Палеоэкологические условия обитания поздних неандертальцев на Алтае: по материалам пещер Чагырская и Окладникова

Автор: Сердюк Н.В., Зажигин В.С., Маркин С.В., Колобова К.А.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 1 т.50, 2022 года.
Бесплатный доступ
Статья посвящена изучению палеоэкологических условий обитания сибирячихинских неандертальцев, популяция которых мигрировала на Алтай в период, соответствующий концу MIS4. С учетом того, что она прибыла на уже заселенную денисовцами территорию, важно определить, был ли выбор мест обитания случайным (т.е. мигранты занимали свободные экологические ниши) или обусловленным определенными факторами. На основе изучения фаун мелких млекопитающих с использованием опубликованных результатов палинологического и палеонтологического анализов реконструированы палеоэкологические условия пещер Чагырская и Окладникова в период проживания в них неандертальцев. В процессе исследования проведен анализ фаун мелких млекопитающих по видам, которые в дальнейшем рассматривались в составе биомов (экологических групп). Для определения эпизодов возможного перемещения остатков млекопитающих между отдельными стратиграфическими подразделениями применялись ординационные статистические методы; полученные результаты сопоставлялись с данными микроморфологического и стратиграфического анализов комплексов Чагырской пещеры. Установлено, что поздние неандертальцы на Алтае обитали в похожих палеоэкологических условиях, в которых доминировали степные и лесостепные ландшафты. Выбор карстовых полостей для заселения, возможно, определялся комплексом факторов, среди которых наиболее значимой была доступность промысловых животных и высококачественного каменного сырья. На основе данных статистического анализа состава фауны мелких млекопитающих, а также результатов стратиграфического и микроморфологического анализов сделан вывод о том, что постседиментационные процессы в карстовых полостях могут сопровождаться перемещением остатков млекопитающих, которое оказывает влияние на результаты палеоэкологических реконструкций.
Алтай, палеолит, статистический анализ, мелкие млекопитающие, палеореконструкция
Короткий адрес: https://sciup.org/145146510
IDR: 145146510 | УДК: 569.32:791(571) | DOI: 10.17746/1563-0102.2022.50.1.014-028
Paleoenvironmental conditions of neanderthal habitation in the Altai: Chagyrskaya and Okladnikov caves
We explore the environments of the Sibiryachikha Neanderthals, who had migrated to the Altai at the end of MIS 4. Given that the territory was already populated by Denisovans, the key question is whether the choice of habitat was random (i.e., the immigrants occupied vacant ecological niches) or motivated by other factors. On the basis ofpublished results relating to the study of small-mammal fauna and pollen analysis, the environments of Chagyrskaya and Okladnikov Caves during the Neanderthal habitation are reconstructed. Species of small mammals are viewed as biome members. To reconstruct the episodic transfer of mammalian remains between stratigraphic units, we used ordination statistics and compared the results with those of micromorphological and stratigraphic analyses of Chagyrskaya Cave. It was found that late Neanderthals of the Altai lived in similar environments, dominated by steppe and forest steppe landscapes. The choice of caves for habitation depended on several factors, the key ones being the availability of game and high-quality raw material for manufacturing tools. On the basis of the statistical analysis of small-mammal fauna and the stratigraphic and micromorphological analyses, we conclude that post-sedimentation processes in caves can include vertical transfer of animal remains, affecting environmental reconstructions.
Текст научной статьи Палеоэкологические условия обитания поздних неандертальцев на Алтае: по материалам пещер Чагырская и Окладникова
В последние годы доказано, что в плейстоцене было не сколько волн миграции неандертальцев на Алтай. Последнюю представляют поздние европейские неандертальцы; они появились на этой территории во время, соответствующее концу MIS4, и, вероятно, не контактировали с неандертальцами первой волны. Поздние европейские неандертальцы принесли с собой свою материальную культуру, которая практически не претерпела никаких изменений за время трансконтинентальной миграции [Slon et al., 2018; Kolobova et al., 2020b; Vernot et al., 2021]. Новые данные свидетельствуют о том, что неандертальцы второй волны пришли на территорию, заселенную денисовца-ми, и заняли примерно на 20 тыс. лет определенную экологическую нишу. В этой связи актуальность приобретают вопросы, касающиеся жизнеобеспечивающих стратегий пришлых гоминин на обживаемом пространстве. Особенно важными являются вопросы адаптации поздних европейских неандертальцев к мозаичным ландшафтным и палеоэкологическим условиям Горного Алтая.
Сегодня известны только два пещерных памятника, относящихся к сибирячихинскому технокомплексу, который является самым восточным проявлением ми-кокской культуры, – Чагырская и Окладникова [Деревянко и др., 2013]. На основе информации об условиях обитания неандертальцев в Чагырской пещере уже созданы палеоэкологические реконструкции, такие данные по популяции из пещеры Окладникова скудны.
В настоящей работе предлагаются комплексные палеореконструкции условий обитания заселявших обе пещеры поздних неандертальцев Алтая, выполненные на основе данных по фаунам мелких млекопитающих и опубликованных результатов биологических анализов. Обе пещеры находятся в низкогорье Алтая. Одинаковые высотные пояса характеризуются сходными растительностью и фаунистическими комплексами, что предполагает эксплуатацию поздними неандертальцами одного спектра природных ресурсов. Мелкие млекопитающие, в свою очередь, отражают палеоэкологические условия в непосредственной близости от стоянок, это позволяет проводить прямые корреляции между исследуемыми памятниками.
Новые результаты, с одной стороны, дополняют опубликованные данные по палинологии и крупной фауне Чагырской пещеры [Rudaya et al., 2017], а с другой стороны, обнаруживают определенные противоречия. Так, по данным палинологического анализа, для ландшафтов близ Чагырской пещеры в период накопления слоев 5 и 6 была характерна значительная остепненность, а в период накопления слоя 7 – доминирование тайги. Однако реконструкция по териологическим данным этого не подтверждает. Неоднозначность выводов, сделанных на основе исследования комплексов Чагырской пещеры разными биологическими методами, требует комплексной интерпретации, поэтому нами были привлечены данные детального стратиграфического и микроморфологического анализов рыхлых отложений Чагырской пещеры, показавших комплексность седиментационных и постседиментационных процессов в карстовых полостях, несомненно, оказывающих влияние на результаты применения различных биологических методов.
Материалы и методы
Остеологическая коллекция Чагырской пещеры собрана традиционными методами. После промывания на ситах и просушивания костные остатки извлекались из породы. Материал определялся под бинокулярной лупой МБС-10 с использованием окуляра-микрометра. При проведении исследований привлекались сравнительные коллекции мелких млекопитающих Палеонтологического института им. А.А. Борисяка РАН и Зоологического музея Московского государственного университета им. М.В. Ломоносова. Полученные сведения обрабатывались в программе Excel. При анализе фаунистического состава определялось процентное соотношение по каждому виду в слое от общего количества определимых остатков, по минимальному количеству особей – не высчитывалось, т.к. ранее было показано, что при использовании двух этих методик количественное соотношение между видами в слое сохраняется [Ивлева, 1990]. Для сравнения ископаемых фаун мелких млекопитающих двух пещер были выделены экологические группы: обитатели открытых пространств – степей, лесостепей, полупустынь; обитатели закрытых пространств – лесов; интерзональные, околоводные и горно-степные виды. Рукокрылые в указанном анализе Н.Г. Ивлевой не учитывались. Для корректного сопоставления их не принимали во внимание и при построении палеореконструкций.
Состав фаун мелких млекопитающих из разных стратиграфических подразделений изучался с использованием мультивариантных ординационных методов. В тех случаях, когда анализировалось значительное количество переменных по нескольким выборкам, применялось трехмерное непараметрическое шкалирование (NMDS). При рассмотрении состава фауны по биотопам с целью сопоставления палеоэкологических условий применялся дискриминантный анализ (LDA). Все статистические тесты выполнялись в программе PAST [Hammer, Harper, Ryan, 2001].
Описание стоянок
Чагырская пещера расположена на левом берегу р. Чарыш в отрогах Тигерецкого хребта на северо-западе Алтая (рис. 1, 1). Она северной экспозиции, находится на высоте 353 м над ур. м. и 19 м над уровнем реки. Памятник известен как ключевой объект сиби-рячихинской фации среднего палеолита Горного Алтая, к которой также относится пещера Окладникова [Kolobova et al., 2019а]. Чагырская пещера интерпретируется как базовая стоянка и лагерь неандертальцев по потреблению охотничьей добычи [Kolobova et al., 2019b]. Объектами охоты неандертальцев являлись бизоны и лошади, которые, вероятно, составляли основу белкового рациона обитателей пещеры [Salazar-García et al., 2021]. Материалы, обнаруженные в самой пещере, представляют практически полный цикл утилизации каменного сырья [Шалагина и др., 2020], а также полный цикл изготовления разнообразных костяных орудий [Baumann et al., 2020], среди которых преобладают костяные ретушеры [Kolobova et al., 2020a, p. 89].
