Patterns in the population history of Northern Eurasia from the Mesolithic to the Early Bronze Age, based on craniometry and genetics

Author: Kozintsev A.G.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 4 т.49, 2021.
Free access
Short address: https://sciup.org/145146404
IDR: 145146404 | DOI: 10.17746/1563-0110.2021.49.4.140-151
Text of the article Patterns in the population history of Northern Eurasia from the Mesolithic to the Early Bronze Age, based on craniometry and genetics
The recent achievements of paleogenetics, especially at the whole-genome level, are increasingly helpful in resolving matters over which specialists in skeletal studies have been arguing for many decades. One of the most illustrative examples is the debate as to whether the cranial specificity of the Neolithic and Early Bronze Age Baikalians was caused by European admixture (Debetz, 1948: 61) or by the preservation of a specific trait combination known as Paleosiberian (Debetz, 1951: 95). Craniologists have failed to reach a consensus on that matter: some believed European admixture to have been minimal (Mamonova, 1983), whereas other claimed that its role was critical and that the term “Paleosiberian” should be abolished (Alekseyev, Gokhman, Tumen, 1987).
Genetic studies have given a quietus to this argument by having convincingly demonstrated that the specificity of Baikalians was not due to European admixture—at least not to the gene flow that, according to Debetz, had reached the Baikal area from the west along the steppes beginning from the Early Bronze Age (Damgaard et al., 2018). This sort of admixture has been detected in just one of 53 Neolithic and Early Bronze Age individuals from the Cis-Baikal and Trans-Baikal regions (1.9 %)—apparently, in a male buried at a Glazkovo cemetery Khaptsagay, on the Upper Lena (Yu et al., 2020)*. Other individuals from Late Neolithic and Early Bronze Age burials in the Cis-Baikal deviate from those with the highest amount of the autosomal component NEA (Northeast Asian), especially marked in Mongoloids, such as those from the Neolithic Devil’s Gate in Primorye, and in Kitoi people**, not toward Bronze Age people of the western steppe, but toward descendants of the Upper Paleolithic Siberians, marked by the ANE (Ancient North Eurasian) autosomal component, first described in a boy buried at Malta some 24 ka BP, as well as in a male and a girl who lived at Afontova Gora 15–17 ka BP (Raghavan et al., 2014; Fu et al., 2016). In the Cis-Baikal, the amount of ANE increases from 14 % during the Kitoi stage to 23 % during later stages of the Neolithic and Early Bronze Age (Yu et al., 2020).
Abandoning the term “Paleosiberian”, therefore, is hardly reasonable, because it likely refers to the morphological correlate of the ANE component. This, of course, does not imply direct connection between cranial features and the analytically separated part of the genome ensuring the most efficient classification of groups. An indirect connection, however, is quite probable, because cranial trait combinations, too, are a classification tool, designed for the same purpose.
The origin of the ANE component is a complete mystery. While archaeological parallels to the Malta-Buret culture in Eastern and Central Europe are well known, the hypothesis that this culture was introduced to Siberia by migrants from those regions is not supported by genetic data (Fu et al., 2016). No more plausible is the opposite hypothesis: although the ANE component was introduced to Central and Western Europe from the east, this happened much later than the Upper Paleolithic, since ANE was absent there before the Yamnaya expansion (Ibid.; Haak et al., 2015).
ANE makes up the principal share of the EHG (Eastern Hunter-Gatherer) autosomal component, whose content is especially high in the genomes of Mesolithic and Early Neolithic inhabitants of northeastern Europe buried at Yuzhny Oleny Ostrov, Popovo, Sidelkino, Lebyazhinka IV, etc. (Haak et al., 2015; Damgaard et al., 2018). They passed EHG on to the Yamnaya people, from whom it was inherited by several filial populations, including Afanasyevans. As early as the Mesolithic, EHG was introduced from northern Russia to Scandinavia, as evidenced by genomes of the Motala people in southern Sweden. Their ancestors had migrated there from the east along the coast of Norway, because the share of EHG in more southern populations, such as the earlier Kunda people of the eastern Baltic, is lower (Haak et al., 2015; Mittnik et al., 2018). As shown by the chronologically diverse materials from the Zvejnieki cemetery in Latvia, the proportion of EHG rises in the Middle Neolithic in parallel with archaeological signs of contact with the PitComb Ware culture (Jones et al., 2017).
