Палеогенетическое исследование древнего населения Горного Алтая

Автор: Чикишева Т.А., Губина М.А., Куликов И.В., Карафет Т.М., Воевода М.И., Ромашенко А.Г.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология
Статья в выпуске: 4 (32), 2007 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14522617
IDR: 14522617 | УДК: 572.08+575.174:599:9
Текст статьи Палеогенетическое исследование древнего населения Горного Алтая
Население любого региона развивается в значительной мере при его взаимодействии с ближайшими соседями, а нередко и при участии групп, значительно удаленных географически, но вовлеченных в этногенез событиями разного характера: массовыми миграциями вследствие изменений экологии среды обитания, перераспределением геополитического влияния и связанной с ним структуры межэтнических отношений, трансформациями в сфере материальной и духовной культуры древних обществ и т.д.
В исторической ретроспективе мы наблюдаем смену культур и народов на одной и той же территории. Объективный подход к истории населения любого региона требует анализа преемственности всех структурных элементов многокомпонентных в своей основе этнокультурных образований, в т.ч. и генетической преемственности. Этот аспект проблемы долгое время решался методами физической антро- пологии, которые позволяли извлекать генетическую информацию только косвенным путем. Современный этап развития биологической науки предоставляет возможность изучать генетическую историю популяций посредством ДНК, выделяемой из палеоантропологического материала. Исследование структуры генофондов путем анализа последовательностей ДНК дает более детальное представление о происхождении популяций.
Теперь уже принято за правило историю освоения человеком любого участка ойкумены решать совместными усилиями археологов, физических антропологов и генетиков. Физическая антропология позволяет провести сканирование морфологических особенностей всего предоставляемого археологами массива останков но сителей древних культур и выделить узловые эпизоды в реконструируемых процессах формирования их антропологического состава. Это помогает сориентировать трудоемкий генетический анализ таким образом, чтобы при минимальных затратах могла быть извлечена достаточная информация для определения этнокультурного пространства, в котором у разных групп населения обнаруживаются
общие генетические маркеры, свидетельствующие об их родстве, прослеживаемом при несходстве элементов культурного комплекса.
Настоящее исследование выполнено на палеоантропологических материалах, полученных из погребальных пямятников нескольких археологических культур, существовавших на территории Горного Алтая в хронологическом диапазоне от эпохи неолита (IV тыс. до н.э.) до рубежа новой эры. Оно проведено методами физической антропологии (краниометрии) и молекулярной генетики (анализа ГВС I контрольного района митохондриальной ДНК) и имеет цель – выяснить основные векторы генетических связей у представляемого ими населения.
Ранее таким же комплексом методов нами было проведено изучение носителей пазырыкской культуры скифского времени (IV–III вв. до н.э.), позволившее выявить их антропологический состав, обсудить варианты формирования выделенных компонентов и наметить перспективную линию преемственности генотипов пазырыкцев в группах южных самодийцев [Чикишева, 2003б; Воевода и др., 2003]. Из блока полученных результатов мы выбрали для дальнейшей разработки вопросы ретроспективного плана, связанные с генетическими истоками пазырыкских групп.
Пазырыкский палеоантропологический материал происходит из нескольких могильников, локализованных в долинах основных рек Горного Алтая. Изучение особенностей изменчиво сти признаков морфологического комплекса в краниологических сериях, сформированных в соответствии с приуроченностью погребений к определенным речным долинам, дает нам право рассматривать данные серии как выборки из популяций [Чикишева, 2000а, 2003б]. Поскольку палеогенетический анализ был проведен нами ранее только в одной пазырыкской популяции – осваивавшей долину р. Ак-Алаха на плоскогорье Укок, мы сочли необходимым начать исследование генетического материала в других палеопопуляциях. В связи с этим были изучены образцы митохондриальной ДНК в палеоантропологической серии из могильника Уландрык-1, расположенного в долине одноименной реки.
Для ретроспективного изучения генетических связей пазырыкцев мы провели исследование митохондриальной ДНК из костных образцов, происходящих из погребений предшествующих культур: неолитической (IV тыс. до н.э.) и каракольской периода развитой бронзы (первая половина – середина II тыс. до н.э.). Следует отметить, что формирование данной выборки определялось двумя задачами. Первая заключается в проверке на генетическом уровне гипотезы об автохтонном происхождении одного из основных компонентов антропологического состава носителей пазырыкской культуры. Он выделен нами на краниологическом уровне [Чикишева, 1994, 1996, 2000а, 2002, 2003б] и может быть охарактеризован как антропологический тип с промежуточным выражением о сновных черт, дифференцирующих монголоидную и европеоидную расы. Индивиды с такой морфологией преобладают в рядовых погребениях пазырыкской культуры [Чикишева, 2003б]. Сравнительный краниологический анализ дал основание предполагать их генетическую связь с людьми эпохи неолита и носителями каракольской культуры эпохи бронзы Горного Алтая. Вторая задача работы связана с разработкой гипотезы, предложенной одним из авторов этой статьи, о протоморфности данного морфологического комплекса и независимости его генезиса от метисации европеоидов и монголоидов [Чикишева, 2000б, 2003б].
Итак, в настоящем исследовании проведено изучение гаплотипиче ского разнообразия ГВС I контрольного района мтДНК из ко стных образцов неолитической эпохи (IV тыс. до н.э.) и из погребений каракольской культуры периода развитой бронзы (первая половина – середина II тыс. до н.э.), а также сопоставление его с таковым для останков более поздней пазырыкской культуры (IV–III вв. до н.э.).
Материал и методы
Выборка включает 11 образцов из погребений различных могильников, расположенных друг от друга на расстоянии от нескольких десятков до нескольких сотен километров в Республике Алтай (рис. 1, табл. 1).