Оптикостимулированное датирование (OSL) с измерением единичных зерен полевого шпата позволило установить, что неандертальцы проживали в пещере (слои 5–6д) относительно недолго – от 59 до 49 тыс. л.н. Слой 7, в котором не выявлены следы обитания древнего человека, датируется 314 тыс. л.н. [Kolobova et al., 2020b].
Рыхлые отложения Чагырской пещеры насчитывают несколько стратиграфических единиц. Нумерация слоев ведется сверху вниз по разрезу – от самых молодых к наиболее древним. Описание стратиграфии (рис. 1, 2 ) выполнено доктором наук M. Край-царжем и приводится по опубликованным данным [Kolobova et al., 2019а, 2020b]. Стратиграфическая колонка Чагырской пещеры состоит из восьми слоев, где слой 8 – это красная глина, встречающаяся в углублениях в скальном основании. Слои 1–4 голоценовые, со слоя 5 – плейстоценовые.
Слой 1 – серый и темно-серый некарбонатный суглинистый песок, слегка уплотненный, с большим количеством мелкой речной гальки. Слой представляет собой верхнюю часть слоя 2, измененного в результате поступления органических веществ и вытаптывания человеком.
Слой 2 – желтовато-коричневый суглинистый песок, похожий на слой 3; содержит большое количество галечного материала. Солифлюкция была основным процессом отложения.
Слой 3 – серовато-коричневый суглинистый песок с большим количеством речных галек. Гальки и песок, скорее всего, были перенесены в полость с древних речных террас, расположенных на склоне над пещерой, коллювиальными процессами через карстовые дымоходы в потолке задней камеры. Многочисленные материалы бронзового века и остатки кострищ в этом слое свидетельствуют о культурном характере отложений. В процессе раскопок обнаружены многочисленные норы грызунов, содержащие заполнитель из слоя 3, проникающий в отложения подстилающих слоев (слои 4, 5) и в верхнюю часть слоя 6а. Заполнение нор и рыхлые отложения слоев, в которых они содержались, раскапывались отдельно.
Слой 4 является разновидностью слоя 5a, имеет более сероватый цвет.
Слой 5 – желтоватые карбонатные иловые отложения. Это комплекс пластов, включающий отложе-
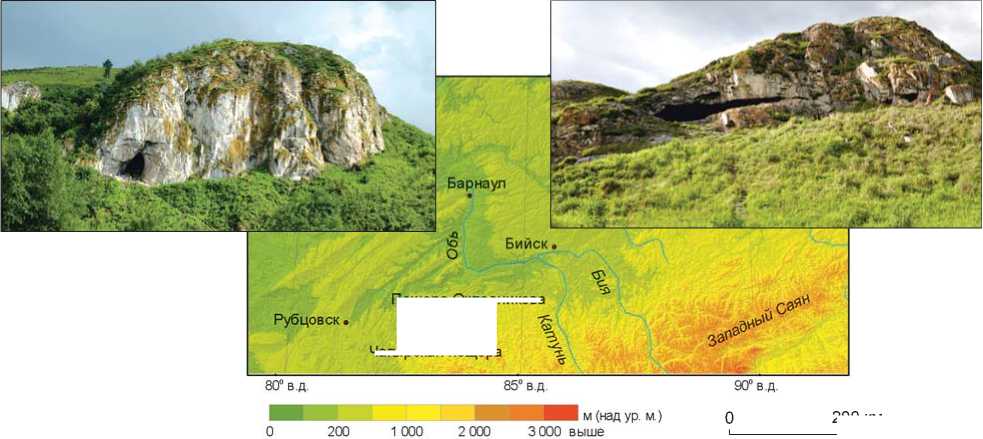
200 км
Рис. 1. Расположение ( 1 ) и стратиграфия пещер Чагыр-ская ( 2 ) и Окладникова ( 3 ).
Пещера Окладникова
Чагырская пещера
ния двух типов: 5a – коллювиальный заполнитель, состоящий из лессовидного ила с редкими округлыми гальками и угловатыми известняковыми обломками, залегает в эрозионных каналах; 5б – угловатые обломки известняка, обычно без рыхлого материала, что указывает на очень быстрое накопление. Осадки типа 5б являются следствием сейсмических событий, которым предшествовало интенсивное механическое выветривание (морозное воздействие).
Слои 6б и 6a – коричневый и оранжевый карбонатный иловый осадок с редкими угловатыми известняковыми обломками, фрагментами костей, каменными артефактами и речной галькой. Слои были определены во время археологических раскопок в 2007–2015 гг. Данные отложения образуют сложную коллювиальную серию, состоящую из более чем двух перемежающихся осадочных единиц. Их следует рассматривать скорее как литологические типы внутри этой серии. Тип 6a более глинистый, оранжевый, похож на слой 7. Тип 6б более алевритовый, немного более плотный, менее пористый, серовато-коричневый и похож на подслой 6c/1. Нижняя граница серии является эрозионной.
Слой 6в (6с) – серый карбонатный алевритовый суглинок с редкими мелкими речными округлыми гальками, многочисленными фрагментами костей, среднепалеолитическими артефактами и немногочисленными обломками известняка. Подслой имеет сложную структуру и может быть расчленен на два подразделения – 6в/1 и 6в/2. Нижнее подразделение 6в/2 представляет собой лессовидный осадок
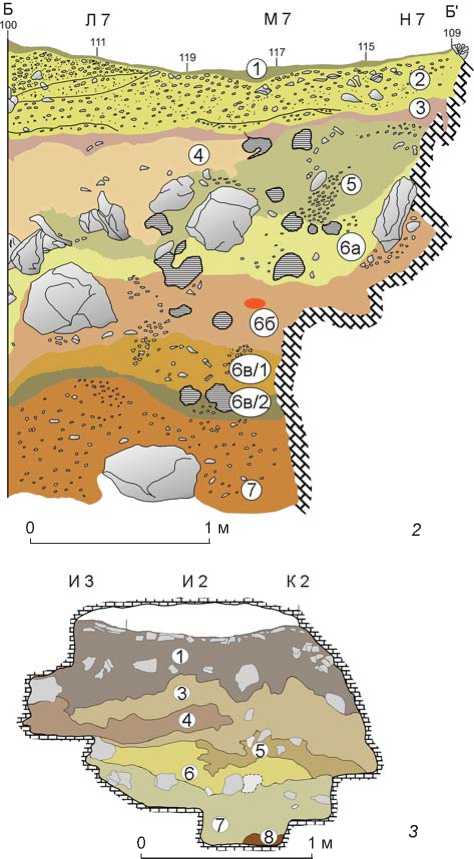
со следами педогенеза. Слой криотурбирован. Крио-турбация произошла после отложения слоя 6a (т.е. значительно позже заселения пещеры неандертальцами). Отложения 6в и нижележащих слоев не были смешаны с отложениями вышележащих слоев. Подслой 6в/2 наиболее насыщен остатками материальной культуры неандетральцев в Чагырской пещере.
Слой 6д (6d) – красновато-коричневый суглинок с мелкими выветрившимися известняковыми обломками, редкими костями и мелкой речной галькой. Содержит отложения из слоя 7, смешанные с материалом слоя 6в (6c) в результате вертикального переотложения под действием мерзлотных процессов.
Слой 7 – красно-коричневый глинистый суглинок с зернами кварца и мелкими, химически выветрившимися обломками известняка и речной гальки. Местами встречаются прослойки зеленоватого ила. Красная глина представляет собой типичный остаточный осадок (тип terra rossa), накопившийся в результате карстового растворения известняка.
Остатки материальной культуры неандертальцев зафиксированы в подошве слоя 5, а также в слоях 6а, 6б, 6в/1 и 6в/2. Слои 6в/1 и 6в/2, согласно данным стратиграфического и микроморфологического анализов, являются непотревоженными. В другие слои археологический материал поступил в результате коллювиального переноса отложений подслоев 6в/1 и 6в/2 из внутренней части пещеры [Kolobova et al., 2020b].
Пещера Окладникова расположена в долине р. Ануй на левом берегу р. Сибирячиха (см. рис 1, 1 ). Она южной экспозиции, находится на высоте 368 м над ур. м., представляет собой карстовую полость с системой разветвленных галерей, гротов и залов, каждый из которых имеет сложную стратиграфию [Деревянко, Маркин, 1992]. Все радиометрические даты, полученные для пещеры Окладникова (включая даты самого молодого слоя 1), находятся в возрастном диапазоне 37–44 тыс. л.н., что соответствует каргинскому интерстадиалу, или, согласно шкале SPECMAP [Imbrie et al., 1984] и уточненной шкале по стеку из низкоширотных океанических скважин ODP 677+MD900963 [Bassinot et al., 1994], этот период относится к MIS3.