The roots of ANE in Siberia are much deeper than those of EHG in Eastern Europe, where, judging by the mitochondrial haplogroups, ANE was introduced from Siberia (Mittnik, 2018). The ancestor of ANE was the ANS (Ancient North Siberian) autosomal component, represented in a male from the Upper Paleolithic Yana site, dating to 31.6 ka BP (Sikora et al., 2019). ANS is thought to have originated among West Eurasians soon after their divergence from East Eurasians about 43 ka BP. The picture is complicated by an approximately 22 % genetic contribution received by early West Eurasians from East Asians shortly after their split (Ibid.).
From Siberia, ANE spread in both directions— westwards and eastwards. The ancestors of American Indians brought it to the New World, where its share in modern aboriginal populations is about 30–40 % (Flegontov et al., 2016; Sikora et al., 2019). Modern Old World groups closest to native Americans in this respect are Chukchi, Koryaks, Kets, and Selkups, and among the ancient groups, Okunev people and those of the Botai culture*, suggesting that both these groups are direct descendants of the Malta-Buret people. In both these populations, ANE is mixed with the Northeast Asian component. The same mixture is seen in two males who are genetically very close to native Americans—that from the Upper Paleolithic site Ust-Kyakhta in the western Trans-Baikal, dating to ~14 ka BP (Yu et al., 2020), and that from the Mesolithic site Duvanny Yar on the Kolyma, dating to ~10 ka BP (Sikora et al., 2019).
It is absolutely unclear from where the Upper Paleolithic inhabitants of western Caucasus (Satsurblia, 13 ka BP) and the Early Neolithic people of Iran (Ganj Dareh, 8th millennium BC) received large amounts of the ANE
*Okunev people have an additional western admixture originating from the Yamnaya-Afanasyevo population (Damgaard et al., 2018; Jeong et al., 2019; Yu et al., 2020).
component (Lazaridis et al., 2018). Judging by the huge distribution area of ANE, it was a legacy of early Homo sapiens , members of the Boreal meta-population (Biasutti, 1941: 275; Kozintsev, 2013, 2014), who inhabited northern Eurasia and America at various stages of the disintegration of this major branch of the human species and were variously affected by contacts between its filial branches.
Genomic studies have revealed a number of early east-west clines passing across northern Eurasia and connecting northeastern Europe (the distribution area of EHG) with Trans-Baikal and the Amur Basin, where the NEA/AEA component was distributed (Damgaard et al., 2018; Jeong et al., 2019; Narasimhan et al., 2019; Kılınç et al., 2021). They might be regarded as genetic correlates of the Caucasoid to Mongoloid vector of traditional classifications, were it not for the fact that the earliest individual carrying ANE—the key component in these clines—was geographically neither western nor eastern, but intermediate. And whereas the considerable (Upper Paleolithic) age of the NEA/AEA component in the eastern part of the gradients is beyond doubt, like its role in the early population history of northern Eurasia (Mao et al., 2021; Yu et al., 2020), the western component, differing from EHG by a large amount of southern admixtures (such as Iranian, Caucasian, and Anatolian) had taken part in this process only since the Early Bronze Age—specifically, since the time of the Yamnaya-Afanasyevo migrations. Before that, the role of the equal partner of the NEA/AEA component was played by ANE.
This means that the challenge for skeletal studies in reconstructing population history is to focus on trait combinations that might be evolutionarily conservative rather than hybrid, such as Paleosiberian and the like. A well-founded hypothesis about one of them, marking the so-called Southern Eurasian Formation, was advanced, postulating its autochthonous nature in the Altai-Sayan region and the adjoining mountain systems (Chikisheva, 2012: 57, 153, 169). The standard example of this trait combination is presented by the Okunev people. Another combination, observed in a number of Neolithic and Early Bronze Age samples from the Baraba forest-steppe, was referred to as the Northern Eurasian Formation by Chikisheva (Ibid.: 6, 56, 59, 123–124, 179–180), who borrowed this term from Viktor Bunak (1956). A similar trait combination, partly matching the broadly defined Uralian race of the traditional classifications, is observed in a number of Mesolithic and Neolithic cranial series from the forest belt of northeastern Europe—precisely those marked by the EHG component. As Chikisheva believes, the Baraba forest-steppe was populated from the forest zone of the Russian Plain in the Early Holocene (Chikisheva, Pozdnyakov, 2021). Both Eurasian formations, Northern and Southern, as she believes, are filial branches of the Boreal meta-population and have the same taxonomic rank as its two principal branches—western and eastern.