По ряду краниологических признаков высокой дифференцирующей значимости многие скелеты, составляющие данную выборку, характеризуются чертами своеобразной промежуточности своих фенотипических параметров по сравнению с представителями европеоидной и монголоидной общностей (основных рас). Это признаки общей уплощенности лицевого скелета и его отдельных структур: назома-лярный и зигомаксиллярный углы горизонтального профиля, угол выступания носовых костей, симоти-ческий и дакриальный указатели выступания переносья (табл. 2).
Выделение тотальной ДНК из кости. Для каждого образца процедуру выделения ДНК из костной ткани повторяли несколько раз. Фрагмент губчатой кости предварительно обжигали в пламени спиртовки и в течение 1 ч облучали ультрафиолетом с каждой стороны, затем размалывали в мелкодисперсный порошок. Для извлечения тотальной ДНК к 1 г костного материала добавляли 4 мл лизирующего буфера (4М гуанидинтио-ционат: 0,1М NaCl, 0,014M β-меркаптоэтанол, 0,025М ЭДТА, 0,5% SDS) и инкубировали 12 ч при температуре 60 оС. Экстракцию ДНК проводили двукратно фенол-
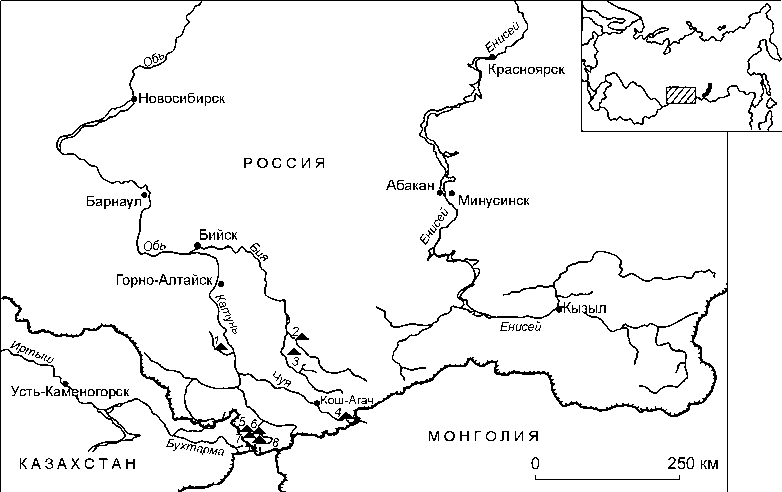
Рис. 1. Локализация памятников, из которых происходят образцы.
1 – Каракол; 2 – Каракол-1; 3 – Каминная пещера; 4 – Уландрык; 5 – Бертек; 6 – Ак-Алаха;
7 – Мойнак; 8 – Кутургунтас.
Таблица 1. Географическая локализация и культурно-хронологическая характеристика образцов
Выделение митохондриальной ДНК из пула тотальной ДНК с использованием магнитных частиц. Отжиг биотинилированного праймера (pr79-Б 16024-ttctttcatggggaagcagattt-16046 или pr80-Б 16422- attgatttcacggaggatggtg-16401) в концентрации 1 мкМ с комплементарным фрагментом первого гипервариабельного сегмента (HVS-1) контрольного района мтДНК проводили в буфере, содержащем 75мM Трис-HCl (pH 9,0), 20мM (NH4)2SO4, 0,01% Тween-20. Процедура включала 10 мин денатурации при температуре 95 оС и 60 мин отжига при 55 оС. Затем комплекс праймер–матрица (мтДНК) переосаждали ПЭГ для удаления праймеров, не связанных с целевой
Таблица 2. Сопоставление параметров краниологических признаков останков из палеовыборки с их предельными значениями в европеоидной и монголоидной антропологических общностях
|
Индивид * |
Пол |
Назома-лярный угол |
Зигомак-силярный угол |
Угол выступления носовых костей |
Симоти-ческий указатель |
Дакри-альный указатель |
Антропологический тип ** |
|
1 |
Жен. |
145,1 |
134,7 |
25 |
46,4 |
50,8 |
Промежуточный |
|
2 |
Муж. |
– |
– |
– |
– |
– |
» |
|
3 |
» |
136,4 |
– |
– |
– |
– |
» |
|
4 |
» |
144,3 |
134,9 |
– |
– |
– |
» |
|
5 |
» |
– |
– |
– |
31,07 |
– |
Монголоидный |
|
6 |
Жен. |
– |
124,9 |
30 |
56,59 |
53,59 |
Европеоидный |
|
7 |
Муж. |
143,9 |
– |
– |
– |
– |
Промежуточный |
|
8 |
» |
147 |
128 |
– |
– |
– |
» |
|
9 |
» |
– |
122,2 |
– |
39,22 |
– |
» |
|
10 |
» |
– |
– |
30 |
37,86 |
46,43 |
Европеоидный |
|
11 |
» |
138,3 |
127,9 |
20 |
64 |
66,67 |
» |
|
Предельные значения (min–max) параметров признака в европеоидной антропологической общности *** |
– |
135–139 |
125–130 |
25–37 |
39,8–49,3 |
35–42 |
– |
|
Предельные значения (min–max) параметров признака в монголоидной антропологической общности *** |
– |
145–148 |
137–142 |
7,5–19 |
19,5–29 |
51–73 |
– |
*Номера в соответствии с табл. 1.
**Антропологический тип диагностирован по опубликованному описанию индивидов и краниометрическим характеристикам [Чикишева, 2000а, 2003б].
***Данные взяты из таблиц краниометрических констант [Алексеев, Дебец, 1964] без учета редко встречающихся очень малых и очень больших значений признаков.
ДНК. Для этого к полученному раствору добавляли равный объем 20% ПЭГ (м.в. 6000) в 2,5М NaCl; смесь инкубировали при температуре 37 оС в течение 15 мин; осаждение осуществляли центрифугированием при 12 000 об./мин в течение 15 мин. Осадок трижды промывали 700 мкл 80% этанола, высушивали при температуре 56 оС и растворяли в 35 мкл деионизованной воды.