Памятник был обнаружен А.П. Деревянко и В.И. Молодиным в 1984 г. и исследовался А.П. Деревянко и С.В. Маркиным в течение четырех лет. Археологический материал происходит из стратиграфических подразделений 1–3, 6, 7.
В пещере выделены девять слоев, но не все прослеживаются в каждой галерее или зале (рис. 1, 3 ). Слой 1 – суглинок с большим количеством кусков известняка и единичными обломками сланцев и песчаников. Слой 2 под навесом включает единичные обломки известняка, в галерее 2 – мелкую щебенку.
Слой 3 – буровато-серый суглинок с разнообразным обломочным материалом в разных частях пещеры. Слой 4 содержит единичные окатанные фрагменты известняка. Слой 5 заключает в себе крупные обломки известняка. Слой 6 – темно-коричневый суглинок, описан только в гроте. Слой 7 – красновато-бурый суглинок с заполнителем в виде выветренных и размягченных сланцев и песчаников. В полостях пещеры в этом слое фиксируются попавшие с поверхности куски известняка, окатанные, что возможно при влиянии водной среды на формирование осадка. Слой 8 состоит из красновато-желтых глин, в галереях – с неокатанными обломками сланцев, песчаников и небольшими обломками кварца. Слой 9 представлен тонкослоистой маломощной супесью на небольшом участке под навесом.
Данные о постдепозиционных процессах в пещере отсутствуют, поскольку в ней не проводились детальные стратиграфические и микроморфологические исследования.
Обе пещеры были заселены в период, соответствующий концу MIS4 – началу MIS3, неандертальцами, генетически близкими к поздним европейским неандертальцам [Veront et al., 2021]. Эта популяция неандертальцев принесла на территорию Алтая ми-кокскую каменную индустрию, для которой характерны радиальное и ортогональное расщепление нуклеусов, а также изготовление плоско-выпуклых бифасов. В орудийных наборах доминируют скребла, в т.ч. простые и конвергентные формы, имеются ретушированные остроконечники и плоско-выпуклые бифасиальные скребла, включающие обушковые ножи типа кайлмессер, тронкированно-фасети-рованные орудия. Неандертальцы из обеих пещер использовали практически одинаковые наборы каменного сырья (ок. 25 типов), в которых наиболее качественными являются яшмоиды засурьинской свиты – из них делали формальные орудия типа плоско-выпуклых бифасов и конвергентные скребла [Деревянко и др., 2015; Шалагина, Кривошапкин, Колобова, 2015].
Материалы проводившихся ранее исследований пещер Чагырская и Окладникова позволяют реконструировать ландшафты в этой части Алтая в период, относящийся к плейстоцену.
Для Чагырской пещеры были проанализированы ископаемые фауны как крупных [Васильев, 2013], так и частично мелких млекопитающих [Деревянко и др., 2013], проведено палинологическое исследование [Rudaya et al., 2017]. На основе полученной информации сделана детальная реконструкция плейстоценовых условий. Во время накопления слоя 5 в условиях аридизации климата преобладали лесные и лесостепные стации с элементами тундры. Судя по сокращению количества лесных видов и увеличе- нию лесостепных и степных, слой 6а является переходным между слоем 5 и 6б, в котором преобладают исключительно степные таксоны. Слой 6в формировался в условиях доминирования открытых биотопов, хотя в начале его накопления отмечена древесная растительность. Все подразделения слоя 6 соответствуют умеренно аридному климату. Аккумуляция слоя 7 связана с распространением перигляциальных ландшафтов. Для этого слоя реконструируется холодный и суровый климат.
Палеореконструкция для пещеры Окладникова опирается также на данные палинологического, териологического и малакологического анализов. При реконструкции не учитывались слои 4 и 5, поскольку в них не обнаружены биологические объекты. Слой 1 формировался в условиях влажного и холодного климата, среди ландшафтов преобладали лесостепи. Для слоя 2 характерен сухой и более теплый климат, чем во время образования слоя 1, были распространены горно-степные и лесостепные стации. Для слоя 3 обычны теплые и сухие условия с повсеместным доминированием степей. Для слоя 6 характерны степи с разнотравно-полынной растительностью, климат был сухим и теплым. Слой 7 накапливался при распространении разнотравно-луговых степей, лесостепей, в условиях влажного и теплого климата.
Результаты
За несколько полевых сезонов (2008, 2009, 2015– 2019, 2021 гг.) был накоплен ко стный материал по мелким позвоночным Чагырской пещеры. Всего идентифицированы более 14 тыс. ко стных элементов, из них более 8 тыс. – определимые до рода и вида. Остеологические остатки, выявленные в 2015–2021 гг., в целом не противоречат полученным ранее данным, они дополняют и подтверждают предположения, сделанные прежде.
Костный материал из Чагырской пещеры разной степени сохранности. Остатки эпохи голоцена белые или кремовые, плейстоцена – светло-желтые, светлокоричневые. Среди плейстоценовых находок встречаются единичные кости и зубы светлой окраски; ранее это было отмечено С.К. Васильевым [2013]. Часть материала имеет следы воздействия желудочного сока, что свидетельствует о том, что она попала в состав тафоценоза из погадок.
Определены до вида 38 мелких млекопитающих, принадлежащих четырем отрядам – Рукокрылые, Насекомоядные, Зайцеобразные и Грызуны. В целом, фауна мелких млекопитающих Чагырской пещеры сходна с ископаемыми фаунами других памятников Северо-Западного Алтая.
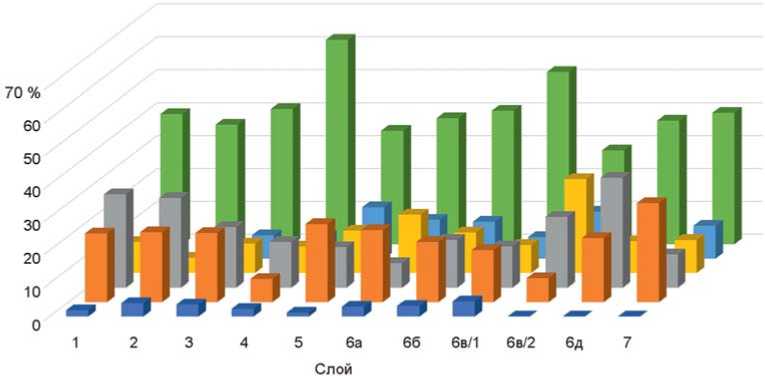
На голоценовые слои приходится половина обработанного материала по мелким млекопитающим. Среди плейстоценовых отложений наибольшее количество остатков содержат слои 5 и 6а. В слоях 6б и 6в их на порядок меньше, что может быть связано с периодичностью заселения пещеры человеком. Именно в слоях 6б и 6в многочисленны каменные орудия и остатки неандертальцев [Vernot et al., 2021]. В тафономии этих слоев проявляет различия фауна крупных млекопитающих [Васильев, 2013; Междисциплинарные исследования…, 2018; Rudaya et al., 2017]. Распределение материалов по экологическим группам показывает, что в фауне Чагырской пещеры преобладают виды, характерные для открытых пространств – степей, лесостепей, лугов, количе ство лесных видов незначительно (рис. 2, 3). Наличие горно-степных видов (скальные полевки и пищухи) свидетельствует о формировании фауны определен-
■1
■2
■3
■5
■6
Рис. 2. Соотношение экологических группировок мелких млекопитающих Чагырской пещеры по слоям.
1 – околоводные; 2 – горно-степные; 3 – лесные; 4 – степные; 5 – полупустынные; 6 – лесостепные.
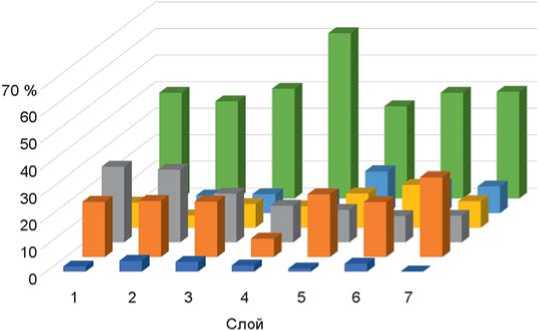
Рис. 3. Соотношение экологических группировок мелких млекопитающих Чагырской пещеры по слоям (слои 6а–6в/2 объединены в один). Усл. обозн. см. на рис. 2.