Anatoly Bagashev (1998), who disagrees with Bunak’s (1956) notion of the Uralian race in the broad sense, denies the genetic affinities between modern populations of Western Siberia (he attributes them to what he calls Western Siberian race), and Eastern European groups, which he believes to be related to other groups of Europe*. Cranial nonmetrics, on the other hand, demonstrate these affinities quite convincingly; in fact, they oppose modern Uralic-speaking groups to all other known human populations (Kozintsev, 1988: 137–140; Kozintsev, 1992; Moiseyev, Kozintsev, 1998). The integration of data relating to four independent trait systems—craniometric, cranial nonmetric, dental, and dermatoglyphic—make this result even more compelling (Moiseyev, 2001). If, therefore, rejecting the common origin of Uralic speakers on both sides of the Urals has become an anachronism, and if Vladimir Napolskikh (1997: 177–178) is right when he describes these facts as a proof of a “flesh-and-blood proto-Uralic people that lived in the past”, the question arises: How deep are the roots of the Uralic people? And how do they relate to the Northern Eurasian Formation?
The objective of this study is to revise the craniometric classification in the light of new genetic and partly linguistic facts, with special attention to distinguishing evolutionary conservatism from admixture. This motivates the choice of analytical techniques.
Material and methods
Male cranial samples representing the following cultures, periods, and territories were used:**.
-
1. Boisman culture, Primorye (Chikisheva, 2012: 38–39);
-
2. Neolithic, Yakutia (Ibid.);
-
3. Neolithic and Bronze Age, Trans-Baikal (Gokhman, 1980);
-
4. Kitoi tradition, eastern Trans-Baikal (Mamonova, 1983);
-
5. Kitoi tradition, western Trans-Baikal (Ibid.);
-
6. Kitoi tradition, Fofanovo (Gerasimova et al., 2010);
-
7. Kitoi tradition, Lena (Mamonova, 1983);
-
8. Kitoi tradition, Angara (Ibid.);
-
9. Kitoi tradition, Shamanka (D.V. Pezhemsky’s unpublished data);
-
10. Isakovo tradition, Angara (Mamonova, 1983);
-
11. Serovo tradition, Lena (Ibid.);
-
12. Serovo tradition, Angara (Ibid.);
-
13. Glazkovo tradition, western Trans-Baikal (Ibid.);
-
14. Glazkovo tradition, Fofanovo (Gerasimova et al., 2010);
-
15. Glazkovo tradition, Olkhon (Mamonova, 1983);
-
16. Glazkovo tradition, Lena (Ibid.);
-
17. Glazkovo tradition, Angara (Ibid.);
-
18. Neolithic, Krasnoyarsk-Kansk forest-steppe (Solodovnikov, Bagashev, Savenkova, 2020);
-
19. Neolithic, Lower Angara (Ibid.);
-
20. Bolshoy Mys culture, Biysk stretch of the Ob (Itkul, Kostenkova Izbushka) (Chikisheva, 2012: 36–37);
-
21. Neolithic, Barnaul stretch of the Ob (Firsovo XI) (Solodovnikov, Tur, 2017);
-
22. Kuznetsk-Altai culture, Upper Ob (Solontsy-5, Ust-Isha) (Chikisheva, 2012: 36–37);
-
23. Neolithic and Chalcolithic, Middle Irtysh (Solodovnikov et al., 2019);
-
24. Neolithic, forest-steppe Trans-Urals and northern Kazakhstan (Botai culture, Gladunino) (Khokhlov, Kitov, 2015);
-
25. Neolithic, Kuznetsk Basin (Solodovnikov, Tur, 2017);
-
26. Neolithic, Novosibirsk-Kamen stretch of the Ob (Ibid.);
-
27. Neolithic, Baraba forest steppe (Chikisheva, 2012: 36–37; Chikisheva, Pozdnyakov, Zubova, 2015);
-
28. Ust-Tartas culture, Sopka 2/3 (Chikisheva, 2012: 69–72);
-
29. Ust-Tartas culture, Sopka 2/3A (Ibid.);
-
30. Odino culture, Sopka 2/4A (Ibid.: 98–101);
-
31. Odino culture, Tartas-1 (Chikisheva, Pozdnyakov, 2019);
-
32. Odino culture, Preobrazhenka-6 (Ibid.);
-
33. Krotovo culture, Sopka 2/4B, C (Chikisheva, 2012: 98–101);