Для уменьшения неспецифической сорбции магнитные частицы перед применением инкубировали в пяти объемах раствора Денхарда. К комплексу биотинилированный праймер – матрица (мтДНК) добавляли равный объем 20× SSC и 10 мкл парамагнитных частиц в 10× SSC. Ковалентное связывание иммобилизованного на частицах стрептавидина с биотинилированными праймерами проводили при комнатной температуре и постоянном помешивании в тече- ние 1 ч. Отделение парамагнитных частиц с иммобилизованными на них праймерами в комплексе с мтДНК от остальных компонентов (пул геномной ДНК, примеси-ингибиторы ПЦР) осуществляли при помощи магнитного планшета по протоколу фирмы-изготовителя (фирма “MERCK” для BioBeads Streptavidin).
Элюцию требуемых для анализа фрагментов мтДНК проводили в 25 мкл деионизованной воды путем денатурации комплекса праймер–мтДНК при температуре 95 оС в течение 5 мин. Элюат отделяли от магнитных частиц при помощи магнитного планшета при визуальном контроле их фиксации на стенках пробирки. Далее ДНК, извлеченную в раствор, использовали в качестве исходной матрицы в полимеразной цепной реакции.
Амплификация древней ДНК. Ее проводили методом nested-PCR. Для nested-PCR фрагмента HVS-1
контрольного района мтДНК использовали две пары праймеров: 1) наружные (фрагмент 398 н.п.) – pr79 16024-ttctttcatggggaagcagattt-16046 и pr80 16422-attgatttcacggaggatggtg-16401; 2) внутренние (фрагмент 336 н.п.) – pr_re79 16052-ccacccaagtattgactcaccc-16073 и pr_re80 16388-ctatctgaggggggtcatccat-16367. Для амплификации наружного фрагмента HVS-1 контрольного района мтДНК ПЦР-смесь объемом 12,5 мкл включала: 75мM Трис-HCl (pH 9,0), 20мM (NH4)2SO4, 0,01% Тween-20, 1мкM pr79 и pr80, 0,6мМ каждого из четырех dNTP, 5мM MgCl2, 3 ед. Тag полимеразы и 3 мкл элюированной ДНК. Циклы амплификации, числом 33, включали денатурацию в течение 1 мин при температуре 95 оС, отжиг в течение 1 мин при 61 оС и синтез в течение 1 мин при 72 оС.
Амплификацию внутреннего фрагмента HVS-1 контрольного района мтДНК проводили в 50 мкл реакционной смеси, содержащей 75мM Трис-HCl (pH 9,0), 20мM (NH4)2SO4, 0,01% Тween-20, 1мкM pr_re79 и pr_re80, 0,6мМ каждого из четырех dNTP, 5мM MgCl2, 5 ед. Тag полимеразы и 3 мкл амплифика-та наружного фрагмента HVS-1 контрольного района мтДНК. Циклы амплификации, числом 25, включали денатурацию в течение 25 с при температуре 95 оС, отжиг в течение 25 с при 55 оС и синтез в течение 25 с при 72 оС. Детекцию ПЦР-продукта проводили методом гель-электрофореза в 4% полиакриламидном геле с последующей окраской бромистым этидием. Далее к амплификату добавляли равный объем 20% ПЭГ (м.в. 6000) в 2,5М NaCl, инкубировали при температуре 37 оС в течение 15 мин, осаждали центрифугированием при 12 000 об./мин 15 мин и трижды промывали 700 мкл 80% этанола при центрифугировании (12 000 об./мин) 15 мин. Полученный осадок высушивали при температуре 56 оС и растворяли в 25 мкл деионизованной воды.
Для секвенирующей реакции использовали праймеры pr_re79 и pr_re80 и BigDye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems). Образцы ДНК анализировали на автоматическом секвенаторе ABI Prism 310 (Applied Biosystems).
Результаты
Исследование структуры последовательностей нуклеотидов ГВС I контрольного района мтДНК 11 образцов древней ДНК (табл. 3) выявило наличие у них девяти гаплотипов, которые в соответствии с общепринятой классификацией [Wallace, 1995; Richards et al., 1998; Kivisild et al., 2002] относятся к пяти гап-логруппам: трем восточно-евразийским – А, С, D и двум западно-евразийским – U5 и H. Для двух образцов ДНК из Каминной пещеры (№ 1) и Ак-Алаха-5, кург. 4 (№ 8) необходимы дополнительные исследования для уточнения гаплогрупп.
Большая часть анализируемых образцов мтДНК имеет западно-евразийские гаплогруппы (63,6 %). Среди них преобладает гаплогруппа Н (57 %), три гаплотипа которой являются CRS-последовательностями, а один вариант имеет замену в позиции 16304.
При сопоставлении данных табл. 2 и 3 выяснилось, что только в случае образцов № 5, 10 и 11 мы наблюдаем прямую корреляцию между расоспеци-фичностью краниологических признаков и принадлежностью гаплотипов их мтДНК к западно- или восточно-евразийским гаплогруппам. Погребенный в кург. 5 Ак-Алаха-5 (№ 5) с генотипом мтДНК восточно-евразийской гаплогруппы А характеризуется типичным монголоидным морфотипом; двое других (№ 10, 11) с гаплотипом CRS европеоидной гапло-группы Н имели европейские черты. Все осталь-
Таблица 3. Гаплотипическое разнообразие ГВС I мтДНК древнего населения Алтая в различные эпохи
|
№ п/п |
Гаплотип |
Гаплогруппа |
Кол-во повторов |
Погребение |
|
1 |
16224 |
H/K |
4 |
Каминная пещера |
|
2 |
16192-16256-16270 |
U5 |
3 |
Каракол-1 (1982 г.) |
|
3 |
16304 |
H |
2 |
Каракол (1985 г.) |
|
4 |
CRS |
H |
3 |
Бертек-56 |
|
5 |
16223-16242-16290-16319 |
A |
3 |
Ак-Алаха-5, кург. 5 |
|
6 |
16093-16129-16223-16298-16327 |
C |
3 |
Ак-Алаха-1 |
|
7 |
16223-16239-16319-16362 |
D |
3 |
Ак-Алаха-5, кург. 3 |
|
8 |
16129-16182C-16183C-16189-16362 |
U5/J |
2 |
То же, кург. 4 |
|
9 |
16223-16311-16316-16362 |
D |
3 |
Мойнак-2 |
|
10 |
CRS |
H |
4 |
Кутургунтас-1 |
|
11 |
CRS |
H |
3 |
Уландрык-1 |
ные останки (за исключением образца № 6) имеют промежуточные между монголоидами и европеоидами антропометрические показатели независимо от расоспецифичности гаплогрупп, к которым принадлежат гаплотипы их мтДНК (см. табл. 3). В случае образца № 6 (женщина), не смотря на европеоидность морфотипа по краниологическим параметрам, гаплотип мтДНК отно сится к гапло-группе С, входящей в состав супергаплогруппы М, варианты которой распространены среди типичных монголоидов.