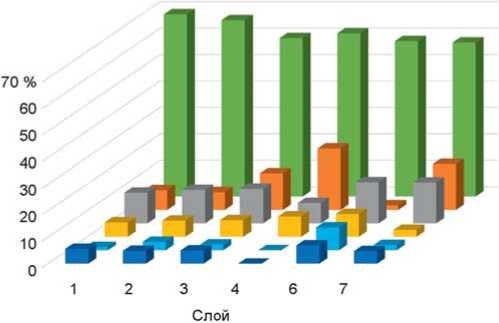
Рис. 4. Соотношение экологических группировок мелких млекопитающих пещеры Окладникова по слоям. Усл. обозн. см. на рис. 2.
ного мозаичного типа, связанного с широтными зонами и вертикальной поясностью.
Анализ ископаемой фауны для пещеры Окладникова был сделан ранее [Ивлева, 1990]. Показано, что влияние открытых пространств нарастает вверх по разрезу – от слоя 7 к слою 1 (рис. 4). На протяжении всей толщи наблюдается мозаичность ландшафта с уменьшением облесненной территории и преобладанием лесостепной зоны [Там же, с. 92]. Индикаторами сухости или увлажнения климата считаются около-водные виды (бобр и водяная полевка), по колебаниям их численности можно судить о климате. Наиболее «сухими» оказались слои 2 и 6 [Там же].
В плейстоценовых слоях Чагырской пещеры обнаружены немногочисленные определимые костные остатки. С учетом того, что слой 6в/1, по данным стратиграфического и микроморфологического анализов, является непотревоженным и при этом содержит фаунистический материал, состав фауны из него был при- нят как реперный для реконструкции палеоэкологических условий и определения следствий постседиментационных процессов.
Сравнивался состав мелких млекопитающих из голоценовых, позднеплейстоценовых и среднеплейстоценовых стратиграфических подразделений пещер Чагырская (табл. 1) и Окладникова (табл. 2). В первую очередь был рассмотрен состав фауны из слоев 6в/1 и 6в/2 пещеры Чагырская, как наименее потревоженных постседиментационными процессами. Кроме того, оценивалась вероятность транспортировки ко стных остатков мелких млекопитающих из одного слоя в другой в результате коллювиальных и зафиксированных мерзлотных процессов.
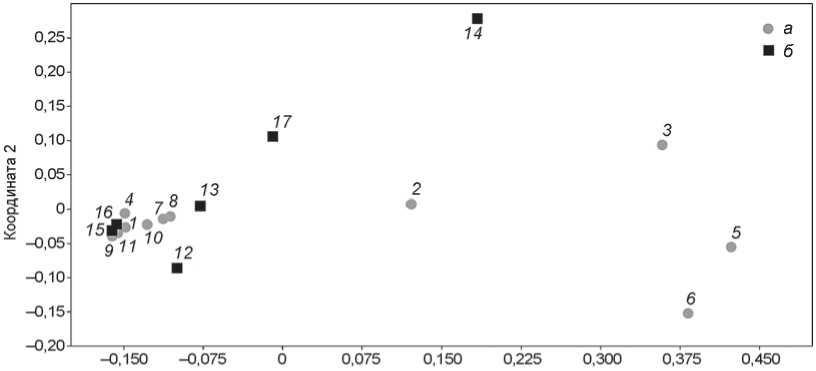
Для проверки предположения о том, что состав мелких млекопитающих отражает палеоэкологические условия, в которых они обитали, использовались статистические орди-национные методы. Поскольку наши данные включают большое количество переменных (31) и небольшое количество выборок (17), было проведено трехмерное непараметрическое шкалирование. Результат шкалирования представлен на диаграмме (рис. 5, уровень стресса 0,02).
Состав мелких млекопитающих из литологических слоев вариабельный. В правой части графика локализируются слои с наибольшим биоразнообразием, в левой – с меньшим биоразнообразием. В последний кластер попали наиболее малочисленные комплексы из современного слоя 1 и слоя 4 Чагырской пещеры. Состав мелких млекопитающих из слоя 7 этой пещеры, соответствующий среднему плейстоцену и тундровым ландшафтам, практически полностью совпадает с таковым вышележащих слоев 6в/2 и 6в/1. Это может быть связано с транспортировкой фаунистических остатков в ходе мерзлотных процессов и плохой сохранностью материала. Во время раскопок в слое 7 выявлены кости только крупных млекопитающих из вышележащего слоя 6в/2.
Состав мелких млекопитающих из слоя 6д, который сформировался в результате мерзлотных процессов из отложений слоев 6в/2 и 7, практически полностью отвечает таковому слоя 6в/2. Это подтверждает выводы, сделанные по составу костного материала из слоя 7: он транспортирован из слоя 6в/2 и представлен остатками плохой сохранности. Скудный со став мелких млекопитающих из непотревоженных слоев 6в/2 и 6в/1 может быть обусловлен особенностями тафономических процессов в этих стратиграфических подразделениях. Тем не менее, выявленное сходство между этими слоями служит
Таблица 1. Костные остатки млекопитающих разных видов в отложениях Чагырской пещеры, экз.
|
Биотопы |
Таксон |
Слой |
||||||||||
|
1 |
2 |
3 |
5 |
6 |
6а |
6б |
6в/1 |
6в/2 |
6д |
7 |
||
|
Полупустынные |
Alactagulus sp. |
– |
– |
– |
1 |
– |
1 |
– |
– |
– |
– |
– |
|
Allactaga sp. |
– |
1 |
2 |
1 |
– |
5 |
– |
– |
– |
– |
– |
|
|
Ellobius talpinus |
– |
– |
2 |
2 |
– |
2 |
– |
– |
– |
– |
1 |
|
|
Lagurus lagurus |
– |
51 |
83 |
184 |
1 |
112 |
3 |
– |
– |
7 |
1 |
|
|
Eolagurus luteus |
– |
7 |
8 |
19 |
– |
24 |
1 |
– |
– |
– |
– |
|
|
Степные |
Lepus sp. |
1 |
4 |
33 |
20 |
– |
15 |
– |
– |
– |
– |
– |
|
Spermophilus sp. |
3 |
25 |
72 |
125 |
– |
190 |
6 |
– |
– |
7 |
2 |
|
|
Marmota sp. |
– |
1 |
2 |
8 |
– |
15 |
– |
– |
– |
– |
– |
|
|
Cricetulus migratorius |
– |
5 |
7 |
20 |
– |
20 |
– |
– |
– |
2 |
– |
|
|
Allocricetulus eversmanni |
1 |
6 |
5 |
4 |
1 |
1 |
– |
– |
– |
– |
– |
|
|
Лесостепные |
Myospalax myospalax |
7 |
98 |
83 |
126 |
3 |
206 |
4 |
4 |
1 |
5 |
1 |
|
Cricetus cricetus |
3 |
51 |
48 |
3 |
– |
4 |
1 |
– |
– |
– |
– |
|
|
Lasiopodomys gregalis |
6 |
103 |
311 |
210 |
– |
242 |
11 |
8 |
– |
22 |
3 |
|
|
Microtus oeconomus |
2 |
45 |
76 |
72 |
– |
25 |
2 |
2 |
– |
7 |
1 |
|
|
Microtus arvalis |
3 |
20 |
19 |
4 |
– |
4 |
– |
– |
– |
1 |
1 |
|
|
Sicista sp. |
– |
7 |
2 |
1 |
– |
– |
– |
– |
– |
– |
– |
|
|
Лесные |
Asioscalops altaica |
– |
13 |
29 |
28 |
1 |
65 |
5 |
– |
– |
10 |
1 |
|
Crocidura sp. |
1 |
7 |
1 |
– |
– |
– |
– |
– |
– |
– |
||
|
Sorex sp. |
2 |
32 |
17 |
10 |
– |
– |
1 |
– |
– |
1 |
1 |
|
|
Sciurus vulgaris |
– |
1 |
– |
4 |
– |
2 |
– |
– |
– |
– |
1 |
|
|
Eutamias sibiricus |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
|
Pteromys volans |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
– |
|
|
Apodemus sp. |
2 |
14 |
7 |
4 |
– |
– |
– |
– |
– |
– |
– |
|
|
Clethrionomys sp. |
9 |
162 |
176 |
116 |
– |
19 |
7 |
1 |
3 |
17 |
– |
|
|
Myopus / Lemmus |
– |
1 |
3 |
5 |
– |
1 |
– |
– |
– |
– |
1 |
|
|
Microtus agrestis |
– |
12 |
15 |
2 |
– |
– |
1 |
– |
– |
3 |
– |
|
|
Околоводные |
Arvicola terrestris |
1 |
36 |
49 |
14 |
– |
36 |
2 |
1 |
– |
– |
1 |
|
Горно-степные |
Alticola sp. |
11 |
177 |
237 |
320 |
– |
267 |
15 |
4 |
– |
15 |
4 |
|
Ochotona sp. |
– |
11 |
47 |
15 |
– |
19 |
2 |
1 |
– |
3 |
– |
|
основанием для предположения об их формировании в одинаковых палеоэкологических условиях и переносе материала из этих слоев в соседние.