-
34. Okunev culture, Uibat (Gromov, 1997);
-
35. Okunev culture, Verkh-Askiz I (Ibid.);
-
36. Okunev culture, Chernovaya IV, VI, VIII (Ibid.);
-
37. Okunev culture, Tas-Khazaa (Ibid.);
-
38. Karakol culture, Altai (Tur, Solodovnikov, 2005);
-
39. Chaa-Khol culture, Tuva (Aimyrlyg) (Gokhman, 1980);
-
40. Yelunino culture, Upper Ob (Solodovnikov, Tur, 2003);
-
41. Samus culture, Upper Ob (Solodovnikov, 2005);
-
42. Chemurchek culture, western Mongolia (Solodovnikov, Tumen, Erdene, 2019);
-
43. Afanasyevo culture, western Mongolia (Gokhman, 1980);
-
44. Afanasyevo culture, southeastern Altai (Solodovnikov, 2009);
-
45. Afanasyevo culture, northwestern Altai (Ibid.);
-
46. Afanasyevo culture, Middle Katun (Ibid.);
-
47. Afanasyevo culture, Ust-Kuyum (Ibid.);
-
48. Afanasyevo culture, Kurota (Ibid.);
-
49. Afanasyevo culture, Ursul (Ibid.);
-
50. Afanasyevo culture, Saldyar (Ibid.);
-
51. Afanasyevo culture, Minusinsk Basin (Ibid.);
-
52. Afanasyevo culture, Karasuk III (Alekseyev, 1981);
-
53. Afanasyevo culture, Afanasyeva Gora (Ibid.);
-
54. Neolithic and Early Chalcolithic, Volga-Ural region (Khokhlov, 2017: 219–223);
-
55. Mesolithic, northern Russian Plain, Yuzhny Oleny Ostrov (Alekseyev, Gokhman, 1984);
-
56. Mesolithic, northern Russian Plain, Popovo (Gokhman, 1984);
-
57. Mesolithic, eastern Baltic, Zvejnieki (Denisova, 1975: 187–188);
-
58. Early Neolithic, eastern Baltic, Zvejnieki (Ibid.: 193–194);
-
59. Middle and Late Neolithic, eastern Baltic, Zvejnieki (Ibid.: 202–203);
-
60. Pit-Comb Ware culture, northern Russia and the Volga-Oka watershed (Chikisheva, 2012: 38–39);
-
61. Volosovo culture, the Volga-Oka watershed (Ibid.);
-
62. Khvalynsk culture, Khvalynsk cemeteries (Khokhlov, 2017: 226–230);
-
63. Khvalynsk culture, Khlopkov Bugor (Ibid.: 230–231);
-
64. Mesolithic, Ukraine (Konduktorova, 1973: 13–14);
-
65. Neolithic, Ukraine, Dnieper-Donets culture (Potekhina, 1999: 190–192);
-
66. Early Chalcolithic, Ukraine, Sredny Stog II culture (Ibid.: 204–208).
The trait battery includes 14 key traits (listed with their standard codes, see (Alekseyev, Debetz, 1964)): cranial length (1), cranial breadth (8), cranial height (17), minimal frontal breadth (9), bizygomatic breadth (45), upper facial height (48), nasal height (55), nasal breadth (54), orbital breadth (51), orbital height (52), naso-malar angle (77), zygo-maxillary angle, simotic index (SS : SC), and nasal prominence angle (75 (1)). Measurements were processed using the multiple discriminant (canonical) analysis, and Mahalanobis D2 distances corrected for sample size were calculated. The distance matrix was subjected to nonmetric multidimensional scaling, and the minimum spanning tree, showing the shortest path between the points in the multivariate space and therefore optimal for detecting clines, was constructed. Boris Kozintsev’s program CANON for calculating Mahalanobis distances corrected for sample size and Øyvind Hammer’s software package PAST (version 4.05) were used (Hammer, 2012)*.
The minimum spanning tree method generates clusters less formally than the traditional cluster analysis does. But such clusters make more sense in terms of geography*. Cluster analysis is unsuited for studying clines, and therefore is used here only as an auxiliary device, in two varieties—unweighted pair-group (UPGMA) and neighbor joining (NJ); both techniques were implemented in respective modules of the PAST package. The resulting dendrograms are not shown, because of their large size, but can be obtained by e-mail upon request.
