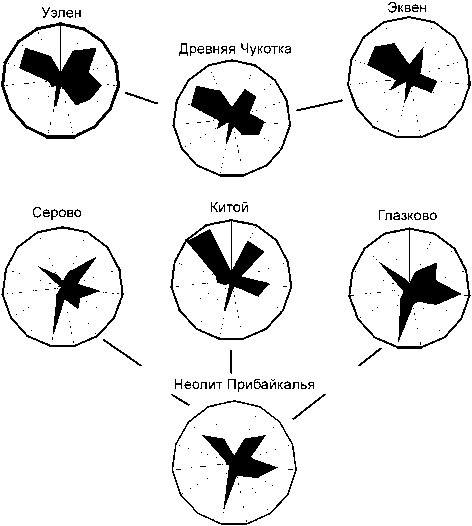
Рис. 4. Сравнение фенетической структуры древних популяций Чукотки и неолита Прибайкалья.
На радиусах по часовой стрелке отложены частоты тех же признаков, что и на рис. 3.
2,5
2,0
i 1,5
I 1-°I °-5
б -0,5
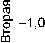
Негидальцы ▲
Ульчи
Монголы
; Чукчи оленные ; Чукчи береговые ; Эскимосы ж
Эквен
Уэлен ▲
Китой х Алеуты А Эвенки
Калмыки „:
Манси А ГлазковоК1чи^цы Тувинцы ш ▲ ▲ Т БурЯТЫ ▲
Серово ▲ I ▲
Ханты ! Теленгеты ▲
Койбалы А Бельтыры ; ▲
; СагайцыА
-1,5
-2,0
-2,0 -1,5 -1,0 -0,5 0 0,5 1,0 1,5
Первая каноническая переменная
Рис. 5. Результаты канонического анализа современных и древних популяций Сибири.
2,0
1,5
1,0
0,5
Чукчи оленные - ▲
Эскимосы
Чукчи береговые
Древняя Чукотка
* -1,0-
£ -1,5 -
-2,0
Шорцы
Сагайцы
Алеуту Бельтыры
* Тувинцы ▲ Теленгеты Буряты *
Неолит Прибайкалья Негидальцы
Ульчи
Эвенки
Мрнголы
Кой балы ‘■Качинцы ж
Ханты
Манси
! Калмыки
-2,51---------!---------I—
-2,0 -1,5 -1,0
О
0,5
1,0
1,5
Первая каноническая переменная
Рис. 6. Результаты канонического анализа современных и древних популяций Сибири с учетом усредненных характеристик неолитических групп.
равно как и популяции неолита Прибайкалья, являлись потомками первоначально единого по своей генетической структуре протомонголоидного палеолитического населения Сибири, создававшего локальные неолитические культуры в процессе освоения обширных территорий Северной Азии.
Полученные результаты находят подтверждение в данных археологии, согласно которым в конце IV тыс. до н.э. начали интенсивно расширяться границы байкальско-ленского ареала. Ленско-алданская неолитическая культура распространялась через верховья Лены, по верхнему течению Ангары вниз и далее на северо-запад и север, к Хатанге и низовьям Лены, а также на восток по Охотскому побережью к северо-восточной оконечности Азиатского материка [Чернецов, 1973].
Исследованиями А.П. Окладникова [1948] установлено наличие культурных связей Прибайкалья в неолитическое время с далекими районами Запад-
ной Сибири и с областями к западу от Урала. Черты сходства с прибайкальским неолитом исследователь находит и в неолитических культурах Енисея, связывая это с распространением на юг и на запад групп ранненеолитического населения Прибайкалья. Более того, отмечаются признаки широких контактов носителей именно китойской культуры с обитателями довольно отдаленных районов как на западе, так и на востоке [Окладников, 1974]. Из Прибайкалья по Лене шло, по-видимому, заселение территории Северной Якутии, где неолитические памятники обнаруживают явное сходство с прибайкальскими. В свою очередь, неолит Якутии оказал большое влияние на неолит Чукотского полуострова, и это в значительной степени обусловило характер последующих культур Чукотки, связанных с этногенезом северо-восточных палеоазиатов [Диков, 1974].
В то же время нельзя игнорировать возможность сохранения южного и тихоокеанского компонентов у представителей арктической группы. Рядом авторов высказывалось предположение о том, что именно через Берингию происходило проникновение тихоокеанских элементов в Америку [Зубов, 2002; Neves et al., 2003]. Мы провели сравнение современного и древнего населения Чукотки с южными и тихоокеанскими монголоидами (табл. 6). У современных арктических групп на нашем материале южная примесь не выявляется, но древние популяции Чукотки обнаруживают определенную близость к бирманцам и папуасам. Примечательно, что тихоокеанский комплекс был выявлен у древних берингоморцев и по альтернативной системе краниоскопических признаков [Козинцев, 1988]. Следовательно, образование протоэскимосоалеутской этнокультурной общности могло происходить на территории Чукотки в процессе культурной и биологической адаптации к экстремальным условиям Берингии переместившихся сюда групп как внутриконтинентального населения Северной Азии, так и мигрантов из Южной Азии и Тихоокеанского региона.
Таблица 6. Генетические расстояния между арктическими популяциями Сибири, южными монголоидами и тихоокеанскими группами
|
Группа |
Эскимосы |
Чукчи |
Алеуты |
Уэлен |
Эквен |
Неолит Чукотки (средние) |
|
|
береговые |
оленные |
||||||
|
Бирманцы |
0,109 |
0,106 |
0,130 |
0,090 |
0,065 |
0,063 |
0,058 |
|
Индийцы |
0,187 |
0,183 |
0,191 |
0,130 |
0,129 |
0,126 |
0,120 |
|
Малайцы |
0,154 |
0,107 |
0,132 |
0,102 |
0,107 |
0,123 |
0,109 |
|
Австралийцы |
0,268 |
0,285 |
0,303 |
0,192 |
0,194 |
0,138 |
0,164 |
|
Папуасы Муруа |
0,105 |
0,088 |
0,113 |
0,077 |
0,057 |
0,073 |
0,059 |
|
Папуасы Авайама |
0,122 |
0,132 |
0,145 |
0,098 |
0,084 |
0,082 |
0,078 |
|
Папуасы (средние) |
0,109 |
0,105 |
0,125 |
0,083 |
0,066 |
0,072 |
0,064 |
|
Меланезийцы |
0,250 |
0,228 |
0,260 |
0,178 |
0,188 |
0,152 |
0,171 |