Анализ распространенности девяти гаплотипов древних останков (см. табл. 3) среди современных популяций человека, заселяющих Горный Алтай и сопредельные территории, показал присутствие только четырех из них в существующих базах данных. Гаплотип 16192-16256-16270 (U5) был выявлен у тувинцев (0,3 %) и хантов (0,4 %); с заменой в позиции 16304 (H) – у тувинцев (3,5 %); CRS (H) – у тувинцев (2,1 %), хакасов (1,1 %), алтайцев (1,1 %), казахов (4,1 %), хантов (1,6 %) и манси (5,3 %); 1609316129-16223-16298-16327 (С) – у тувинцев (3,5 %), алтайцев (1,1 %) и манси (2,6 %).
Гаплотип с заменой в позиции 16224 широко представлен в популяциях Западной Европы, у населения Хорватии и у русских. Гаплотип 16223-16242-1629016319 гаплогруппы А (№ 5) не обнаружен в популяциях Центральной Азии и финно-угров, но выявлен у селькупов с частотой 3,3 % (неопубликованные данные М.А. Губиной, Л.П. Осиповой). В Центральной Азии распространены два других – 16223-1629016319-16362 (хакасы – 1,1 %) и 16242-16290-16293С-16319 (тувинцы – 1,2 %).
Два гаплотипа гаплогруппы D – 16223-1631116316-16362 (№ 9) и 16223-16239-16319-16362 (№ 7) (см. табл. 3) – в современных популяциях не обнаружены, но их производные зарегистрированы у населения Центральной и Юго-Восточной Азии. Гаплотип 16129-16182C-16183C-16189-16362 (№ 8) отсутствует в базах данных; близкие к ним 1605116092-16129-16182-16183-16189-16270-16362 и 16069-16182-16183-16189-16190-16362, принадлежащие гаплогруппам U5b1 и J соответственно, выявлены у англичан (0,7 %) [Helgason et al., 2001].
Обсуждение результатов
В историко-культурных исследованиях физико-географическая Алтае-Саянская горная страна (та ее часть, которая лежит на территории России) произвольно рассматривается иногда как особый регион Южная Сибирь, а иногда Горный Алтай, Кузнецкий Алатау, Кузнецкая котловина и Салаирский кряж включаются в состав Западной Сибири.
Западная и Южная Сибирь расположены между ареалами западного и восточного круга основных антропологических общностей – европеоидной и монголоидной. В антропологической литературе оба региона также часто определяются как переходные зоны, соответственно северная и южная. Под переходностью подразумевается географически детерминированная и на отдельных этапах истории реализованная возможность контактов и перемещений людей, характеризующихся контрастными физическими признаками европеоидного и монголоидного расовых комплексов. Именно это обстоятельство стало одной из главных причин доминирования концепции метисации европеоидов и монголоидов в решении вопросов происхождения антропологического своеобразия носителей практически всех археологических культур, открытых на территории Горного Алт ая. Однако полученные в по следние годы палеоантропологические материалы позволяют рассматривать морфологические особенности хотя бы части этого населения как результат сохранения определенной протоморфности, заключающейся в некоординированном сочетании важнейших расодифференцирующих краниологических признаков. Предполагаемые протоморфные морфологические комплексы в группах древнего населения Горного Алт ая характеризуются большой шириной лица, его равномерной уплощенностью на уровне орбит и скуловых костей, высоким переносьем, несильным выступанием над общей линией профиля лица носовых косточек, средней шириной носового отверстия, широкими и невысокими орбитами. Отнести такой комплекс к кругу монголоидных или европеоидных антропологических типов затруднительно. Он чрезвычайно устойчив на территории Горного Алтая, обнаруживая себя практически в неизменном виде в антропологическом составе групп носителей нескольких культур от эпохи неолита до раннего железного века включительно. Мы проанализировали палеоантропологические материалы, представляющие это население, с двух позиций: по результатам традиционного краниометрического анализа и особенностям вариации структуры митохондриальной ДНК.
Краниометрические данные широко используются при исследовании происхождения и процессов формирования древних и современных групп населения с середины XIX в., и их интерпретационные возможности хорошо известны любому специалисту в области генезиса культур и народов. Иная ситуация наблюдается в настоящее время вокруг данных молекулярной палеогенетики, делающей первые попытки их систематизации в этнокультурном пространстве человечества.
В начале 1990-х гг. несколькими ведущими группами исследователей был выполнен глобаль-
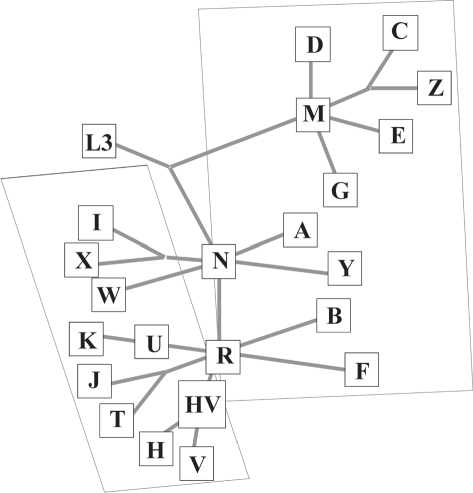
Рис. 2. Схема филогенетического дерева гаплогрупп митохондриальной ДНК народов Евразии [Wаllace, 1995; Kivisild et al., 2002; Бермишева и др., 2002].