Сходство состава мелких млекопитающих из слоя 6б коллювиального генезиса и непотревоженных слоев 6в/2 и 6в/1 свидетельствует о том, что именно из этих слоев поступали остатки мелких млекопитающих.
Совершенно другую картину мы наблюдаем в плейстоценовых слоях 6а (коллювиальный генезис) и 5 (сложный генезис, включающий сейсмические и коллювиальные процессы разной степени интенсивности). Для слоя 5 реконструированы лесостепи, для слоя 6а – сухие аридные степи. По составу фауна мелких млекопитающих этих слоев подобна, но значительно отличается от таковой нижележащих слоев (6в/2, 6в/1, 6б). Слои 5 и 6а содержат остатки, выделяющиеся значительно большим биоразнообразием, что сближает их по составу с голоценовым слоем 3, который содержит материальные о статки афанасьевской культуры бронзового века.
Преобладающая часть остатков мелких млекопитающих, возможно, была перенесена в слой 6а из вышележащих слоев 3 и 5, а не из нижележащих слоев 6в/2, 6в/1, как в случае со слоем 6б. Небольшая часть остатков мелких млекопитающих все-таки могла транспортироваться из нижележащих слоев вместе с остатками материальной культуры поздних неандертальцев.
Таблица 2. Костные остатки млекопитающих разных видов в отложениях пещеры Окладникова, экз. [Ивлева, 1990]
|
Биотопы |
Таксон |
Слой |
|||||
|
1 |
2 |
3 |
4 |
6 |
7 |
||
|
Полупустынные |
Erinacea sp. |
0 |
0 |
1 |
0 |
0 |
0 |
|
Allactaga sp. |
1 |
1 |
3 |
0 |
0 |
2 |
|
|
Ellobius sp. |
5 |
10 |
19 |
0 |
1 |
2 |
|
|
Eolagurus sp. |
3 |
4 |
10 |
0 |
4 |
9 |
|
|
Степные |
Lepus tolai |
1 |
0 |
3 |
0 |
0 |
2 |
|
Spermophilus sp. |
16 |
16 |
43 |
0 |
3 |
9 |
|
|
Marmota sp. |
10 |
4 |
14 |
0 |
1 |
3 |
|
|
Cricetulus sp. |
14 |
9 |
37 |
1 |
1 |
3 |
|
|
Лесостепные |
Myospalax myospalax |
197 |
44 |
136 |
4 |
12 |
31 |
|
Cricetus cricetus |
169 |
32 |
69 |
2 |
3 |
19 |
|
|
Lasiopodomys gregalis |
26 |
109 |
257 |
0 |
9 |
132 |
|
|
Microtus oeconomus |
40 |
44 |
218 |
2 |
7 |
137 |
|
|
Microtus arvalis-agrestis |
85 |
87 |
245 |
0 |
3 |
62 |
|
|
Лесные |
Asioscalops altaica |
12 |
7 |
47 |
0 |
1 |
8 |
|
Sorex sp. |
15 |
10 |
15 |
0 |
3 |
9 |
|
|
Crocidura sp. |
14 |
7 |
22 |
0 |
0 |
4 |
|
|
Pteromys volans |
3 |
2 |
2 |
0 |
0 |
0 |
|
|
Clethrionomys |
24 |
20 |
90 |
1 |
4 |
52 |
|
|
Myopus sp. |
1 |
0 |
3 |
0 |
0 |
0 |
|
|
Microtus agrestis |
18 |
14 |
23 |
0 |
1 |
28 |
|
|
Интерзональные |
Castor fiber |
2 |
3 |
1 |
0 |
0 |
1 |
|
Околоводные |
Arvicola terrestris |
40 |
20 |
76 |
0 |
4 |
30 |
|
Горно-степные |
Alticola sp. |
42 |
25 |
169 |
2 |
0 |
99 |
|
Ochotona sp. |
14 |
7 |
45 |
1 |
1 |
14 |
|
Координата 1
Рис. 5. Непараметрическое трехмерное шкалирование состава мелкой фауны из стратиграфических последовательностей пещер Чагырской ( а ) и Окладникова ( б ).
1 – слой 1; 2 – слой 2; 3 – слой 3; 4 – слой 4; 5 – слой 5; 6 – слой 6а; 7 – слой 6б; 8 – слой 6в/1; 9 – слой 6в/2; 10 – слой 6д;
11 – слой 7; 12 – слой 1; 13 – слой 2; 14 – слой 3; 15 – слой 4; 16 – слой 6; 17 – слой 7.
В пользу данных предположений свидетельствуют высокая степень биоразнообразия состава млекопитающих и сходство состава голоценового слоя 3 с таковым слоев 5 и 6а.
По материалам пещеры Окладникова фиксируется значительное биоразнообразие в плейстоценовых слоях 2, 3, 7. При этом состав мелкой фауны из слоев 4 и 6 практически идентичен составу фауны из слоев 6в/2 и 6в/1 Чагырской пещеры.
По результатам статистического анализа, учитывающего состав фауны мелких млекопитающих из стратиграфических подразделений Чагырской пещеры, была построена схема, включающая также результаты стратиграфического и микроморфологического анализов. Направления транспортировки костных остатков практически полностью соответствуют схемам перемещения рыхлых отложений (рис. 6).
С целью ординации и корреляции палеоэкологи-че ских условий, в которых обитали поздние неандертальцы Алтая, проведен дискриминантный анализ на основе данных по биотопам. Для сокращения переменных остатки мелких млекопитающих в зависимости от их принадлежности к определенным биотопам были суммированы (см. табл. 1, 2). Для проведения анализа выборки разделили на три группы, в которые вошли плейстоценовые и голоценовые слои Чагырской пещеры и литологические слои пещеры Окладникова.
График дискриминант, который отражает соотношение различных среднепалеолитических слоев, принадлежащих различным биотопам, а также взаимное соотношение комплексов слоев отно сительно друг друга, показывает, что накопление плейстоценовых отложений Чагырской пещеры происходило в услови-
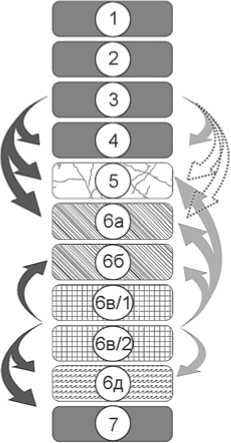
Рис. 6. Схема переноса археологических и остеологических материалов в отложениях Чагырской пещеры.
1 – литологический слой; 2 – литологический слой коллювиального и сейсмического генезиса; 3 – литологический слой коллювиального генезиса; 4 – литологический слой in situ ; 5 – литологический слой мерзлотного генезиса;
6 – перемещение мелкой фауны; 7 – перемещение остатков материальной культуры неандертальцев; 8 – перемещение рыхлых отложений в результате норной деятельности грызунов.
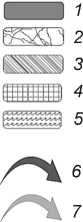

ях доминирования степных – лесостепных ландшафтов, голоценовых – в условиях господства лесостепных – лесных (рис. 7). Рыхлые отложения в пещере Окладникова формировались в условиях преобладания лесостепной и околоводной фауны.
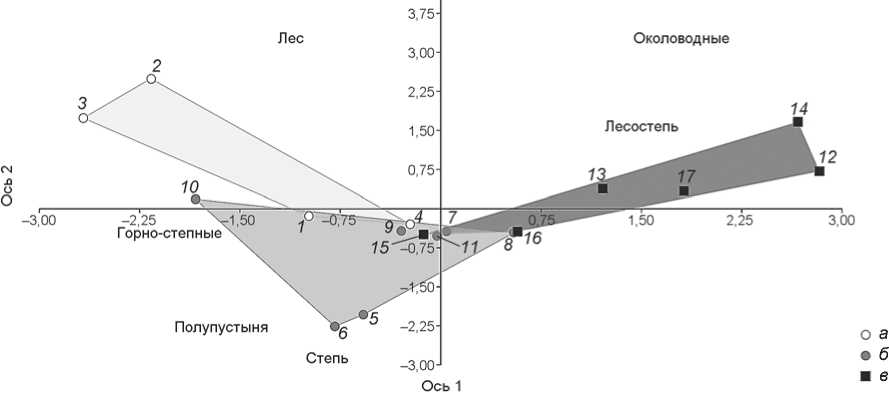
Рис. 7. Дискриминантный анализ состава мелкой фауны по биотопам из голоценовых ( а ) и плейстоценовых ( б ) отложений пещеры Чагырской и плейстоценовых отложений ( в ) пещеры Окладникова.