Results
The two-dimensional projection of the group constellation is shown in Fig. 1. We will examine it in the direction of the principal gradient of craniometric variation—from the eastern (Mongoloid) extreme to the western (Caucasoid). This gradient is generally consistent with the geographic position of groups.
Groups of the eastern extreme and Paleosiberian. Samples with the utmost expression of Mongoloid features include the following (ranked in the order from greater to lesser expression, i.e., in the increasing order of NMDSCAL coordinate 1): Kitoi of eastern Trans-Baikal (No. 4), Boisman of Primorye (No. 1), Kitoi of western Trans-Baikal (No. 5), and Neolithic of Yakutia (No. 2).
Does craniometry show the Kitoi people to be more Mongoloid than representatives of later traditions of the Baikal Neolithic and Bronze Age, as the results of the genetic studies suggest? Perhaps the use of individual data would confirm that, but the comparison of averages yields an indeterminate result. The mean value of coordinate 1 (Fig. 1) in six Kitoi samples (No. 4–9) is –0.147 ± 0.032, and the respective value for eight samples from later burials on both coasts of Lake Baikal (No. 10–17) equals –0.098 ± 0.007. The difference, according to the Mann-Whitney test, may be due to chance ( U = 16, p = 0.33). If only distances between the Baikal groups are subjected to NMDSCAL (Fig. 2), the difference between Kitoi and the remaining groups remains insignificant, despite the extreme expression of Mongoloid features in the Kitoi sample from Trans-Baikal**.
The geographic factor turns out to be more important than that relating to chronology. In Fig. 1, the TransBaikal cluster is opposed to that of Cis-Baikal by being definitely more Mongoloid (apart from the nine samples from the Cis-Baikal area, the latter cluster includes two Neolithic crania from the Lower Angara (No. 19) and two Chemurchek crania from western Mongolia
(No. 42))*. The mean value of coordinate 1 in five TransBaikal samples (disregarding the pooled Trans-Baikal group) equals –0.172 ± 0.029, whereas the respective value for nine Cis-Baikal samples is -0.088 ± 0.006. According to the Mann-Whitney test, the difference is highly significant ( U = 0.5, p = 0.004). Trans-Baikal groups differ from those of the Cis-Baikal also in Fig. 2, where other groups are excluded from the analysis; but in this case it is only a tendency, which does not reach the required significance level on any of the two NMDSCAl coordinates.
Eurasian formations—Northern and Southern. After Chikisheva’s publications it appeared that the eastern boundary of the distribution area of the Northern Eurasian Formation is marked by the Ob River. Apart from the seven Neolithic and Early Bronze Age samples from the Baraba forest-steppe, which she mentioned (Fig. 1, No. 27–33), this cluster includes the Neolithic sample from the Novosibirsk-Kamen stretch of the Ob (No. 26). Among the two cluster analytic techniques, admittedly, this is confirmed only by NJ, but not by UPGMA. However, one more Neolithic sample from a much more easterly area, the Kuznetsk Basin (No. 25), merges with the Baraba cluster by all the techniques used—minimum spanning tree and both versions of cluster analysis, implying that the Northern Eurasian formation should be extended eastwards, possibly even as far as the Yenisei. Notably, it is the Northern, not the Southern Eurasian, formation that is directly linked with the Eastern Siberian clusters: the minimum spanning tree edge connects the sample from Preobrazhenka-6 (No. 32) with that from the Lower Angara (No. 19).
On the contrary, the Southern Eurasian formation, as it appeared until recently, spread from the Yenisei in the western direction, because the Neolithic people of the Krasnoyarsk-Kansk forest-steppe (No. 18) seemed to be the best candidates for the role of the ancestors of the Okunev people (No. 34–37). It now turns out, however, that the Neolithic and Chalcolithic people who lived well to the west—on the Middle Irtysh (No. 23)—are likewise suited for that role. The authors of the publication attempt to split this group in terms of geography, claiming that crania from the forest and forest-steppe zone display features of the Northern Eurasian formation, whereas those from the steppe are closer to the Southern Eurasian formation (Solodovnikov et al., 2019). But the sample is very small, and according to its average values falls within the Southern formation, which is supported by the cluster analysis. Four samples from the Upper Ob—Bolshoy
*The advantage of the minimum spanning tree method over cluster analysis is evident in this case: none of the two clustering algorithms, neither UPGMA nor NJ, is capable of separating Cis-Baikal groups from those of Trans-Baikal. Instead, members of each of these clusters are separated from one another and intermixed with samples from other regions.