Европеоидные гаплогруппы – HV, H, V, J, T, U, K, I, W, X; монголоидные – А, В, E, F, Y, M, C, D, G, Z.
ный скрининг вариабельно сти митохондриального генома в основных расовых группах человека. В результате удалось выявить мутации, являющиеся ключевыми для определения расоспецифичных кластеров, или гаплогрупп мтДНК [Wallace, 1995; Richards et al., 1998].
Данные секвенирования гипервариабельных участков ГВС I, ГВС II и рестрикционного анализа всего митохондриального генома показали, что для европеоидных популяций наиболее характерны гаплогруппы HV, H, V, J, T, U, K, I, W, X [Ibid], а для монголоидных – A, B, E, F, Y, М (C, D, G, Z) (рис. 2) [Wallace, 1995; Kivisild et al., 2002; Берми-шева и др., 2002].
Рассмотрим степень сопряженности полученных нами данных краниометрического и палеогенетичес-кого анализов. Наиболее древние образцы выборки относятся к эпохе неолита. Антропологических находок этой эпохи в Горном Алтае всего две: скелет женщины из погребения в Каминной пещере [Чикишева, 2000в] и скелет мужчины из погребения в пещере Нижне-тыткескенская-1 [Ким, Чикишева, 1995]. Обе могут быть отнесены к одному краниологическому типу, ведущими чертами которого являются мезобрахикрания и средняя высота мозгового отдела черепа, большая ширина и уплощенно сть лицевого отдела, отно си-тельно высокое переносье и среднее выступание носа над линией общего профиля лица. Аналогии такому сочетанию краниологических признаков обнаружены в материале из погребений Красноярско-Канской лесостепи – могильников Базаиха [Алексеев, 1961] и Долгое Озеро [Герасимова, 1964]. Разумеется, сходство не доходит до идентичности, но описанный морфологический комплекс присутствует у всех индивидов. Генезис этого комплекса обсуждался на материале из Базаихи и был охарактеризован как ослабленный монголоидный: В.П. Алексеев [1961] видел природу этой ослабленности в европеоидной примеси, а Г.Ф. Дебец [1948] в равной степени допускал ее протоморфность.
Что касается краниологических материалов эпохи неолита из предгорной зоны Алтая, то они обнаруживают сложный антропологический состав. Серия из могильника Солонцы-5 демонстрирует сочетание особенностей упомянутого выше компонента неолитического населения Алтае-Саянского нагорья и монголоидного компонента популяций Прибайкалья той же эпохи (в наибольшей степени тяготеющего к представленному в серовской краниологической серии из бассейна р. Лены) [Кунгурова, Чикишева, 2002]. Серии из могильников Усть-Иша и Иткуль представляют собой вариант метисации европеоидов древнего гиперморфного восточно-средиземноморского типа, чье происхождение связано с южными районами Средней Азии, и монголоидов палеосибирского типа из Прибайкалья [Дрёмов, 1980]. Таким образом, для неолитического населения предгорий Алтая обоснован вариант генезиса на основе смешения монголоидных и европеоидных антропологических компонентов.
В период ранней бронзы (конец IV – начало II тыс. до н.э.) на территории Горного Алтая существовали две культуры – афанасьевская (представленная большим числом археологических памятников и хорошо изученная в антропологическом отношении) и большемысская (представленная только керамическими комплексами в отдельных поселениях, с полным отсутствием каких-либо сведений о людях). Ареал афанасьевской культуры включает также Минусинскую котловину, северо-западную часть Монгольского плато и Турфанскую котловину в Синьцзяне. Афанасьевцы по комплексу антропологических признаков почти тождественны носителям синхронной ямной культуры Восточно-Европейской равнины. По вопросу о происхождении афанасьевского населения имеется несколько гипотез. В первых антропологических исследованиях морфологическое сходство афанасьевцев с населением далекого Запада (но сителями ямной культуры) объяснялось переселением в Южную Сибирь и Центральную Азию больших масс людей с территории основного ареала расселения европеоидов [Дебец, 1948; Алексеев, 1961]. Позднее была выдвинута гипотеза о возможности существования в восточных районах степной полосы Евразии в доафанасьевское время самостоятельного очага формирования одного из локальных вариантов европеоидной расы, с которым был связан генезис носителей афанасьевской культуры [Алексеев, 1989, с. 350–355].
При планировании нашего исследования мы предполагали изучение мтДНК в образцах, относящихся к неолитическому населению предгорной зоны Алтая и к афанасьевцам Горного Алтая. К сожалению, на данный момент выделить ДНК и генотипировать эти образцы в полном объеме не удалось. Мы продолжаем работать с ними.
В период развитой бронзы между XVIII–XVII и X вв. до н.э. на территории Горного Алтая существовали культуры, самобытность и яркость которых проявляется в великолепных образцах первобытного искусства – полихромных рисунках, выполненных на каменных плитах саркафагов и на скалах. Археологи пока точно не установили, к единой или разным культурам относятся все погребальные и поселенческие комплексы этого хронологического периода. Однозначно выделяется каракольская культура [Молодин, 1991, 2002, 2006], памятники которой (Озерное, Каракол, Каракол-1, Беш-Озек, Усть-Куюм, Кара-Коба-1) локализованы в центральной части Горного Алтая. Они имеют многоплановые параллели с памятниками окуневской культуры Минусинской котловины, а также определенное сходство с синхронными памятниками кротовской культуры лесостепного междуречья Оби и Иртыша.