Дискуссия
Для голоценовых слоев Чагырской пещеры характерна высокая концентрация остатков рукокрылых. По ним определены шесть видов: северный кожанок Eptesicus nilssonii , ночница Брандта Myotis brandti , Myotis dasycneme , прудовая ночница Myotis blitii , сибирский ушан Plecotus ognevi , сибирский трубконос Murina leucogaster . В плейстоценовых слоях количество остатков значительно меньше, чем в голоценовых. В слоях 6б и 6в/1 обнаружено лишь два зуба. Это можно объяснить тем, что кости летучих мышей хрупкие и плохо сохраняются, кроме того, в период заселения пещеры человеком рукокрылые могли не использовать пещеру для дневок или зимовок [Agadjanian, Serdyuk, 2005]. Фауна рукокрылых в настоящей работе не анализируется, ее использование для палеореконструкций затруднительно, поскольку, во-первых, летучие мыши – достаточно мобильная группа мелких млекопитающих, во-вторых, количество их остатков мало. В отложениях пещеры наиболее многочисленны остатки грызунов, представлено более 30 видов; из насекомоядных определено не менее пяти видов. Выявлены также остатки зайцеобразных. Для каждого слоя Чагырской пещеры определен состав мелких млекопитающих.
Слой 1. Характерны виды, встречающиеся на территории современного Алтая; виды-экзоты не обнаружены. Морфология зубов фоновых видов полевок близка к современной. Общий со став фауны этого слоя свидетельствует о распространении лесостепных стаций.
Слой 2. Состав фауны такой же, как в слое 1, но в нем представлены степная пеструшка Lagurus lagurus , желтая пеструшка Eolagurus luteus и лемминг рода Lemmus. Первые два вида населяют степи и полупустыни, третий характерен для тундровых ассоциаций. Все три вида являются маркерами «смешанных» фаун [Вангенгейм, 1977; Громов, 1948; Марков и др., 1965; Черский, 1891]. Хорошо известны голоценовые рефугиумы крупных млекопитающих [Kovacs, 2011; Вартанян, 2004; Stuart et al., 2004; Верещагин, 1988]; мелкие млекопитающие, в отличие от крупных, более устойчивы к изменениям внешней среды [Попова, 2014]. Вероятно, во время накопления слоя 2 недалеко от пещеры какое-то время сохранялся тундростепной биоценоз. В пыльцевом спектре преобладают синантропные виды растений.
Слой 3. Состав фауны такой же, как и в слое 2. На фоне преобладания лесостепных формаций заметны элементы «смешанных» перигляциальных фаун и древесно-кустарничковых биотопов (мышовка Sicista и лесные мыши рода Apodemus), фиксируется тушканчик Allactaga, предпочитающий полупустынные стации. По биоразнообразию слой 3, согласно нашим статистическим выкладкам, близок к слоям 5 и 6а. Возможен частичный перенос материала между слоями. Из крупных млекопитающих отмечен бизон – типичный обитатель открытых ландшафтов на обширных пространствах – от лесостепей до тундро-степей [Междисциплинарные исследования…, 2018]. Наличие остатков этого парнокопытного в районе Ча-гырской пещеры позволяет предполагать, что данная территория в голоцене была рефугиумом мамонтовой фауны. Пыльцевые спектры для этого слоя фиксируют лесостепи с древесными компонентами, что соответствует составу мелких млекопитающих.
Слой 4. Количество материала в нем для голоценового пласта весьма незначительное, преобладают полевки – узкочерепная, обыкновенная, полевка-экономка, остальные виды единичны. Судя по составу мелких млекопитающих, слой может быть захороненной кротовиной, в которую некогда попали погадки хищника [Шалабаев, 2011].
Слой 5. Фауна мелких млекопитающих в нем наиболее обильная для плейстоценовых отложений Чагырской пещеры. Она свидетельствует о присутствии степных и полупустынных биотопов в период формирования слоя. Имеются остатки тушканчика Allactaga и тарбаганчика Alactagulus, предпочитающего такыры с плотным глинистым грунтом. Для этих полупустынь характерны также степная пеструшка и желтая пеструшка Eolagurus luteus. Слой 5 формировался, видимо, в условиях перигляциальной холодной степи; на это указывают о статки лемминга Lemmus sibiricus. Палинологический анализ фиксирует в этом слое таежный биом. Типично таежные виды мелких млекопитающих представлены несколькими зубами и ко стями белки; материалы других типичных обитателей лесов не обнаружены. Остатки белки могут быть связаны с небольшими лесами по берегам рек, не игравшими значительной роли [Междисциплинарные исследования…, 2018]. О наличии древесно-кустарничковой растительности свидетельствуют остатки лесных мышей. Среди крупных млекопитающих преобладали степные и лесостепные виды. Как показано ранее, в формировании тафоценоза крупных млекопитающих принимали участие пещерные гиены [Kolobova et al., 2019b]; по праву считающиеся уникальными коллекторами, они подбирают на своей охотничьей территории остатки практически полного спектра фауны средних и крупных млекопитающих. Скорее всего, в данное время человек посещал пещеру редко, следовательно, она была также привлекательна для хищников, собирающих мелкую фауну, например, хищных птиц или мелких куньих. В контексте нашего исследования немаловажно, что крупные хищники, устраивая в пещере лежки, вероятно, вскрывали рыхлые отложения и таким образом обеспечивали некоторое пе- ремещение материала, который мог частично попадать в вышележащие или подстилающие слои.
Слой 6а. В нем, как и в предыдущем слое, многочисленны костные остатки. Для слоя характерно большее, чем для слоя 5, количество пустынных и полупустынных видов мелких млекопитающих. Отмечен сибирский лемминг. Слои 5 и 6а формировались в разных условиях. В фауне крупных млекопитающих количество лесостепных видов в слое 6а меньше, чем в слое 5. Данные палинологии предполагают переходный характер этого слоя от «таежного» (слой 5) к «степному» (слой 6б). Фауна мелких млекопитающих не отражает явной «переходности». Поскольку в слое 5 не было явной таежной фауны, то в слое 6а не наблюдается сокращение количества лесных видов и нарастание количества степных. В целом, период формирования слоев 6 совпадает со временем присутствия человека в пещере. Это объясняет малочисленность остатков мелких млекопитающих: дикие животные и птицы избегали близкого соседства с человеком.
Слой 6б. Количество материала по мелким млекопитающим невелико. В слое не представлены тушканчики и древесно-кустарничковые обитатели. В фауне обычны представители луговых и лугово-степных биотопов (суслики, цокор, полевки обыкновенная, экономка), количество степных видов незначительно. Фауна крупных млекопитающих фиксирует преобладание степных видов. В пыльцевых спектрах наблюдается увеличение количества пыльцы злаков. Несоответствие этому данных по мелким млекопитающим можно объяснить малым количеством материала или другими тафономическими особенностями.
Слои 6в/1 и 6в/2. Количество остатков мелких млекопитающих тоже мало. Видовой состав практически не отличается от такового слоя 6б. Согласно результатам статистического и микроморфологического анализов, слой 6в является источником формирования тафоценоза слоя 6б. Частичный перенос материала из слоя возможен, если учитывать хозяйственную деятельность человека в пещере в то время. В фауне преобладают остатки представителей лугово-степных стаций, единичны остеологические материалы обитателей каменистых россыпей. Среди крупных млекопитающих доминируют степные виды. Данные палинологии фиксируют сокращение количества злаковых.
Слой 6д. Среди мелких млекопитающих этого слоя доминируют серые полевки. Состав микрофауны слоя, практически полностью соответствующий составу слоя 6в/2, подтверждает выводы геологов о формировании слоя в результате действия мерзлотных процессов.
Слой 7. Количество материала для биологического анализа из этого слоя незначительно. Среди мелких млекопитающих фиксируются элементы «смешанной» перигляциальной фауны, отмечена слепушонка, избегающая ковыльных и полынно-злаковых степей. Представлены обитатели горностепных и луговых биотопов. Климат, вероятно, был аридный. Среди крупных млекопитающих преобладают степные виды. Палинологические сборы регистрируют таежный, степной и тундровый биомы; эти данные идут вразрез с результатами палеотериологии. Такое возможно при попадании в этот слой отложений перекрывающих слоев, что не противоречит результатам статистического анализа.
Данные микротериологического анализа находок из пещеры Окладникова указывают на преобладание лесостепных биотопов в районе пещеры и потепление климата ближе к голоцену. В пещере обнаружены также остатки представителей «смешанных» перигляциальных ландшафтов. Обнаружены четыре зуба лесного лемминга рода Myopus [Ивлева, 1990]. Эти лемминги обитали в разнообразных лесах – еловых, пихтовых, кедровых, смешанных, т.е. биотопическая приуроченность у них не такая, как у тундрового лемминга. Согласно последним данным, лесные лемминги отличаются от последних по третьему верхнему моляру и морфометрическим характеристикам других зубов [Пономарев и др., 2011]. Вероятно, необходима ревизия материалов по леммингам из пещеры Окладникова, чтобы подтвердить или опровергнуть наличие тундровых компонентов в районе этой пещеры в плейстоцене.