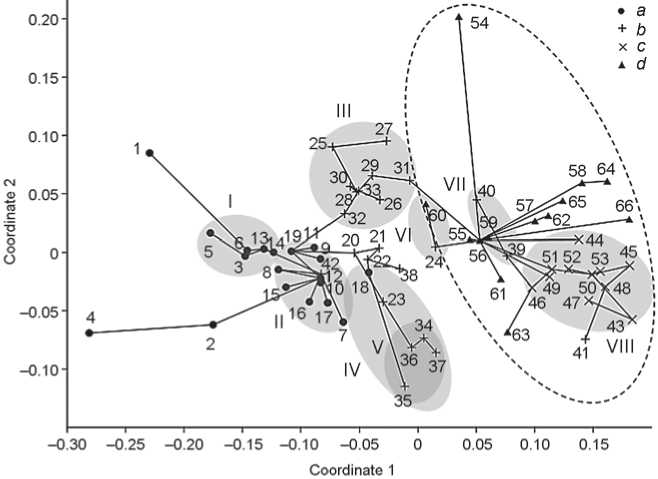
Fig. 1 . The position of male cranial samples in the space generated by two axes of the nonmetric multidimensional scaling of Mahalanobis D2 distances, corrected for sample size.
Straight lines are edges of the minimum spanning tree showing the shortest path between points in the original multivariate space. Spots show clusters: I – Paleosiberian, Trans-Baikal, II – Paleosiberian, Cis-Baikal, III – Northern Eurasian formation, IV – Southern Eurasian formation, V – Okunev, VI – Pit-Comb Ware, VII – Chaa-Khol-Yelunino, VIII – Afanasyevo. The dashed contour encloses groups displaying the western trait combination. a – groups of Eastern Siberia and the Russian Far East; b – groups of Southern and Western Siberia, and Western Mongolia except Afanasyevan; c – Afanasyevan; d – European. Numbers of groups refer to the list (see text).
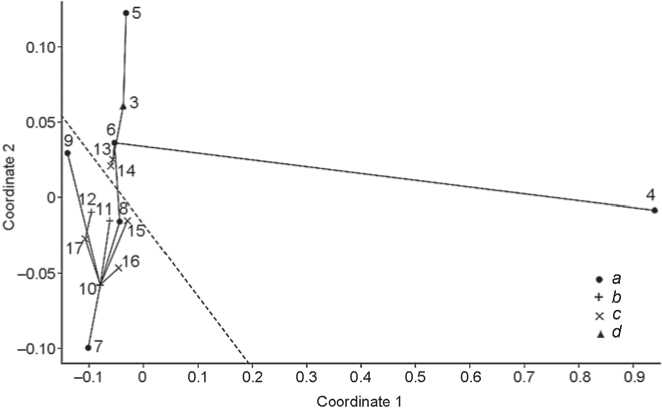
Fig. 2 . The position of male cranial samples from Cis-Baikal and Trans-Baikal in the space generated by two axes of the nonmetric multidimensional scaling of the Mahalanobis D2 distances corrected for sample size.
a – Kitoi, b – Isakovo and Serovo, c – Glazkovo, d – tradition not indicated. The dashed line separates Cis-Baikalian groups from those of Trans-Baikal. See Fig. 1 for other conventions. Numbers of groups refer to the list (see text).
Mys from Itkul and Kostenkova Izbushka (No. 20), Neolithic from Firsovo XI (No. 21), that of the Kuznetsk-Altai culture from Ust-Isha and Solontsy-5 (No. 22), and Karakol (No. 38)—take an intermediate position between the two formations, without falling in any of them, according to the results of cluster analysis. Not far from these groups, in the interval between them and the western hyper-cluster, is a sample from the forest-steppe Trans-
Urals and northern Kazakhstan (No. 24), which includes the few Botai crania. Displaying the “westernmost” trait combination among all the Neolithic groups of northern Asia, this sample forms a separate pair with the Pit-Comb Ware group (No. 60)—the most Mongoloid-like among the European groups.