В палеоантропологическом материале каракольской культуры мы обнаружили аналогии краниологическому типу неолитических черепов, что дало нам основание предполагать генетическую связь обоих культурных образований. Эти материалы получены из двух могильников, раскопанных разными исследователями (А.П. Погожевой [1984] и В.Д. Кубаревым [1988]) в центральном и северо-западном районах Алтая (Онгудайском и Усть-Канском по административному делению), но имеющих одинаковое название – Каракол. Они исследованы и опубликованы [Чикишева 2000в, 2003б]. Черепа из обоих памятников имеют большое морфологическое сходство. Характерный для них антропологический комплекс сближает их как с окуневскими из Минусинской котловины, так и с более ранними рубежа неолита из пещер Горного Алтая, а также из Базаихи и Долгого Озера на среднем Енисее.
На базе материалов двух других памятников примерно того же хронологического периода, открытых на юге Горного Алтая (культовый комплекс Кучерла-1 и курган Бертек-56), В.И. Молодин допускает возможно сть существования особой археологической культуры, предполагая, что в основе орнаментации ее керамики лежит афанасьевская традиция [2002]. Од- нако исследование скелетных остатков мужчины 30– 35 лет и ребенка 6,5–7 лет из кургана Бертек-56 [Чики-шева, 2003б] показало, что европеоидный компонент, обнаруженный у этих субъектов, не связан с афанасьевской антропологической средой. Морфологическая специфика погребенных, возможно, сформировалась в результате метисации антропологического типа, свойственного носителям каракольской культуры (об этом могут свидетельствовать строение лобной кости, горизонтальная уплощенность лицевого скелета, особенности строения зубов), и европеоидного варианта, характеризующегося общей массивностью, широким и очень высоким лицом. Среди европеоидных популяций краниологический вариант с такими особенностями известен по материалам могильника Ранний Тулхар, представляющим скотоводческие племена Бешкентской долины (Таджикистан) II тыс. до н.э. [Кияткина, 1976].
Период поздней бронзы (X–VIII вв. до н.э.) представлен в Горном Алтае очень незначительными материалами, среди которых отсутствуют какие-либо антропологические находки. Это изделия из бронзы, керамика, оставленные населением предалтайских и западно-сибирских лесостепей. Какие-либо местные культурные образования пока не выявлены. Полагают, что морфологический комплекс эпохи поздней бронзы соответствует таковому но сителей карасукской культуры Минусинской котловины и культур безве-щевых погребений Тувы и Монголии.
В культурно-хронологическом интервале эпох неолита – бронзы молекулярно-генетический анализ мтДНК был проведен для образцов № 1–4 (см. табл. 3). Гаплотип образца № 1 (женщина, неолитическое погребение в Каминной пещере) содержит замену в позиции 16224, характерную для вариантов западно-евразийских гаплогрупп Н и К*. Гаплотипы образцов № 2, 3 (погребения каракольской культуры середины II тыс. до н.э.) и 4 (погребение Бертек-56, датируемое началом II тыс. до н.э.) также относятся к западно-евразийским гаплогруппам – U5 и H. Представляет интерес тот факт, что, по данным некоторых исследований, гаплогруппа Н возникла на Алтае на рубеже среднего и верхнего палеолита (примерно 50 тыс. л.н.) и далее широко распространилась в Европе после ледникового периода [Loogvali et al., 2004]. Гаплотип с единичной заменой в позиции 16304 (образец № 3) представлен по всему миру, хотя в популяциях Центральной Азии этот вариант выявлен только у тувинцев (3,5 %). Варианты гапло-группы U5, к которой принадлежит гаплотип образца № 2, встречаются во многих североевропейских популяциях и в наибольшей степени у саамов [Lahermo et al., 1999; Torroni et al., 1993; Wallace et al., 1999]. Эта гаплогруппа входит в состав суперкластера U, возникшего на Евразийском континенте и претерпевшего существенную филогенетическую дифференциацию и географическую дисперсию. Так, гаплогруппа U6 обнаружена на Африканском континенте, U7 типична для популяций Иордании, Кувейта, Ирана, Саудовской Аравии, а U1–U5 выявлены в Западной Европе [Richards et al., 1996, 1998]. Важно отметить, что наиболее древние носители анализируемых образцов мтДНК (№ 1–4), принадлежащих к западно-евразийским гаплогруппам H и U5 (см. табл. 3), имеют промежуточные краниологические параметры (см. табл. 2).
Эпоха раннего железа (ранних кочевников, или скифское время, – VIII–II вв. до н.э.) изучена в Горном Алтае наиболее полно по сравнению с другими археологическими периодами. Имеются основания для культурно-хронологического разграничения памятников VIII (или конца IX) – начала VI в. до н.э. и конца VI – начала II в. до н.э. Первые выделяются исследователями в особую культуру [Могильников, 1983; Степанова, 1986; Кирюшин, Тишкин, 1997]. Для нее пока не подобран термин, а также не решен вопрос о ее происхождении. Предполагается, что население раннескифского времени Горного Алтая развивалось на той же основе, что и носители алды-бельской культуры Тувы и некоторые группы саков Казахстана, а общие истоки формирования их погребального обряда (как одного из ведущих элементов культурного комплекса) находятся в Северо-Западной Монголии [Кирюшин, Тишкин, 1997; Савинов, 1994]. Палеоантропологический материал из раннескифских курганов относительно немногочислен и чрезвычайно смешан: выделяются монголоидные, европеоидные, а также и монголоидно-европеоидные комплексы признаков [Тур, 1997].
По отношению к памятникам следующего хронологического этапа (конца VI – начала II в. до н.э.) в археологической литературе распространено понятие “пазырыкская культура”. В последние три десятилетия были также открыты две группы памятников, синхронные пазырыкским, но отличающиеся некоторыми чертами погребального обряда. На основе одной из них была выделена кара-кобинская культура [Могильников, 1983; Суразаков, 1983]. Другая группа названа чумышско-ишинской. Она немногочисленна, но, тем не менее, ее рассматривают как особую культуру [Суразаков, 1988]. Ряд исследователей, однако, считает неправомерным выделение памятников конца VI – начала II в. до н.э. за рамки пазырыкской культуры [Полосьмак, 1994; Шульга, 1986].