Проведенный дискриминантный анализ подтвердил результаты как микротериологического анализа, так и многомерного шкалирования, а также выявил статистически значимое сходство в составе фауны мелких млекопитающих (Чагырская пещера – слои 6в/1, 6в/2, 6б, пещера Окладникова – слой 6). Согласно результатам дискриминантного анализа, поздние неандертальцы Алтая, следы пребывания которых найдены в обеих пещерах, обитали преимущественно в лесостепных ландшафтах. Значительное присутствие околоводных видов в отложениях пещеры Окладникова может быть связано с меньшим расстоянием до реки, по сравнению с расстоянием до реки от пещеры Чагырская.
В целом, данные по мелким млекопитающим двух пещер фиксируют сходные ландшафты и условия в каргинский период. Согласно летописи микротериологии, почти весь период осадконакопления в низ-когорье Алтая преобладали лесостепи.
Выводы
Исследование показало, что на Алтае в период, соответствующий финалу MIS4 – началу MIS3, поздние неандертальцы обитали в сходных палеоэкологических условиях, в которых доминировали степные – ле- состепные ландшафты. При этом большая доля около-водных видов в фаунистических комплексах пещеры Окладникова, вероятно, свидетельствует о различиях в гидрологических режимах рек Сибирячиха и Чарыш в плейстоцене. Возможно, Чарыш тогда не был столь полноводен, как сейчас.
Несмотря на незначительность выборки из двух объектов, можно предполагать, что выбор поздними алтайскими неандертальцами – обитателями двух пещер, находящихся в сходных палеоэкологических условиях, вряд ли был случайным. Неандертальцы из микокских популяций Центральной и Восточной Европы практиковали охоту на крупных млекопитающих широкого спектра, в который входили практиче ски все представители мамонтовой фауны [Richter, 2006]. В комплексах Кавказа и Крыма, наиболее близких географически, неандертальцы охотились преимущественно на крупных травоядных (бизоны, плейстоценовые ослы и др.) [Uthmeier, Chabai, 2010; Голованова и др., 2018; Ramírez-Pedraza et al., 2020]. Вероятно, сибирячи-хинские неандертальцы, которые также практиковали охоту на бизонов и лошадей, занимали наиболее пригодные для этой деятельно сти экологические ниши – пещеры в руслах рек, служивших транзитными коридорами для сезонных миграций крупных травоядных. Подобная зависимость адаптационных стратегий древних гоминин от среды обитания и миграций плейстоценовых животных уже фиксировалась в нескольких районах Центральной и Северной Азии [Agadjanian, Shunkov, 2018; Хацено-вич и др., 2021; Zavala et al., 2021].
Дополнительной причиной, обусловившей выбор именно этих карстовых полостей для заселения, вероятно, была возможность добычи яшмоидов за-сурьинской свиты и халцедонолитов – высококачественного каменного сырья, которое использовалось для изготовления орудий, типичных для сибирячихин-ских индустрий (плоско-выпуклых бифасов и конвергентных скребел). Пещеры Чагырская и Окладникова – единственные среднепалеолитические памятники в регионе, расположенные на источниках этих горных пород [Деревянко и др., 2015; Kolobova et al., 2019b]. В настоящее время только для верхнепалеолитических памятников Северной и Центральной Азии отмечена зависимость выбора мест расселения человеческих коллективов от наличия высококачественного каменного сырья [Rybin et al., 2020].
Статистиче ское изучение состава фауны мелких позвоночных доказало свою полезность для сравнения комплексов, содержащих десятки видов, а также позволило сделать выводы о постседиментационных процессах, которые не только не противоречат дан- ным стратиграфии и микроморфологии, но и дополняют их. Было определено, что на состав фауны отдельных слоев Чагырской пещеры оказывает влияние собственная сохранность органики. Так, в слое 7, седиментация которого, по палинологическим данным, происходила в условиях тайги/тундры, были зафиксированы мелкие млекопитающие из вышележащих стратиграфических подразделений (слой 6в/2), по составу отвечающие степным – лесостепным ландшафтам. Состав фауны мелких млекопитающих из слоев коллювиального генезиса (6б) в значительной степени совпадает с таковым слоя-источника переотложения, что подтверждает предположение о транспортировке не только остатков материальной культуры неандертальцев, но и фауны. На состав фаун оказывают влияние и различные нарушения слоев, прежде всего норы грызунов из вышележащих слоев, несмотря на то, что раскопки проводились по современной методике. Так, в слое 6а было зафиксировано бóльшее влияние вышележащих слоев (3 и 5), чем непотревоженного стратиграфического подразделения, содержащего многочисленный археологический материал и крупные кости животных.
Данные по фауне мелких млекопитающих в сочетании с результатами стратиграфических, микро-морфологических и палинологических исследований могут использоваться при реконструкции постседиментационных процессов в карстовых памятниках. В Чагырской пещере зафиксированные перемещения остатков мелких млекопитающих между слоями практически полностью совпадают с постседиментационными процессами.
Обычно палеореконструкции по фауне мелких млекопитающих и по палинологии проводятся в отрыве от детальных стратиграфических реконструкций, учитывающих эпизоды перемещения или переотложения стратиграфических подразделений. Комплексы каждого слоя рассматриваются как дискретные единицы, отражающие последовательное развитие палеоландшафтов снизу вверх по разрезам, что может не полностью соответствовать процессам седиментации. Исследование комплексов Чагырской пещеры выявило сложность стратиграфических процессов в карстовых полостях, их влияние на состав фауны мелких млекопитающих и в итоге – на палеореконструкции.
Анализ фауны мелких млекопитающих выполнен при финансовой поддержке РФФИ, проект № 18-09-40070, статистические исследования выполнены за счет РНФ, проект № 21-18-00376.
Список литературы Палеоэкологические условия обитания поздних неандертальцев на Алтае: по материалам пещер Чагырская и Окладникова
- Вангенгейм Э.А. Палеонтологическое обоснование стратиграфии антропогена Северной Азии. – М.: Наука, 1977. – 172 с.
- Вартанян С.Л. Палеогеография позднего неоплейсто-цена и голоцена территории острова Врангеля: дис. … канд. геогр. наук. – СПб., 2004. – 16 с.
- Васильев С.К. Фауна крупных млекопитающих из плейстоценовых отложений Чагырской пещеры (Северо-Западный Алтай) по материалам раскопок 2007–2011 годов // Археология, этнография и антропология Евразии. – 2013. – № 1. – С. 28–44.
- Верещагин Н.К. Палеогеография и палеоэкология зверей мамонтовой фауны // Общая и региональная териогеография. – М.: Наука, 1988. – C. 19–32.
- Голованова Л.В., Дороничев В.Б., Александровский А.А. Пещеры Лагонакского нагорья: стратиграфия и палеоэкология археологических памятников каменного века // Вопросы географии. – 2018. – Вып. 147: Спелеология и карстоведение. – С. 268–298.
- Громов В.И. Палеонтологическое и археологическое обоснование стратиграфии континентальных отложений четвертичного периода на территории СССР. – М.: Изд-во АН СССР, 1948. – 521 с.
- Деревянко А.П., Маркин С.В. Мустье Горного Алтая (по материалам пещеры им. Окладникова). – Новосибирск:Наука, 1992. – 288 с.
- Деревянко А.П., Маркин С.В., Зыкин В.С., Зыкина В.С., Зажигин В.С., Сизикова А.О., Солотчина Э.П., Смолянинова Л.Г., Антипов А.С. Чагырская пещера – стоянка среднего палеолита Алтая // Археология, этнография и антропология Евразии. – 2013. – № 1. – С. 2–27.
- Деревянко А.П., Маркин С.В., Кулик Н.А., Колобова К.А. Эксплуатация каменного сырья представителями сибирячихинского варианта среднего палеолита Алтая // Археология, этнография и антропология Евразии. – 2015. – Т. 43, № 3. – С. 3–16.
- Ивлева Н.Г. Микротериологические материалы из пещер им. Окладникова и Денисова на Алтае // Комплексные исследования палеолитических объектов бассейна р. Ануй. – Новосибирск: ИИФФ СО РАН, 1990. – С. 82–104.
- Марков К.К., Лазуков Г.И., Николаев В.А., Величко А.А. Четвертичный период: (ледниковый период – антропогеновый период). – М.: Изд-во Моск. гос. ун-та: Недра, 1965. – Т. I. – 371 с.