As T.A. Chikisheva believes, Siberian members of the Northern Eurasian formation, unlike those belonging to the Southern Eurasian formation, are somewhat similar to Mesolithic and Neolithic groups of the forest belt of northeastern Europe. Indeed, the Odino sample from Tartas-1 (No. 31) is directly connected with the Mesolithic sample from Popovo (No. 56) by the minimum spanning tree’s edge. However, the average D2 distance separating eight Mesolithic and Neolithic samples from the forest zone of northeastern Europe (No. 54–61) from nine members of the Northern Eurasian formation of Siberia (No. 25–33) is only slightly less than that separating the former from the six members of the Southern Eurasian formation (No. 18, 23, 34–37): 13.90 versus 15.60, respectively. According to the Wilcoxon test for paired data, the difference is insignificant ( z = 1.82, p = 0.069). Therefore, the available data do not warrant the idea of a single Northern Eurasian formation spanning the area from the eastern Baltic to the Kuznetsk Basin. So far, it can only be stated that both Eurasian formations of Siberia taken together and groups that are close to them fill in the gap between Paleosiberian, specifically Cis-Baikal, populations and European groups in which the expression of western traits is relatively weak.
Western groups. Apart from groups of the forest zone (see above), the weaker expression of western traits is seen in the Khvalynsk sample from Khlopkov Bugor (No. 63), which, on the western scale, is markedly different from the Khvalynsk sample from the eponymous cemeteries (No. 62), and in two Siberian groups of an arguably European origin—Chaa-Khol (No. 39) and Yelunino (No. 40), which make up a separate pair. A considerable temporal variation is observed in the Zvejnieki population: a moderately western pattern in the Mesolithic (No. 57) is replaced by a pronouncedly western combination in the Early Neolithic (No. 58), after which the expression degree of western features drops again sharply, and the series from the Middle and Late Neolithic burials (No. 59) is markedly shifted toward the Pit-Comb Ware sample. The opposite, western, extreme of the scale is taken by three samples from Ukraine (No. 64–66), and by most Afanasyevan series, to which the Samus sample (No. 41) is close.
Discussion
Material and methods used in this study have failed to demonstrate the difference between the Kitoi people as a whole and those representing later traditions of the Baikal Neolithic and Bronze Age. There is no doubt, however, that the eastern component spread westwards from across Lake Baikal. This is evidenced by two extremely Mongoloid crania from the eastern Trans-Baikal (in this respect they surpass even the Boisman sample), by a direct link between the latter and the Trans-Baikal cluster, and by a statistically significant difference between the Trans-Baikal and Cis-Baikal clusters.
Because the European admixture, as we now know, reached Lake Baikal only in exceptional cases at that time, the position of groups on the east-to-west axis indicates admixture between the eastern component and that traditionally known as Paleosiberian, or, in genetic terms, between groups marked by the NEA/AEA component and those marked by the ANE component, respectively. The same applies to both Eurasian formations. At the stage when the process of disintegration of the Boreal meta-population had not yet terminated, genetic and craniometric gradients directed east to west, from Eastern Siberia to Eastern Europe (and perhaps further west), testify, apparently, not so much to admixture as to the isolation by distance effect. This effect was evidently the principal reason why numerous groups inhabiting vast territories of Eurasia demonstrate all transitional stages between the two extremes revealed by craniometric traits—eastern and western.
Those closest to the western extreme in our sample are three groups from the Ukraine, including the Sredny Stog people (No. 66), and the Afanasyevans. Indications that the former were likely ancestors of the latter have been provided both by archaeologists (Nikolaeva, 2019a, b) and by physical anthropologists (Solodovnikov, 2009). The plot (Fig. 1) shows this rather clearly. The temporal dynamics of the physical type of the Zvejnieki people, specifically the “eastern” tendency displayed by the sample from the Middle and Late Neolithic burials, recalls archaeological evidence of contacts with the Pit-Comb Ware people and genetic facts showing the rise in the EHG autosomal component at that stage (Jones et al., 2017).
Genetic data suggest that the role of the Early Bronze Age migrants from the western steppe as a source of European admixture in Siberia should not be overstated. In the words of Narasimhan et al. (2019: Suppl. materials, p. 235), on their way to the east, Afanasyevans “leapfrogged” the autochthonous populations, without intermixing with them. Chikisheva (2012: 180), therefore, was absolutely right in claiming that vis-à-vis the arguably local origin of populations belonging to the Southern Eurasian formation, “the impact of migrations on the origins of the Altai-Sayan groups was somewhat exaggerated” (for a new summary of data relating to the Okunev population and supporting this idea, see (Kozintsev, 2020)).