Пазырыкская и кара-кобинская культуры хорошо изучены в антропологическом отношении [Чикишева,
1994, 1996, 2000б, 2002, 2003а]. Преобладающим в составе пазырыкских племен оказался автохтонный морфологический компонент, предположительно восходящий к неолитическому населению Горного Алтая и носителям культур окуневского круга (каракольской). Сравнительный анализ морфологического комплекса ранних кочевников Горного Алтая с опубликованными в литературе данными показал, что этот комплекс доминирует у целого ряда этнокультурных групп скифского времени в южных районах Евразии и обусловливает антропологическое сходство пазы-рыкского населения Южной Сибири с частью племенных объединений сако-усуньской этнокультурной общности Центральной и Средней Азии. Пазырыкцы наиболее близки к предшествующему им на Алтае т.н. раннескифскому населению и тяготеют к сакским и усуньским племенам Джунгарского Алатау и Тянь-Шаня. В антропологическом составе погребенных в курганах кара-кобинского типа, видимо, присутствует компонент, связывающий эту группу кочевников Горного Алтая с племенами лесостепного Обь-Иртыш-ского междуречья.
Палеогенетический анализ антропологических находок, относящихся к пазырыкской культуре, ранее был проведен на репрезентативном материале из курганов в долине р. Ак-Алаха на плоскогорье Укок. В данной работе мы продолжили это исследование с выборкой из шести образцов: № 5–8 из могильника Ак-Алаха, № 9 из кургана Мойнак-2, № 10 из кургана Кутургунтас-1 (см. табл. 3). Кроме того, был проведен анализ образца из кургана в долине р. Уландрык (№ 11) (см. табл. 3). В пазырыкских образцах мы наблюдаем более высокий уровень генетического разнообразия мтДНК, чем в тех, что относятся к предшествующим культурам (см. табл. 3). Помимо западно-евразийских гаплогрупп нами были выявлены три восточно-евразийские – A, D и C (в образцах № 5–7, 9).
Примечательно, что в антропологических материалах пазырыкской культуры гаплотипы гаплогруппы А ранее не встречались [Молодин и др., 2000]. Гаплотип 16223-16242-16290-16319 (образец № 5), несомненно, можно отнести к одному из предковых ее вариантов, встречающихся в настоящее время в популяциях хакасов (гаплотип 16223-16290-16293-16319 – 1,1 %) и тувинцев (гаплотип 16242-16290-16293С-16319 – 1,2 %). Все они являются производными от варианта мтДНК с тремя заменами в ГВС I контрольного района (16223-16290-16319), наивысшая частота которого отмечена в этнических группах Средней Азии и Китая [Wallace et al., 1999]. Этот гаплотип обнаружен у айнов (3,9 %), эвенов (3,1 %), китайцев (7,7 %), корейцев (3,1 %), монголов (2,9 %), уйгуров (7,3 %), казахов (9,3 %), киргизов (6,3 %) и у народов Ближнего Востока (2,7 %) [Comas et al., 1998; Yao et al., 2002; Деренко,
Шилдс, 1997; Derenko et al., 2000]. Внутрипопуля-ционные попарные нуклеотидные различия между близкими к нему последовательностями мтДНК в популяциях Восточной и Средней Азии имеют значения 2,60 и 3,21 соответственно, что указывает в большей мере на среднеазиатское происхождение предкового гаплотипа гаплогруппы А [Молодин и др., 2004]. По некоторым оценкам он возник 48 тыс. л.н. [Иванова, 1993]. Не исключено, что появление среди пазырык-цев носителей вариантов гаплогруппы А связано с миграцией (до IV–III вв до н.э.) групп (или группы) ранних кочевников на территорию Горного Алтая с юго-востока или юга. В пользу этого предположения свидетельствует тот факт, что погребенный в кург. 5 могильника Ак-Алаха-5, мтДНК которого относится к восточно-евразийской гаплогруппе А (образец № 5), имеет четко выраженный монголоидный морфотип (см. табл. 2). Гаплотипы данной гаплогруппы также встречаются в популяциях Волго-Уральского региона, но с низкой частотой [Бермишева и др., 2002].
Два пазырыкских образца (№ 7 и 9) с промежуточными краниологическими признаками (см. табл. 2) являются носителями двух разных вариантов монголоидной гаплогруппы D, гаплотипы которой относительно широко распространены в современных коренных популяциях Алтае-Саянского нагорья и Северной Азии [Richards et al., 1996; Kolman, Sambuughin, Bermingham, 1996; Деренко и др., 2001; Иванова, 1993]. Например, гаплогруппа D представлена у 15 % тувинцев и 7,2 % хакасов [Дамба и др., 2003]. Гаплотип 16223-16311-16316-16362 образца № 9 отсутствует в базе данных мтДНК, но варианты без замен в позициях 16316 или 16311 обнаружены у казахов (1,4 %) и тувинцев (0,3 %). Вариант 1622316239-16319-16362 образца № 7 имеет дополнительную замену в позиции 16239 в сравнении с распространенным среди тувинцев (4,1 %) алтайцев (3,7 %) и казахов (3,4 %) гаплотипом 16223-16319-16362 с тремя заменами. Исходя из полученных данных, можно констатировать, что промежуточные антропометрические признаки, видимо, свойственны носителям гаплотипов как европеоидных, так и монголоидных гаплогрупп (см. табл. 2, 3).