- Междисциплинарные исследования Чагырской пещеры – стоянки среднего палеолита Алтая / Деревянко А.П., Маркин С.В., Колобова К.А., Чабай В.П., Рудая Н.А., Виола Б., Бужилова А.П., Медникова М.Б., Васильев С.К., Зыкин В.С., Зыкина В.С., Зажигин В.С., Вольвах А.О., Робертс Р.Г., Якобс З., Бо Ли. – Новосибирск: Изд-во ИАЭТ СО РАН, 2018. – 468 с.
- Пономарев Д.В., Исайчев К.И., Бикбаева Г.В., Кряжева И.В. Материалы к родовой диагностике леммингов Lemmus and Myopus // Вестн. Института геологии Коми Науч. центра УрО. – 2011. – № 11. – С. 10–11.
- Попова Л.В. Судьбы мамонтовой фауны и феномен Черной королевы // Вісник Національного науково-природничого музею. – 2014. – № 12. – С. 20–32.
- Хаценович А.М., Вишневская И.А., Базаргур Д., Вольвах А.О., Клементьев А.М., Ге Дж., Жилич С.В., Марченко Д.В., Окунева Т.Г., Когай С.А., Гунчинсурэн Б., Рыбин Е.П., Олсен Дж.У., Деревянко А.П. Природная среда и эпизоды заселения Центральной Монголии в позднем плейстоцене: по материалам памятников в долине реки Орхон // Археология, этнография и антропология Евразии. – 2021. – Т. 49, № 2. – С. 3–22.
- Черский И.Д. Описание коллекции послетретичных млекопитающих животных, собранных Ново-Сибирской экспедицией 1885–1886 гг. – СПб.: Имп. акад. наук, 1891. – 706 с.
- Шалабаев Р.Н. Избирательность хищных птиц в питании Micromammalia на остепненной территории // Современные проблемы науки и образования. – 2011. – № 4. – С. 51–56.
- Шалагина А.В., Кривошапкин А.И., Колобова К.А. Тронкированно-фасетированные изделия в палеолите Северной Азии // Археология, этнография и антропология Евразии. – 2015. – Т. 43, № 4. – С. 33–45.
- Шалагина А.В., Харевич В.М., Мори С., Боманн М., Кривошапкин А.И., Колобова К.А. Реконструкция технологических цепочек производства бифасиальных орудий в индустрии Чагырской пещеры // Сиб. истор. исследования. – 2020. – № 3. – С. 130–151.
- Agadjanian A.K., Serdyuk N.V. History of Mammalian Communities and Paleogeography of Altai Mountains in the Paleolithic // Paleontol. J. – 2005. – Vol. 39, N 6. – P. 645–820.
- Agadjanian A.K., Shunkov M.V. Late Pleistocene Mammals of the Northwestern Altai : Report 1. Anui Basin // Paleontol. J. – 2018. – Vol. 52. – P. 1450–1460.
- Bassinot F.C., Labeyrie L.D., Vincent E., Quidelleur X., Shackleton N.J., Lancelot Y. The astronomical theory of climate and the age of the Brunhes-Matuyama magnetic reversal // Earth and Planetary Science Letters. – 1994. – Vol. 126. – P. 91–108.
- Baumann M., Plisson H., Rendu W., Maury S., Kolobova K., Krivoshapkin A. Neandertalian bone industry at Chagyrskaya cave (Altai, Russia) // Quat. Intern. – 2020. – Vol. 559. – P. 68–88.
- Imbrie J., Hays J.D., Martinson D.G., McIntyre A., Mix A.C., Morley J.J., Pisias N.G., Prell W.L., Shackleton N.J. The Orbital Theory of Pleistocene Climate: Support from a Revised Chronology of the Marine δ18O Record // Milankovitch and Climate. – N. Y.: Springer, 1984. – Pt. 1. – P. 269–305.
- Hammer Ø., Harper D.A.T., Ryan P.D. PAST: paleontological statistics software package for education and data analysis // Palaeontologia Electronica. – 2001. – Vol. 4 (1). – P. 4.
- Kolobova K.A., Chabai V.P., Shalagina A.V., Krajcarz M.T., Krajcarz M., Rendu W., Vasiliev S.K., Markin S.V., Krivoshapkin A.I. Exploitation of the natural environment by Neanderthals from Chagyrskaya Cave (Altai) // Quartär. – 2019a. – Vol. 66. – P. 7–31.
- Kolobova K.A., Shalagina A.V., Chabai V.P., Markin S.V., Krivoshapkin A.I. Signifi cation des technologies bifaciales au Paléolithique moyen des montagnes de l’Altaï // L’Anthropologie. – 2019b. – Vol. 123, iss. 2. – P. 276–288.
- Kolobova K., Rendu W., Shalagina A., Chistyakov P., Kovalev V., Baumann M., Koliasnikova A., Krivoshapkin A. The application of geometric-morphometric shape analysis to Middle Paleolithic bone retouchers from the Altai Mountains, Russia // Quat. Intern. – 2020a. – Vol. 559. – P. 89–96.
- Kolobova K., Roberts R., Chabai V., Jacobs Z., Krajcarz M., Shalagina A., Krivoshapkin A., Li B., Uthmeier T., Markin S., Morley M., O’Gorman K., Rudaya N., Talamo S., Viola B., Derevianko A. Archaeological Evidence for Two Separate Dispersals of Neanderthals into Southern Siberia // Proceedings of the National Academy of Sciences. – 2020b. – Vol. 117 (6). – P. 2879–2885.
- Kovacs J. Radiocarbon chronology of Late Pleistocene large mammal faunas from the Pannonian basin (Hungary) // Bull. of Geosciences. – 2011. – Vol. 87. – P. 13–19.
- Ramírez-Pedraza I., Rivals F., Uthmeier T., Chabai V.P. Palaeoenvironmental and seasonal context of the Late Middle and Early Upper Palaeolithic occupations in Crimea: an approach using dental wear patterns in ungulates // Archaeological and Anthropological Sciences. – 2020. – Iss. 268. – doi:https://doi.org/10.1007/s12520-020-01217-9.
- Richter J. Neanderthals in their landscape // Neanderthals in Europe. – Liège; Tongeren: ERAUL 117 & ATVAT VCA, 2006. – P. 17–32.
- Rybin E.P., Paine C.N., Khatsenovich A.M., Bolorbat Ts., Talamo S., Marchenko D.V., Rendu W., Klementiev A.M., Odsuren D., Gunchinsuren B., Zwyns N. A new Upper Paleolithic occupation at the site of Tolbor-21 (Mongolia): site formation, human behavior and implications for the regional sequence // Quat. Intern. – 2020. – Vol. 559. – P. 133–149.
- Rudaya N., Vasiliev S., Viola B., Talamo S., Markin S. Palaeoenvironments during the period of the Neanderthals settlement in Chagyrskaya cave (Altai Mountains, Russia) // Palaeogeography, Palaeoclimatology, Palaeoecology. – 2017. – Vol. 467. – P. 265–276.
- Salazar-García D.C., Power R.C., Rudaya N., Kolobova K., Markin S., Krivoshapkin A., Henry A.G., Viola B. Dietary evidence from Central Asian Neanderthals: A combined isotope and plant microremains approach at Chagyrskaya Cave (Altai, Russia) // J. of Hum. Evol. – 2021. – Vol. 156. – doi: https://doi.org/10.1016/j.jhevol.2021.102985.
- Stuart A.J., Kosintsev P.A., Higham T.F. G., Lister A.M. Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth // Nature. – 2004. – Iss. 431. – P. 684–689.
- Slon V., Mafessoni F., Vernot B., Filippo C. de, Grote S., Viola B., Hajdinjak M., Peyrégne S., Nagel S., Brown S., Douka K., Higham T., Kozlikin M.B., Shunkov M.V., Derevianko A.P., Kelso J., Meyer M., Prüfer K., Pääbo S. The genome of the offspring of a Neanderthal mother and a Denisovan father // Nature. – 2018. – Iss. 561. – P. 113–116.
- Uthmeier Th., Chabai V.P. Neanderthal subsistence tactics in theCrimean Micoquian // Settlement dynamics of the Middle Paleolithic and Middle Stone Age. – Tübingen: Kerns Verlag, 2010. – Vol. III. – P. 195–234.
- Vernot B., Zavala E.I., Gómez-Olivencia A., Jacobs Z., Slon V., Mafessoni F., Romagné F., Pearson A., Petr M., Sala N., Pablos A., Aranburu A., Castro J.M.B., Carbonell E., de, Li B., Krajcarz M.T., Krivoshapkin A.I., Kolobova K.A., Kozlikin M.B., Shunkov M.V, Derevianko A.P., Viola B., Grote S., Essel E., Herráez D.L., Nagel S., Nickel B., Richter J., Schmidt A., Peter B., Kelso J., Roberts R.G., Arsuaga J.-L., Meyer M. Unearthing Neanderthal population history using nuclear and mitochondrial DNA from cave sediments // Sci. – 2021. – Vol. 372. – Iss. 6542.