But the same conclusion, to all appearances, is true with regard to the Northern Eurasian formation. I see no particular reason to ascribe its origin to a migration from the forest zone of northeastern Europe to Western Siberia. As concerns the relationship between the Northern Eurasian formation and the Uralian race sensu lato of traditional classifications, the issue cannot be resolved with the database used in this study. First, it proved impossible to demonstrate that either of the two Eurasian formations of Siberia is closer to the Mesolithic and Neolithic people of the northeastern European forest zone than the other. Second, given the unusually high degree of biological specificity of modern Uralic-speaking groups on both sides of the Urals (see above), looking for the sources of proto-Uralians in such a vast territory in the Mesolithic or Neolithic is pointless. No doubt the Uralic homeland was situated in the taiga zone of Western Siberia and, possibly, in the adjoining part of northwestern Urals (Napolskikh, 1997: 132, 140; Janhunen, 2009). Early cranial finds from those territories are quite scarce. The more western parts of the forest zone, specifically the Volga basin and the Baltic, were populated by the Uralic (specifically Finno-Ugric) speakers no earlier that the second millennium BC (Napolskikh, 1997: 125, 197–198; Janhunen, 2009).
The same applies to the more southerly regions, such as the forest-steppe zone of Western Siberia, where Uralic speakers appeared likewise late, as evidenced by cranial nonmetric data, especially sensitive to the Uralic component (Gromov, Moiseyev, 2004; Moiseyev, 2006). The combination of craniometric and cranial nonmetric traits displayed either by the Okunev people, or by those buried at Sopka, regardless of their chronological position, or by people with a western genetic legacy (Afanasyevo, Andronovo, Karasuk, Irmen, and Tagar) does not point in the Uralian direction (Kozintsev, 2004). The “Uralic” trait combination first appears in a group from Yelovka II in the Tomsk stretch of the Ob, dating to the Andronovo era but hardly representing migrants from the west (Ibid.), and this is the earliest evidence suggestive of a southward migration of Uralic speakers from the taiga to the subtaiga zone of Western Siberia in the Final Bronze Age.
The origin of the Chemurchek people remains mysterious. Genetic studies suggest that they had originated from a complex mixture of western, eastern, and southern constituents. One of the main components, ANE, could have been received from the Botai people (Jeong et al., 2020; Wang et al., 2021). Craniometric analysis (Fig. 1) places the two Chemurchek crania (No. 42) in the CisBaikal cluster, but because of the admixture revealed by genetics, this fact is hardly indicative of origin.
The sample that includes the few Botai crania (No. 24) takes a more “western” position (Fig. 1), forming a pair with the Pit-Comb Ware group of European Russia, and this is supported by archaeological data suggesting that these cultures are related (Mosin, 2003: 97–98). However, the idea that they were associated with Finno-Ugrians
(Ibid.) disagrees with numerous facts demonstrating a later penetration of Finno-Ugric tribes into areas west of the Cis-Urals (see above).
Conclusions
-
1. Craniometric data support the conclusions made by geneticists about the early (no later than the Neolithic) penetration of the eastern component from the eastern Trans-Baikal to the Cis-Baikal, where it mixed with the autochthonous (Paleosiberian) component.
-
2. The Northern and the Southern Eurasian formations, as well as groups that are close to them, take an intermediate position between the Cis-Baikal (Paleosiberian) cluster and European groups with a weak expression of western traits. The relationship between the Northern Eurasian formation and the Uralian race of traditional classifications is unclear.
-
3. East-west gradients revealed by both genetic and craniometric traits in northern Eurasia apparently do not indicate admixture, the extent of which during the Neolithic and Early Bronze Age was minor, at least in central and western parts of northern Eurasia. Rather, they suggest that the divergence of groups filial with regard to the Boreal meta-population was still incomplete at that time.
-
4. Groups that can be considered admixed (Chaa-Khol and Yelunino) constitute a small minority and are opposed both to autochthonous groups, specifically members of the Paleosiberian cluster and both Eurasian formations, on the one hand, and migrant ones such as the Afanasyevan, on the other.
Acknowledgments
I am thankful to Denis Pezhemsky for permission to use his unpublished data about the Shamanka sample, and to Tatiana Chikisheva for her valuable considerations.