Наиболее впечатляющий результат нами получен для образца № 6 с европеоидными краниологическими чертами, который оказался носителем гаплотипа восточно-евразийской гаплогруппы С (см. табл. 3). Необходимо отметить, что аналогичное несовпадение антропометрических данных и генетических показателей мы уже наблюдали раннее у пазырыкцев, погребенных на могильнике Верх-Кальджин [Воевода и др., 1998], со сходным гаплотипом мтДНК. Варианты гаплогруппы С достаточно широко представлены в современных популяциях региона. У хакасов и тувинцев их частота встречаемости достигает 35 и 40 % соответственно [Дамба и др., 2003], у казахов – 13,8 % (неопубликованные данные). В некоторых типичных монголоидных популяциях (эвенки, юкагиры) гаплотипы гаплогруппы С составляют более 50 % [Schurr et al., 1999; Starikovskaya et al., 1998]. У населения Южной и Западной Европы эта гаплогруппа либо отсутствует, либо частота встречаемости составляет не более 1 %, что характерно и для финно-угров Волго-Уральского региона (от 0 у коми-зырян до 3 % у удмуртов), и для балто-финнских этносов (не превышает 2 %) [Бермишева и др., 2002; Meinila, Finilla, Majamaa, 2001; Villems et al., 1998]. Идентичные гаплотипу 16093-16129-16223-16298-16327 (образец № 6) последовательности мтДНК встречаются и в современных популяциях Горного Алтая, но с низкой частотой: у тувинцев – 3,5 %, алтайцев – 1,1 % [Дамба и др., 2003].
Нам предстоит еще установить, к какой гапло-группе принадлежит западно-евразийский гаплотип 16129-16182С-16183С-16189-16362, обнаруженный у образца № 8 (см. табл. 3). Он отсутствует в базе данных мтДНК, но существует вариант с дополнительной заменой в позиции 16270 у одного англичанина [Helgason et al., 2001]. Древний носитель гаплотипа (образец № 8) имеет промежуточные краниологические признаки, тогда как два других пазырыкца (№ 10 и 11) с западно-евразийскими гаплотипами CRS характеризуются типичными европеоидными антропометрическими параметрами (см. табл. 2, 3).
Заключение
Изучение динамики антропологического со става автохтонного населения Горного Алтая приводит к выводу о преобладании компонента, занимающего промежуточное положение по краниологическим признакам, дифференцирующим расы первого порядка (монголоидную и европеоидную). По результатам молекулярно-генетического анализа мтДНК исследованных образцов, относящихся к эпохам неолита и бронзы, наблюдается присутствие лишь западноевразийских гаплотипов. Однородность структуры генофонда мтДНК на протяжении двух-трех тысячелетий указывает на отсутствие значимого дрейфа генетиче ского материала в популяции. Смешение западных и восточных генных потоков на территории Горного Алтая “летописью” мтДНК зафиксировано только в эпоху железа у носителей пазырыкской культуры. Это подтверждается также комплексом данных по этнокультурогенезу [Молодин, 2003].
Интегральная оценка полученных результатов позволяет заключить, что, по всей видимости, может и не быть строгой корреляции между расоспецифично стью морфологических признаков людей и при- надлежностью гаплотипов их мтДНК к монголоидной или европеоидной гаплогруппе. Краниологические особенности, как и любые другие полигенные признаки, формируются при участии многих локусов ядер-ного генома, кодирующих структурные белки остеогенеза, гормоны, ростовые цитокины и их рецепторы, транскрипционные факторы и другие регуляторные белки. Генетическое содержание наследуемого по материнской линии факультативного материала, каким является митохондриальная ДНК, возможно, только косвенно влияет на процессы морфогенеза. Механизмы согласованного функционирования и эволюции ядерного и митохондриального геномов эукариот в настоящее время не ясны.
Тем не менее полученный нами результат демонстрирует успешность проведения параллельно краниологического и палеогенетического анализов в интерпретации случаев промежуточных краниометрических комплексов. По антропологическим критериям такие комплексы могут быть в равной мере отнесены и к европеоидно-монголоидным метисам, и к специфической антропологической общности, отделившейся от монголоидного или европеоидного расового ствола еще до того периода, когда сформировались ярко выраженные антропологические европеоидные и монголоидные комплексы. Структура основных кластеров мтДНК (гаплогрупп) четко сопряжена с филогенезом популяций, а их подкластеры географически векторизированы. Это качество позволяет нам, обнаружив в митохондриальном геноме автохтонного населения Горного Алтая только западно-евразийские гаплогруппы, сделать вывод о его генетической принадлежности к генному пулу европеоидных популяций. Носители афанасьевской культуры не внесли вклада в антропологический состав этого населения, что не исключает определенного влияния на внешнюю сторону жизни, культуру, хозяйство, верования. Установленное нами отсутствие биологических корреляций между митохондриальным геномом и краниологическими признаками требует проверки выводов, полученных на основе морфологии скелета, критериями генетического уровня.
С большой вероятностью можно предполагать, что европеоидный компонент, обнаруженный в антропологическом составе племен Горного Алтая II тыс. до н.э., свидетельствует о их связях со скотоводческим населением Передней и Средней Азии. Существование таких связей подтверждается археологическими материалами [Молодин и др., 2004; Полосьмак и др., 2006], а также данными палеогене-тического анализа.
Является небезынтересным, что древнее население Алтае-Саянского нагорья преимущественно представлено западно-евразийскими гаплотипами гаплогрупп H и U5, распространенными в Западной
Сибири и Европе. Тогда как в современных популяциях, населяющих этот регион, преобладают восточноевразийские гаплогруппы [Губина и др., 2006].
Учитывая результаты антропологического анализа, извлекающего генетическую информацию хотя и косвенно, но из несопоставимо большего массива древних образцов, можно сделать вывод, что выделенная нами южная евразийская антропологическая формация [Чикишева, 2000а, б; 2003б] является реликтом древней протопопуляции. Эти данные не противоречат высказанной ранее гипотезе о формировании антропологического состава современного населения Евразии на основе исходной протопопуляции, заселявшей предположительно территорию Средней и Центральной Азии [Воевода и др., 2003].
